
12.3: 继承法
- Page ID
- 202882

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 从遗传学和减数分裂事件的角度解释孟德尔的隔离定律和独立分类
- 使用分叉线法和概率规则来计算来自多个基因杂交的基因型和表型的概率
- 解释连锁和重组对配子基因型的影响
- 解释基因间上位效应的表型结果
孟德尔将豌豆植物实验的结果概括为四个假设,其中一些有时被称为 “定律”,描述了二倍体生物显性遗传和隐性遗传的基础。 正如你所了解的那样,存在更复杂的孟德尔主义扩展,它们表现出不同的 F 2 表型比 (3:1)。 尽管如此,这些定律总结了经典遗传学的基础知识。
成对的单位因子或基因
孟德尔首先提出,配对的遗传单位因子分别通过配子发生和受精期间配对因子的分离和重联而忠实地代代相传。 孟德尔在交叉具有对比特征的豌豆并发现隐性特征在F 2 代中重新浮出水面后,推断出遗传因子必须作为离散单位进行遗传。 这一发现与当时的看法相矛盾,即父母的特征融入了后代。
等位基因可以是显性的,也可以是隐性的
孟德尔的支配定律指出,在杂合子中,一个特征将掩盖相同特征的另一个特征的存在。 显性等位基因不是两个等位基因都构成表型,而是专门表达。 隐性等位基因将保持 “潜伏” 状态,但将以与显性等位基因传播相同的方式传播给后代。 隐性特征只能由具有两个该等位基因拷贝的后代表达(图\(\PageIndex{1}\)),而这些后代在自我交叉时会真正繁殖。
自从孟德尔对豌豆植物进行实验以来,其他研究人员发现支配定律并不总是正确的。 相反,已发现存在几种不同的继承模式。

等位基因的平等分离
孟德尔观察到具有对比性状的真正繁殖豌豆植物产生了所有表现显性状的F 1 代,而F 2 代则以 3:1 的比例表达了显性和隐性特征,因此提出了分离定律。 该定律规定,配对单位因子(基因)必须平均分离成配子,这样后代继承任一个因子的可能性相同。 对于单杂交杂交的 F 2 生成,可能产生以下三种可能的基因型组合:纯合显性、杂合或纯合隐性遗传。 由于杂合子可能来自两种不同的途径(从父母任一方获得一个显性等位基因和一个隐性等位基因),并且由于杂合子和纯合显性个体在表型上是相同的,因此该定律支持孟德尔观察到的 3:1 表型比率。 等位基因的平等分离是我们可以应用Punnett方块来准确预测具有已知基因型的父母的后代的原因。 孟德尔分离定律的物理基础是减数分裂的第一个分裂,在该分裂中,每个基因具有不同版本的同源染色体被分离成子核。 在孟德尔的一生中,科学界不了解染色体减数分离在有性生殖中的作用。
独立分类
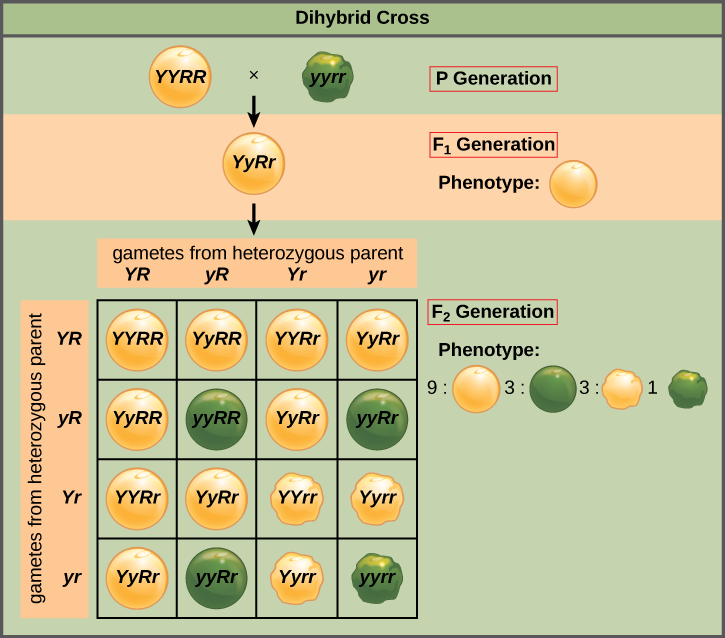
孟德尔的独立分类定律指出,在将等位基因分类为配子方面,基因不会相互影响,每个基因的所有可能的等位基因组合都同样有可能发生。 基因的独立分类可以用二杂交杂交来说明,二杂交是两个真正繁殖的亲本之间的杂交,它们表现出两个特征的不同特征。 以两种豌豆植物的种子颜色和种子质地的特征为例,一种有绿色的皱纹种子(yyrr),另一种有黄色的圆形种子(YYRR)。 因为每个亲本都是纯合的,所以隔离定律表明绿色/皱纹植物的配子都是你的,而黄色/圆形植物的配子都是 YR。 因此,F 1 代的后代都是 yyRR(图\(\PageIndex{2}\))。
练习
在豌豆植物中,紫花(P)比白花(p)占主导地位,黄豌豆(Y)在青豆(y)中占主导地位。 ppyY 和 ppy Y 豌豆植物之间的杂交可能有哪些基因型和表型? 你需要多少个方格才能对这个十字架进行 Punnett 方块分析?
