
15.2: 原核转录
- Page ID
- 203079

培养技能
- 列出原核转录的不同步骤
- 讨论启动子在原核转录中的作用
- 描述转录如何以及何时终止
原核生物,包括细菌和古细菌,大多是单细胞生物,顾名思义,它们缺乏膜结合的核和其他细胞器。 细菌染色体是一个共价封闭的圆圈,与真核染色体不同,它不是围绕组蛋白组织的。 原核生物 DNA 所在的细胞中心区域称为核样体。 此外,原核生物通常具有丰富的质粒,质粒是较短的环状DNA分子,可能只包含一个或几个基因。 在细胞分裂过程中,质粒可以独立于细菌染色体转移,并且通常具有抗生素耐药性等特征。
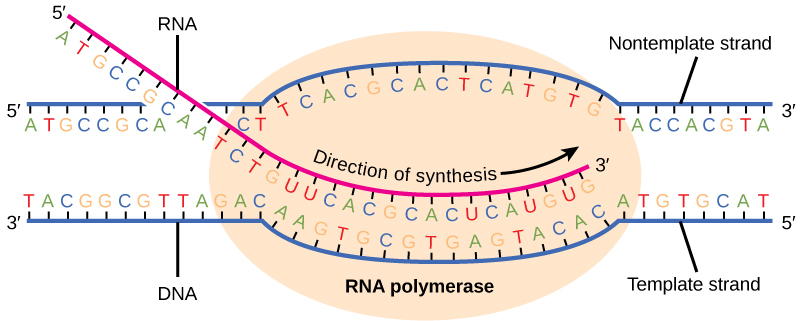
原核生物(和真核生物)中的转录需要 DNA 双螺旋在 mRNA 合成区域部分解开。 放卷区域称为转录气泡。 对于每个基因,转录总是从相同的 DNA 链进行,这称为模板链。 mRNA 产物与模板链互补,与另一条 DNA 链(称为非模板链)几乎相同。 唯一的区别是,在 mRNA 中,所有 T 核苷酸都被 U 核苷酸所取代。 在 RNA 双螺旋中,A 可以通过两个氢键结合 U,就像 DNA 双螺旋中的 A—T 配对一样。
DNA双螺旋中与第一个5'mRNA核苷酸转录的位点相对应的核苷酸对称为+1位点或起始位点。 起始位点之前的核苷酸被指定为负数,并被指定为上游。 相反,起始位点之后的核苷酸用 “+” 编号表示,被称为下游核苷酸。
在原核生物中启动转录
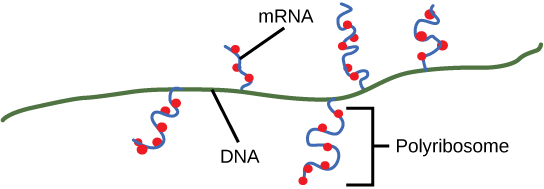
原核生物没有膜封闭的核。 因此,转录、翻译和 mRNA 降解过程可以同时发生。 在同一 DNA 模板上同时发生的多个转录和翻译事件可以快速扩增细菌蛋白的细胞内水平。 原核转录通常涵盖多个基因,并产生指定多种蛋白质的多环共体 mRNA。
我们在这里的讨论将通过描述大肠杆菌(一种经过充分研究的细菌物种)中的转录过程来举例说明转录。 尽管大肠杆菌的转录与古细菌中的转录之间存在一些差异,但对大肠杆菌转录的理解几乎可以应用于所有细菌物种。
原核 RNA 聚合酶
原核生物使用相同的 RNA 聚合酶来转录其所有基因。 在大肠杆菌中,聚合酶由五个多肽亚基组成,其中两个是相同的。 其中四个亚单位,分别表示为 α、α、β 和 β',构成了聚合酶核心酶。 这些亚基每次转录基因时都会组装,转录完成后它们就会分解。 每个亚单位都有独特的作用;两个 α-亚基是在 DNA 上组装聚合酶所必需的;β-亚基与将成为新生的 “新生的” mRNA 分子一部分的三磷酸核苷结合;β' 结合 DNA 模板链。 第五个亚单位 α 仅参与转录启动。 它具有转录特异性,使得聚合酶开始从适当的起始位点合成 mRNA。 如果没有 α,核心酶将从随机位点转录,并产生指定蛋白质胡言乱语的 mRNA 分子。 由所有五个亚单位组成的聚合酶称为全酶。
原核生物启动子
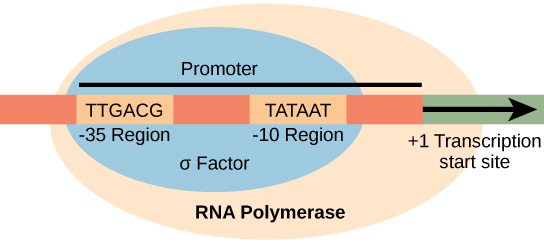
启动子是一种DNA序列,转录机制在该序列上结合并启动转录。 在大多数情况下,启动子存在于它们调节的基因的上游。 启动子的特定序列非常重要,因为它决定了相应的基因是一直被转录、有时转录还是不经常被转录。 尽管启动子因原核生物基因组而异,但仍有一些元素是保守的。 在起始位点上游的-10和-35区域,有两个启动子共识序列,或者所有启动子和各种细菌物种之间相似的区域(图\(\PageIndex{1}\))。 -10 共识序列,称为 -10 区域,是 TATAAT。 -35 序列 TTGACA 被识别并受到 α 的绑定。 一旦发生这种相互作用,核心酶的亚单位就会与该位点结合。 富含 A—T 的 -10 区域便于 DNA 模板的解开,并形成了多个磷酸二酯键。 转录启动阶段以产生流产转录本而告终,这些转录本是制造和释放的大约10个核苷酸的聚合物。
链接到学习
观看这段 MolecularMovies 动画,看看 TATA 盒子转录的第一部分和基本序列重复。
原核生物的伸长率和终止率
