
15.3: 真核生物转录
- Page ID
- 203066

培养技能
- 列出真核生物转录的步骤
- 讨论 RNA 聚合酶在转录中的作用
- 比较和对比三种 RNA 聚合酶
- 解释转录因子的重要性
原核生物和真核生物的转录过程基本相同,但有一些关键区别。 原核生物和真核生物之间最重要的区别是后者的膜结合核和细胞器。 在基因结合在细胞核中后,真核细胞必须能够将其mRNA运送到细胞质中,并且在翻译之前必须保护其mRNA免受降解。 真核生物还使用三种不同的聚合酶,每种聚合酶转录不同的基因子集。 真核生物 mRNA 通常是单基因的,这意味着它们指定了单一蛋白质。
在真核生物中启动转录
与可以单独与DNA模板结合的原核聚合酶不同,真核生物需要其他几种称为转录因子的蛋白质才能首先与启动子区域结合,然后帮助招募适当的聚合酶。
三种真核生物 RNA 聚合酶
真核生物 mRNA 合成的特征明显比原核生物更复杂。 真核生物不是包含五个亚基的单一聚合酶,而是三种聚合酶,每个聚合酶由 10 个或更多亚基组成。 每种真核聚合酶还需要一组不同的转录因子才能将其带入DNA模板。
RNA 聚合酶 I 位于核仁中,核仁是一种特殊的核子结构,其中核糖体 RNA (rRNA) 被转录、加工并组装成核糖体(表\(\PageIndex{1}\))。 rRNA 分子被认为是结构 RNA,因为它们具有细胞作用,但不会转化为蛋白质。 rRNA 是核糖体的组成部分,对翻译过程至关重要。 RNA 聚合酶 I 合成了除 5S rRNA 分子之外的所有 rRNA。 “S” 名称适用于 “Svedberg” 单位,这是一个非累加值,用于表征离心过程中粒子沉积的速度。
表\(\PageIndex{1}\):三种真核生物 RNA 聚合酶的位置、产物和灵敏度
| RNA 聚合酶 | 蜂窝隔间 | 转录产物 | α-Amanitin 敏感性 |
|---|---|---|---|
| 我 | Nucleolus | 除 5S rRNA 之外的所有 rRNA | 麻木不仁 |
| 二 | 核 | 所有蛋白质编码的核 pre-mRNA | 极其敏感 |
| 三 | 核 | 5S rRNA、tRNA 和小型核 RNA | 中度敏感 |
RNA 聚合酶 II 位于细胞核中,可合成所有编码蛋白质的核 pre-mRNA。 真核生物 pre-mRNA 在转录后但在翻译之前经过大量处理。 为清楚起见,本模块对真核生物转录和翻译的讨论将使用 “mRNA” 一词来仅描述准备翻译的成熟、经过处理的分子。 RNA 聚合酶 II 负责转录绝大多数真核基因。
RNA 聚合酶 III 也位于细胞核中。 这种聚合酶转录各种结构RNA,包括5S pre-rrna、转移前 RNA(pre-trNA)和小型核前 RN A。 tRNA在翻译中起着至关重要的作用;它们是mRNA模板和不断增长的多肽链之间的适配分子。 小核 RNA 具有多种功能,包括 “剪接” pre-mRNA 和调节转录因子。
对新基因进行表征的科学家可以通过测试该基因是否在特定的蘑菇毒药α-amanitin存在的情况下表达来确定哪种聚合酶对其进行了转录(上表)。 有趣的是,由死亡帽蘑菇 Am anita phalloides 生产的 α-amanit in 对这三种聚合酶的影响截然不同。 RNA 聚合酶 I 对 α-amanitin 完全不敏感,这意味着聚合酶可以在存在这种毒药的情况下体外转录 DNA。 相比之下,RNA聚合酶II对α-氨基丁极为敏感,而RNA聚合酶III对α-氨基丁非常敏感。 了解转录聚合酶可以让研究人员了解正在研究的基因的一般功能。 由于 RNA 聚合酶 II 转录了绝大多数基因,因此我们将在后续关于真核生物转录因子和启动子的讨论中重点讨论这种聚合酶。