- 回答
-
可能的基因型有 ppyY、ppyY、ppyY 和 ppyY。 前两种基因型将产生有紫花和黄豌豆的植物,而后两种基因型将产生有白花和黄豌豆的植物,每种表型的比例为 1:1。 你只需要一个 2×2 的 Punnett 方块(总共四个方格)即可进行此分析,因为其中两个等位基因是纯合的。
对于 F 2 世代,隔离定律要求每个配子获得 R 等位基因或 r 等位基因,以及 Y 等位基因或 y 等位基因。 独立分类定律指出,将 r 等位基因排序到的配子同样可能包含 Y 等位基因或 y 等位基因。 因此,当 yyrR 杂合子自交时,可能形成四种同样可能的配子,如下所示:YR、Y r 、y R 和 yr。 沿着 4×4 Punnett 方块的顶部和左侧排列这些配子(图\(\PageIndex{2}\))可以得出 16 种同样可能的基因型组合。 从这些基因型中,我们推断出表型比为 9 圆形/黄色:3 圆形/绿色:3 皱纹/黄色:1 皱纹/绿色(图\(\PageIndex{2}\))。 假设我们在样本量足够大的情况下进行了杂交,这些就是我们所期望的后代比率。
由于独立的分类和优势地位,9:3:3:1 的双杂交表型比率可以分解为两个 3:1 的比例,这是任何遵循显性和隐性模式的单杂交杂交的特征。 忽略种子颜色,只考虑上述 dihybrid 十字架中的种子质地,我们预计 F 2 世代后代中有四分之三是圆形的,四分之一会起皱。 同样,仅分离种子颜色,我们假设四分之三的 F 2 后代是黄色的,四分之一是绿色的。 纹理和颜色的等位基因排序是独立的事件,因此我们可以应用乘积规则。 因此,圆形和黄色 F 2 后代的比例预计为 (3/4) × (3/4) = 9/16,皱纹和绿色后代的比例预计为 (1/4) × (1/4) = 1/16。 这些比例与使用 Punnett 方块获得的比例相同。 圆形、绿色和有皱纹的黄色后代也可以使用乘积法则进行计算,因为这些基因型中的每一种都包含一种显性表型和一种隐性表型。 因此,每个比例的计算公式为 (3/4) × (1/4) = 3/16。
独立分类定律还表明,黄色、皱纹(yyRR)和绿色圆形(y yRR)亲本之间的交叉将产生与 YYRR x yr r 十字架中相同的 F 1 和 F 2 后代。
独立分类定律的物理基础也在于减数分裂 I,其中不同的同源对以随机方向排列。 每个配子可以包含父系和母系染色体(因此也包括其上的基因)的任意组合,因为四边形在中期平面上的取向是随机的。
分叉线法
当考虑两个以上的基因时,Punnett-Square 方法就会变得笨拙。 例如,检查涉及四个基因的交叉需要一个包含256个盒子的16×16网格。 手动输入每种基因型会非常麻烦。 对于更复杂的交叉点,首选分叉线和概率法。
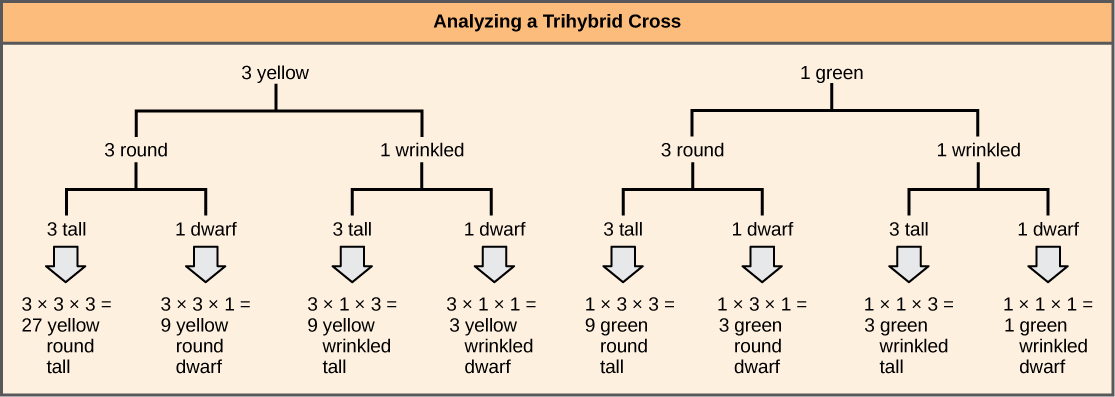