转录伸长阶段从聚合酶中释放 α 亚单位开始。 α 的解离允许核心酶沿着 DNA 模板进行,以每秒大约 40 个核苷酸的速度在 5' 到 3' 方向上合成 mRNA。 随着伸长率的推进,DNA在核心酶前面不断展开,然后在核心酶后面倒带(图\(\PageIndex{2}\))。 DNA 和 RNA 之间的碱基配对不够稳定,无法维持 mRNA 合成成分的稳定性。 相反,RNA聚合酶充当DNA模板和新生RNA链之间的稳定连接器,以确保伸长不会过早中断。
原核生物终止信号
基因被转录后,需要指示原核聚合酶与DNA模板分离并释放出新产生的mRNA。 根据被转录的基因,有两种终止信号。 一种是基于蛋白质的,另一种是基于RNA的。 Rho 依赖性终止由 rho 蛋白控制,该蛋白在不断增长的 mRNA 链上追踪聚合酶后面。 在基因快要结束时,聚合酶在DNA模板上遇到一连串的G核苷酸并停滞不前。 结果,rho 蛋白与聚合酶发生碰撞。 与 rho 的相互作用会从转录气泡中释放 mRNA。
不@@ 依赖于 Rho 的终止由 DNA 模板链中的特定序列控制。 当聚合酶接近被转录基因的末端时,它会遇到一个富含C—G核苷酸的区域。 mRNA 会自行折叠,互补的 C—G 核苷酸结合在一起。 结果是一个稳定的发夹,一旦聚合酶开始转录富含 A—T 核苷酸的区域,它就会停滞。 mRNA 转录本的互补 U-A 区域仅与模板 DNA 形成微弱的相互作用。 再加上停滞的聚合酶,诱导了足够的不稳定性,使核心酶能够分离并释放新的mRNA转录本。
终止后,转录过程即告完成。 当终止时,原核转录本已经被用来开始合成大量编码蛋白质拷贝了,因为这些过程可以同时发生。 转录、翻译甚至 mRNA 降解的统一是可能的,因为所有这些过程都发生在相同的 5' 到 3' 方向上,而且原核细胞中没有膜分隔(图\(\PageIndex{3}\))。 相比之下,真核细胞中存在细胞核就无法同时进行转录和翻译。
链接到学习
观看这部 BioStudio 动画,看看原核生物转录的过程。
摘要
在原核生物中,mRNA合成始于DNA模板上的启动子序列,该启动子序列包含两个招募RNA聚合酶的共识序列。 原核聚合酶由一个由四个蛋白质亚单位组成的核心酶和一个仅在启动时起作用的 α 蛋白组成。 伸长率以每秒 40 个核苷酸的速度在 5' 到 3' 方向上合成 mRNA。 终止可以释放 mRNA,要么是通过 rho 蛋白相互作用发生的,要么是通过 mRNA 发夹的形成。
词汇表
- 共识
- 许多物种使用的 DNA 序列来执行相同或相似的功能
- 核心酶
- 由 α、α、β 和 β' 组成的原核 RNA 聚合酶,但缺少 α;这种复合物具有伸长作用
- 下游
- 在 mRNA 转录方向上跟随起始位点的核苷酸;一般而言,相对于 mRNA 上的位点朝向 3' 末端的序列
- 簪
- RNA 自身折叠并在互补核苷酸之间形成分子内氢键时的结构
- 全酶
- 由 α、α、β、β' 和 α 组成的原核 RNA 聚合酶;这种复合物负责转录的启动
- 起始部位
- 从 5' 到 3' 方向合成 mRNA 的核苷酸;用 “+1” 表示
- 非模板链
- 不用于转录 mRNA 的 DNA 链;该链与 mRNA 相同,只是 DNA 中的 T 核苷酸被 mRNA 中的 U 核苷酸所取代
- 质粒
- 染色体外、共价封闭的圆形 DNA 分子,可能只含有一个或几个基因;常见于原核生物
- 启动者
- RNA 聚合酶和相关因子结合并启动转录的 DNA 序列
- 依赖于 Rho 的终止
- 在原核生物中,通过 RNA 聚合酶和 rho 蛋白在 DNA 模板上的 G 核苷酸中的相互作用来终止转录
- 与 Rho 无关
- 终止序列依赖性终止原核生物 mRNA 合成;由 mRNA 中形成发夹导致聚合酶停滞所致
- TATA 盒子
- 真核生物和原核生物中的保守启动子序列,有助于建立转录起始位点
- 模板链
- 指定互补 mRNA 分子的 DNA 链
- 转录气泡
- 允许转录 mRNA 的局部解开 DNA 区域
- 上游的
- 起始位点之前的核苷酸;一般而言,相对于mRNA上某个位点朝向5'末端的序列