RNA 聚合酶 II 启动子的结构
真核生物启动子比原核生物启动子大得多,也更复杂,但两者都有 TATA 盒子。 例如,在小鼠胸苷激酶基因中,TATA盒相对于起始(+1)位点位于大约-30(图\(\PageIndex{1}\))。 对于这个基因,确切的 TATA 盒序列是 TATAAAA,如非模板链上在 5' 到 3' 方向上读取的那样。 该序列与大肠杆菌 TATA 盒不同,但它保留了富含 A—T 的元素。 A—T 键的热稳定性很低,这有助于 DNA 模板局部解开,为转录做准备。
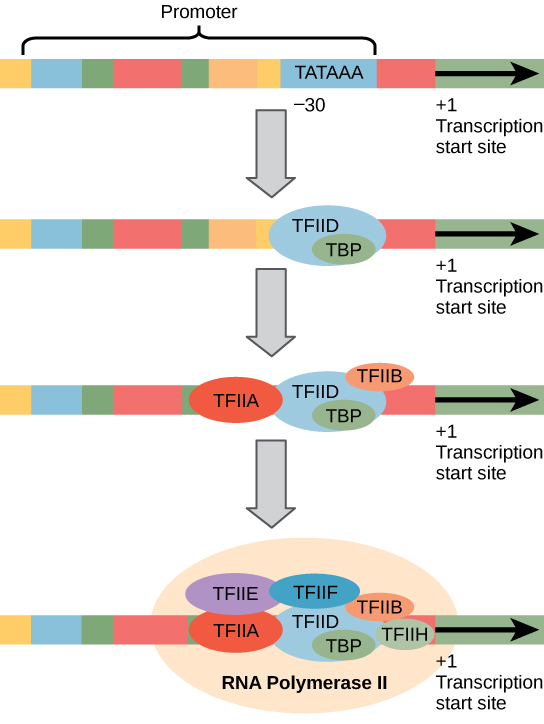
艺术连接

一位科学家在细菌基因前面拼接真核生物启动子,然后将该基因插入细菌染色体中。 你会指望细菌转录基因吗?
小鼠基因组包括细胞质胸苷激酶的一个基因和两个假基因。 假基因是已经失去蛋白质编码能力或不再被细胞表达的基因。 这些假基因是从 mRNA 中复制并整合到染色体中。 例如,小鼠胸苷激酶启动子还有一个保守的 CAAT 盒(GGCCAATCT),大约为 -80。 该序列是必需的,与结合转录因子有关。 在 TATA 盒子的上游,真核生物启动剂还可能包含一个或多个富含 GC 的盒子(GGCG)或八聚体盒(ATTTGCAT)。 这些元素结合了提高转录启动效率的细胞因子,通常在细胞不断表达的更 “活性” 的基因中鉴定。
RNA 聚合酶 II 的转录因子
真核生物转录的复杂性并不以聚合酶和启动子结束。 一大批基础转录因子、增强剂和消音剂也有助于调节从基因合成 pre-mRNA 的频率。 增强剂和消音剂会影响转录效率,但不是转录进行所必需的。 基础转录因子对于在 DNA 模板上形成启动前复合物至关重要,该复合物随后会招募 RNA 聚合酶 II 进行转录启动。
基础转录因子的名称以 “TFII”(这是 RNA 聚合酶 II 的转录因子)开头,并用字母 A—J 表示。转录因子系统地落在 DNA 模板上,每种转录因子进一步稳定启动前复合物并做出贡献用于招募 RNA 聚合酶 II。
将 RNA 聚合酶 I 和 III 引入 DNA 模板的过程涉及稍微不那么复杂的转录因子集合,但总体主题是一样的。 真核生物转录是一个严格监管的过程,需要各种蛋白质相互作用以及与 DNA 链相互作用。 尽管真核生物的转录过程比原核生物的转录需要更多的代谢投资,但它可以确保细胞精确地转录蛋白质合成所需的前 mRNA。
进化连接:发起人的演变
基因的进化可能是一个熟悉的概念。 在DNA复制过程中,基因中可能发生突变,其结果可能对细胞有益,也可能没有益处。 通过改变酶、结构蛋白或其他因素,突变过程可以改变功能或物理特征。 但是,真核启动子和其他基因调控序列也可能进化。 例如,假设一个经过几代人对细胞更有价值的基因。 也许该基因编码一种结构蛋白,细胞需要大量合成才能发挥某种功能。 如果是这样的话,那么该基因的启动子更有效地招募转录因子并增加基因表达将对细胞有益。
研究启动子序列演变的科学家报告了不同的结果。 部分原因是很难准确推断出真核生物启动子的起点和终点。 一些启动子存在于基因内;其他启动子位于它们所调控的基因的上游甚至下游。 但是,当研究人员将检查局限于人类核心启动子序列(这些序列通过实验定义为结合启动前复合物的序列)时,他们发现启动子的进化速度甚至比蛋白质编码基因还要快。
目前尚不清楚启动子的进化与人类或其他更高生物的进化有何对应。 但是,启动子进化以有效地或多或少地产生给定基因产物,这是基因本身进化的有趣替代方案。 1
RNA 聚合酶 I 和 III 的启动子结构