为了绘制由于 AABBCC 和 aabbcc 亲本之间交叉而产生的 F 1 杂合子之间的交叉线图,我们首先创建等于所考虑的基因数量的行,然后根据分叉线将每行中的等位基因分开单个单杂交杂交的概率(图\(\PageIndex{3}\))。 然后,我们将每条分叉路径上的值相乘以得出 F 2 后代概率。 请注意,此过程是产品规则的图表版本。 每条分叉路径上的值可以成倍增加,因为每个基因都是独立分类的。 对于 trihybrid 十字架,F 2 表型比为 27:9:9:9:3:3:1。
概率法
虽然分叉线法是一种跟踪交叉概率的图表化方法,但概率法给出了在没有额外视觉辅助的情况下预计会表现出每种表型(或基因型)的后代的比例。 两种方法都利用乘积法则并分别考虑每个基因的等位基因。 早些时候,我们使用分叉线法检查了三杂交交叉的表型比例;现在我们将使用概率法来检查具有更多基因的杂交的基因型比例。
对于三混合交叉来说,写出分叉线方法很乏味,尽管不像使用 Punnett-Square 方法那么繁琐。 但是,为了充分证明概率法的力量,我们可以考虑特定的遗传计算。 例如,对于所有四个基因都是杂合子的个体之间的四杂交杂交叉体,其中所有四个基因都以显性和隐性模式独立排序,那么预计所有四个等位基因的后代中有多少比例是纯合隐性遗传? 我们可以使用概率法,而不是写出所有可能的基因型。 我们知道,对于每个基因,纯合隐性后代的比例将为1/4。 因此,将四个基因中每个基因的这个分数乘以 (1/4) × (1/4) × (1/4),我们确定1/256的后代将是四倍纯合隐性遗传。
对于同一个四杂交杂交系,在所有四个基因座上具有显性表型的后代的预期比例是多少? 我们可以使用表型比例来回答这个问题,但让我们用艰难的方法来回答这个问题——使用基因型比例。 这个问题询问了 1) 在 A 处纯合占优势或在 A 处为杂合子,2) 在 B 处为纯合子或 B 处为杂合子等的后代的比例。 在每种情况下注明 “或” 和 “和” 可以清楚地表明在哪里适用总和和和和和规则。 A 处纯合占优势的概率为 1/4,A 处杂合子的概率为 1/2。 使用总和规则,纯合子或杂合子的概率为 1/4 + 1/2 = 3/4。 其他每个基因可以用相同的方式获得相同的概率,因此,使用乘积法则,A、B、C 和 D 处显性表型的概率等于 3/4×3/4×3/4 或 27/64。 如果你不确定如何合并概率,那么回到分叉线方法应该很清楚。
多杂交施肥规则
预测特定杂交后代的基因型和表型是检验你对孟德尔遗传学知识的最佳方法。 鉴于多杂交叉遵循独立分类并遵循显性和隐性模式,因此存在几种通用规则;在进行遗传计算时,您可以使用这些规则来检查结果(表\(\PageIndex{1}\))。 要应用这些规则,首先必须确定 n,即杂合基因对的数量(每个分离两个等位基因的基因数量)。 例如,aa BB 和 a aBb 杂合子之间的交叉点的 n 为 2。 相比之下,AaBB 和 aaBB 之间的交叉点的 n 为 1,因为 A 不是杂合子。
| 一般规则 | 杂合基因对的数量 |
|---|---|
| 不同 F 1 配子的数量 | 2 n |
| 不同 F 2 基因型的数量 | 3 n |
| 鉴于显性遗传和隐性遗传,不同 F 2 表型的数量 | 2 n |
关联基因违反了独立分类法
尽管孟德尔的所有豌豆特征都按照独立分类定律行事,但我们现在知道有些等位基因组合不是相互独立继承的。 位于单独的非同源染色体上的基因将始终独立排序。 但是,每条染色体包含数百或数千个基因,这些基因在染色体上呈线性组织,就像绳子上的珠子一样。 等位基因向配子的分离可能受到连锁的影响,在连锁中,位于同一染色体上物理上彼此靠近的基因更有可能成对遗传。 但是,由于重组或 “交叉” 的过程,同一染色体上的两个基因有可能独立运行,或者好像它们没有关联。 要理解这一点,让我们考虑一下基因连锁和重组的生物学基础。
同源染色体以相同的线性顺序具有相同的基因。 同源染色体对上的等位基因可能有所不同,但它们对应的基因却不一样。 在为减数分裂的第一个分裂做准备时,同源染色体会复制和突触。 就像同系物上的基因相互对齐。 在这个阶段,同源染色体的片段交换遗传物质的线性片段(图\(\PageIndex{4}\))。 这个过程被称为重组或交叉,这是一个常见的遗传过程。 由于基因在重组过程中是排列的,因此基因顺序不会改变。 相反,重组的结果是母系和父系等位基因合并到同一个染色体上。 在给定的染色体上,可能会发生几次重组事件,导致等位基因的大规模洗牌。
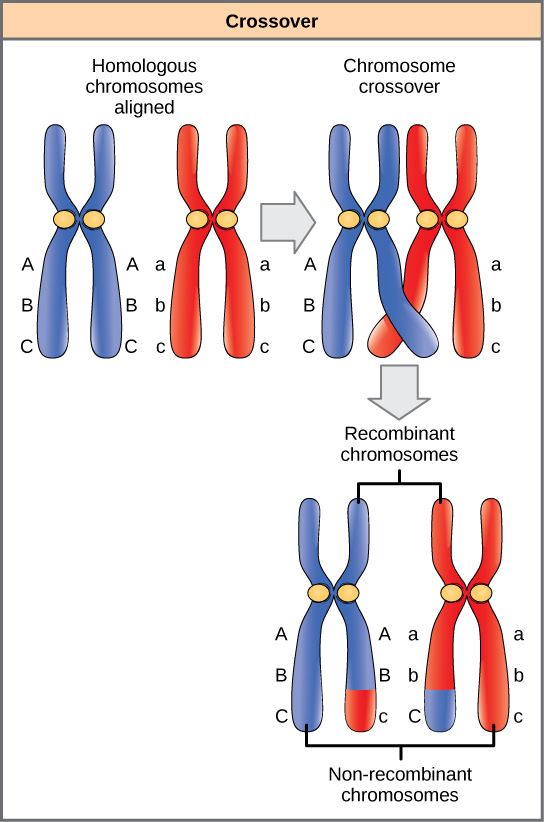
当两个基因位于同一条染色体上非常接近时,它们被认为是相互关联的,它们的等位基因往往通过减数分裂一起传播。 举例来说,想象一下涉及花朵颜色和植物高度的双杂交叉体,其中染色体上的基因彼此相邻。 如果一条同源染色体具有高大植物和红花的等位基因,而另一条染色体具有短植物和黄花的基因,那么当配子形成时,高大的和红色的等位基因将一起变成配子,而短和黄色的等位基因将进入其他配子。 它们之所以被称为亲本基因型,是因为它们是从产生配子的个体的父母那里完好无损地继承的。 但是,与基因在不同的染色体上不同的是,不会有具有高和黄色等位基因的配子,也没有带有短和红色等位基因的配子。 如果你用这些配子创建 Punnett 方块,你会发现孟德尔对双混合交叉结果的 9:3:3:1 的经典预测不适用。 随着两个基因之间距离的增加,它们之间发生一次或多次交叉的可能性增加,并且这些基因的行为更像它们在不同的染色体上。 遗传学家使用重组配子(不像亲本的配子)的比例来衡量染色体上基因之间的距离。 利用这些信息,他们为包括人类在内的经过充分研究的生物绘制了染色体上精心制作的基因图谱。
孟德尔的开创性出版物没有提及联系,许多研究人员质疑他是否遇到过联系,但选择不发表这些十字架,因为担心它们会使他的独立分类假设失效。 园豌豆有七条染色体,有人认为他选择的七个特征并非巧合。 但是,即使他检查的基因不在单独的染色体上,他也可能因为重组的广泛洗牌效应而没有观察到连锁反应。
科学方法连接:检验独立分类的假设
为了更好地了解孟德尔实验所投入的大量劳动力和聪明才智,请继续研究孟德尔的双杂交十字架之一。
问题:双杂交十字架的后代会是什么?