在真核生物中,由RNA聚合酶I、II和III转录的基因的保守启动子元素不同。 RNA 聚合酶 I 转录在 -45 至 +20 区域有两个富含 GC 的启动子序列的基因。 光是这些序列就足以发生转录启动,但是在起始位点上游的-180至-105区域具有额外序列的启动子将进一步增强转录的启动。 由RNA聚合酶III转录的基因具有上游启动子或存在于基因本身中的启动子。
真核生物伸长率和终止
启动前复合物形成后,聚合酶从其他转录因子中释放出来,允许伸长率像在原核生物中一样进行,聚合酶沿5'至3'方向合成pre-mRNA。 如前所述,RNA聚合酶II转录了大部分真核基因,因此本节将重点介绍这种聚合酶如何实现伸长和终止。
尽管真核生物和原核生物的酶促伸长过程基本相同,但DNA模板更为复杂。 当真核细胞没有分裂时,它们的基因以DNA和称为染色质的蛋白质的弥漫块形式存在。 DNA 以重复的间隔紧密包裹在带电的组蛋白周围。 这些DNA—组蛋白复合物,统称为核小体,间隔规律,包括146个DNA核苷酸,缠绕在八个组蛋白周围,就像线轴周围的线一样。
为了实现多核苷酸的合成,转录机制需要在每次遇到核小体时将组蛋白移开。 这是由一种名为F AC T的特殊蛋白质复合物完成的,它代表 “促进染色质转录”。 当聚合酶沿着DNA模板移动时,这种复合物会将组蛋白从DNA模板中拉出。 一旦合成了 pre-mRNA,FACT 复合物就会取代组蛋白来重现核小体。
对于不同的聚合酶,转录的终止是不同的。 与原核生物不同,RNA聚合酶II在真核生物中的伸长发生在转录基因末端以外的1,000—2,000个核苷酸。 在 mRNA 处理过程中,这条 mRNA 之前的尾巴随后会通过裂解去除。 另一方面,RNA聚合酶I和III需要终止信号。 由 RNA 聚合酶 I 转录的基因包含可被终止蛋白识别的特定 18 核苷酸序列。 RNA 聚合酶 III 的终止过程涉及一个 mRNA 发夹,类似于原核生物中不依赖于 rho 的转录终止。
摘要
真核生物中的转录涉及三种聚合酶中的一种,具体取决于被转录的基因。 RNA 聚合酶 II 转录所有蛋白质编码基因,而 RNA 聚合酶 I 转录 rRNA 基因,而 RNA 聚合酶 III 转录 rRNA、tRNA 和小核 RNA 基因。 真核生物转录的启动涉及几种转录因子与通常位于被复制基因上游的复杂启动子序列的结合。 mRNA 在 5' 到 3' 方向上合成,FACT 复合物在聚合酶经过时移动并重新组装核小体。 RNA聚合酶I和III通过依赖蛋白质或RNA发夹的方法终止转录,而RNA聚合酶II转录了基因模板以外的1,000个或更多核苷酸,并在mRNA前处理过程中分解多余的核苷酸。
艺术联系
图\(\PageIndex{2}\):一位科学家在细菌基因前拼接真核生物启动子,然后将该基因插入细菌染色体中。 你会指望细菌转录基因吗?
- 回答
-
不。 原核生物使用的启动子与真核生物不同。
脚注
- 1 H Liang 等人,“灵长类动物基因组中核心启动子的快速进化”,《分子生物学与进化》25(2008):1239—44。
词汇表
- CAAT 盒子
- (GGCCAATCT) 参与结合转录因子的必需真核生物启动子序列
- 事实
- 通过在转录 RNA 聚合酶 II 之前分解核小体,然后在聚合酶通过后重新组装核小体,“促进染色质转录” 的复合物
- 富含 GC 的盒子
- (GGCG) 结合细胞因子以提高转录效率的非必需真核生物启动子序列;可能在启动子中多次出现
- 八聚体盒
- (ATTTGCAT) 结合细胞因子以提高转录效率的非必需真核生物启动子序列;可能在启动子中多次出现
- 启动前复合物
- 招募 RNA 聚合酶 II 以转录 DNA 模板的转录因子和其他蛋白质集群
- 小核 RNA
- 由 RNA 聚合酶 III 合成的分子,具有多种功能,包括剪接前 mRNA 和调节转录因子