背景:假设豌豆植物在一个生长季节成熟,你可以进入一个大花园,在那里你可以种植成千上万的豌豆植物。 有几种真正繁殖的植物具有以下两对特征:带有膨胀豆荚的高大植物和带有收缩豆荚的矮植物。 在植物成熟之前,你要从十字架中高大/膨胀的植物中移除产生花粉的器官,以防止自我受精。 植物成熟后,通过将花粉从矮树/收缩植物转移到高/膨胀植物的柱头上,手动交叉植物。
假设:两个特征对将根据孟德尔定律独立排序。 当真正繁殖的父母交叉时,所有 F 1 后代都很高并且有膨胀的豆荚,这表明高大和膨胀的特征分别比矮人和收缩性状占主导地位。 F 1 杂合子的自交会产生 2,000 F 2 后代。
检验假设:由于每个特征对都是独立排序的,因此 tall: dwarf 和 inflated: constricted 的比率预计均为 3:1。 高/矮人特质对被称为 t/t,膨胀/收缩特质对被指定为 i/i。 因此,F 1 代的每个成员都有 t tiI 的基因型。 构造一个类似于 Figure 的网格\(\PageIndex{2}\),你可以在其中交叉两个 ttiI 个体。 每个人可以捐赠两种特征的四种组合:T I、Ti、Ti 或 ti,这意味着后代基因型有 16 种可能性。 由于 T 和 I 等位基因占主导地位,任何拥有其中一个或两个等位基因的个体都将分别表现出较高或膨胀的表型,无论他们是否还有 t 或 i 等位基因。 只有 tt 或 ii 的个体才能分别表达矮人和收缩的等位基因。 如图所示\(\PageIndex{5}\),你预测你会观察到以下后代比例:tall/inflated: tall/constricted: dwarf/inflated: dwarf/constricted 以 9:3:3:1 的比例收缩。 从网格中注意到,单独考虑高/矮人和膨胀/收缩特征对时,它们都是以 3:1 的比例继承的。
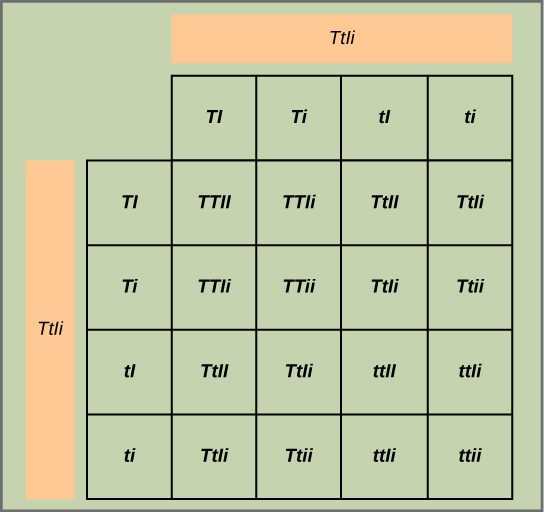
检验假设:你穿过矮人和高大的植物,然后自我穿越后代。 为了获得最佳效果,对数百甚至数千株豌豆植物重复使用这种方法。 在十字架和种植植物时应采取哪些特别预防措施?
分析您的数据:您在 F 2 世代中观察到以下植物表型:2706 个高/膨胀、930 个高/收缩、888 个矮人/膨胀和 300 个矮人/收缩。 将这些发现降低到一定比例,并确定它们是否符合孟德尔定律。
得出结论:结果是否接近预期的 9:3:3:1 表型比率? 结果支持预测吗? 鉴于等位基因随机分离成配子,如果使用的植物少得多,会观察到什么? 试着想象种那么多豌豆植物,并考虑实验错误的可能性。 例如,如果有一天风很大,会发生什么?
鼻出血
孟德尔对豌豆植物的研究表明,个体表型的总和受基因(或他所说的单位因子)的控制,因此每个特征都由单个基因明显而完全地控制。 实际上,单一可观察的特征几乎总是受到多个基因(每个基因有两个或多个等位基因)协同作用的影响。 例如,至少有八个基因会影响人类的眼睛颜色。
在某些情况下,几个基因可以为共同表型的各个方面做出贡献,而它们的基因产物不会直接相互作用。 例如,就器官发育而言,基因可以按顺序表达,每个基因都增加了器官的复杂性和特异性。 基因可能以互补或协同的方式发挥作用,因此需要同时表达两个或多个基因才能影响表型。 基因也可能相互对立,一个基因会改变另一个基因的表达。
在鼻出血中,基因之间的相互作用是拮抗的,因此一个基因掩盖或干扰另一个基因的表达。 “Epistasis” 这个词由希腊语词根构成,意思是 “站立”。 据说被掩盖或沉默的等位基因比起掩盖作用的上位基因低估了。 通常,鼻出血的生化基础是一种基因途径,其中一个基因的表达取决于该途径中先于或之后的基因的功能。
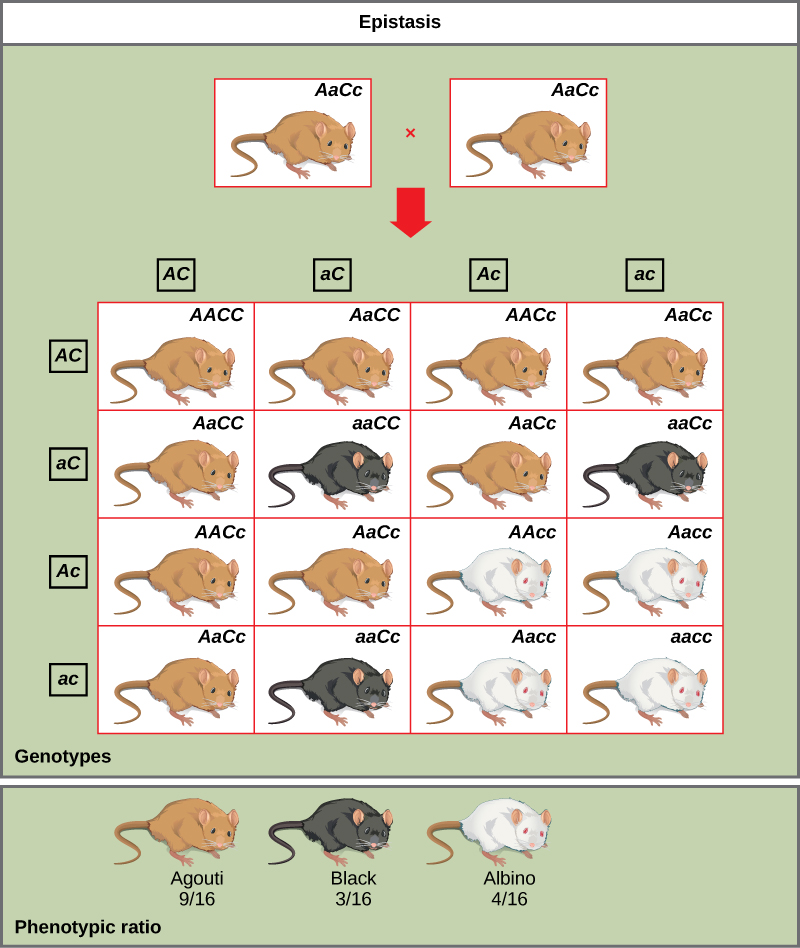
鼻出血的一个例子是小鼠的色素沉着。 野生型外套颜色agouti(AA)在纯色毛皮(aa)中占主导地位。 但是,色素产生需要一个单独的基因(C)。 无论轨迹A存在什么等位基因,在该轨迹上具有隐性 c 等位基因的小鼠都无法产生色素并且是白化病(图\(\PageIndex{6}\))。 因此,Aacc、Aac c 和 aac c 的基因型都产生相同的白化表型。 两个基因(aacC x aac C)的杂合子之间的交叉将产生表型比为 9 agouti: 3 纯色:4 白化病的后代(图\(\PageIndex{6}\))。 在这种情况下,C 基因是 A 基因的上位基因。
当显性等位基因掩盖了单独基因的表达时,也可能发生鼻出血。 夏季南瓜中的水果颜色就是这样表达的。 W 基因 (w w) 的纯合隐性表达与 Y 基因(YY 或 Y y)的纯合显性或杂合表达相结合,会生成黄色果实,而 wwyy 基因型会产生绿色果实。 但是,如果 W 基因的显性拷贝以纯合或杂合形式存在,则无论Y等位基因如何,夏季南瓜都会产生白色果实。 两个基因(WWYY × WWYY)的白色杂合子之间的杂交将产生表型比为 12 白色:3 黄色:1 绿色的后代。
最后,鼻出血可能是互惠的,因此任何一个基因在以显性(或隐性)形式存在时都表现出相同的表型。 在牧羊人的钱包植物(Capsella bursa-pastoris)中,种子形状的特征由两个具有显性上位关系的基因控制。 当基因 A 和 B 都是纯合隐性遗传(aabb)时,种子是卵形的。 如果这两个基因中的任何一个都存在显性等位基因,则结果是三角种子。 也就是说,除了 aabb 以外的所有可能的基因型都会产生三角种子,而两个基因(aaBB x aabB)的杂合子之间的交叉将产生表型比为 15 三角形:1 卵形的后代。
在研究遗传学问题时,请记住,任何导致表型比总数为16的单一特征都是两个基因相互作用的典型特征。 回想一下孟德尔双杂交叉的表型遗传模式,它考虑了两个非相互作用的基因—— 9:3:1。 同样,我们预计相互作用的基因对也将表现出以16个部分表示的比率。 请注意,我们假设相互作用的基因没有关联;它们仍在独立分类成配子。
摘要
孟德尔假设基因(特征)是作为成对的等位基因(特征)遗传的,这些等位基因(特征)呈显性和隐性模式。 等位基因分离成配子,因此每个配子同样有可能获得二倍体个体中存在的两个等位基因中的任何一个。 此外,基因被分成相互独立的配子。 也就是说,等位基因通常不太可能分离成具有另一个基因的特定等位基因的配子。 当所讨论的基因位于不同的染色体上或在同一条染色体上彼此相距较远时,dihybrid 杂交会表现出独立的分类。 对于涉及两个以上基因的杂交,使用分叉线或概率方法来预测后代的基因型和表型,而不是 Punnett 方块。
尽管在减数分裂期间染色体独立分类为配子,但孟德尔的独立分类定律指的是基因,而不是染色体,单条染色体可能携带超过1,000个基因。 当基因位于同一条染色体上很近时,它们的等位基因往往会一起遗传。 这导致后代比例违反了孟德尔的独立分类定律。 但是,重组用于交换同源染色体上的遗传物质,因此母系和父系等位基因可以在同一个染色体上重组。 这就是为什么给定染色体上的等位基因并不总是一起遗传的原因。 重组是发生在染色体任何地方的随机事件。 因此,由于中间的染色体空间中发生了重组事件,同一染色体上相距很远的基因很可能仍会独立分组。
无论它们是否独立排序,基因都可能在基因产物层面上相互作用,因此一个基因的等位基因的表达掩盖或改变了另一个基因的等位基因的表达。 这被称为鼻出血。
词汇表
- dihyb
- 两个真正育种的父母之间交叉的结果,他们对两个特征表现出不同的特征
- 鼻出血
- 基因之间的拮抗相互作用,使一个基因掩盖或干扰另一个基因的表达
- 支配法则
- 在杂合子中,一个特征会掩盖相同特征的另一个特征的存在
- 独立分类法则
- 在将等位基因分类为配子方面,基因不会相互影响;所有可能的等位基因组合都同样有可能发生
- 隔离法则
- 配对的单位因子(即基因)平均分离成配子,因此后代继承任何因子组合的可能性相同
- 联系
- 一种现象,即位于同一染色体上彼此非常接近的等位基因更有可能一起遗传