
12.2:特征和特征
- Page ID
- 202880

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 解释显性和隐性基因系统中基因型和表型之间的关系
- 开发一个 Punnett 方块来计算单杂交杂交中基因型和表型的预期比例
- 解释测试交叉的目的和方法
- 识别非孟德尔遗传模式,例如不完全占主导地位、共占优势、隐性致死、多个等位基因和性别联系
孟德尔在豌豆植物中评估的七个特征均表示为两个版本或性状之一。 特征的物理表达是通过表达染色体上携带的基因来实现的。 豌豆的基因组成由每条染色体的两个相似或同源的副本组成,每个亲本各一个。 每对同源染色体具有相同的基因线性顺序。 换句话说,豌豆是二倍体生物,因为它们每条染色体有两个拷贝。 许多其他植物和几乎所有动物也是如此。 二倍体生物利用减数分裂产生单倍体配子,其中包含每条同源染色体的一份副本,这些染色体在受精时结合形成二倍体合子。
对于单个基因控制单一特征的情况,二倍体生物有两个遗传拷贝,它们可能编码也可能不会编码该特征的相同版本。 由突变产生并存在于同源染色体上相同相对位置的基因变异称为等位基因。 孟德尔研究了只有两种等位基因形式的基因的遗传,但在自然群体中,任何给定基因都会遇到两个以上的等位基因。
表型和基因型
二倍体生物体中给定基因的两个等位基因被表达并相互作用以产生物理特征。 生物体表达的可观察特征被称为其表型。 生物体的潜在基因构成,包括物理上可见的和未表达的等位基因,被称为其基因型。 孟德尔的杂交实验证明了表型和基因型之间的差异。 当一个亲本有黄豆荚而另一个有绿色豆荚的真育植物进行交叉受精时,所有 F 1 杂交后代都有黄色的豆荚。 也就是说,杂交后代在表型上与有黄豆荚的真正繁殖的亲本相同。 但是,我们知道,父母捐赠的带有绿色豆荚的等位基因并不仅仅是因为它在某些 F 2 后代身上重新出现而丢失。 因此,F 1 植物在基因型上一定与有黄色豆荚的亲本不同。
孟德尔在实验中使用的P 1 植物对于他正在研究的特征来说都是纯合的。 在给定基因或轨迹上纯合的二倍体生物在其同源染色体上具有该基因的两个相同的等位基因。 孟德尔的亲本豌豆植物总是真正繁殖的,因为生成的两个配子都具有相同的特征。 当具有对比性状的 P 1 植物进行交叉受精时,所有后代都具有对比性状的杂合子,这意味着它们的基因型反映出它们对正在检查的基因有不同的等位基因。
显性及隐性等位基因
我们对纯合和杂合生物的讨论使我们明白了为什么 F 1 杂合子后代与其中一个亲本相同,而不是同时表达两个等位基因。 在豌豆植物的所有七个特征中,两个对比鲜明的等位基因中有一个是显性的,另一个是隐性的。 孟德尔称显性等位基因为表达单位因子;隐性等位基因被称为潜在单位因子。 我们现在知道这些所谓的单位因子实际上是同源染色体对上的基因。 对于以显性和隐性模式表达的基因,纯合显性和杂合生物看起来相同(也就是说,它们将具有不同的基因型但表型相同)。 隐性等位基因只能在纯合隐性遗传个体中观察到(表\(\PageIndex{1}\))。
| 显性特征 | 隐性特征 |
|---|---|
| 软骨发育不全 | 白化病 |
| Brachydactyly | 囊性纤维化 |
| 亨廷顿氏病 | 杜兴氏肌营养不良症 |
| 马凡综合症 | 半乳糖血症 |
| 神经纤维瘤病 | 苯丙酮尿症 |
| 寡妇的巅峰 | 镰状细胞贫血 |
| 毛茸茸的头发 | 泰萨克斯病 |
有几种提及基因和等位基因的惯例。 出于本章的目的,我们将使用基因相应显性特征的第一个字母来缩写基因。 例如,紫罗兰是豌豆植物花色的主要特征,因此花色基因将缩写为 V(请注意,习惯上使用斜体表示基因名称)。 此外,我们将分别使用大写和小写字母来表示显性等位基因和隐性等位基因。 因此,我们将紫罗兰花的纯合优势豌豆植物的基因型称为 VV,一种以白花为vv的纯合隐性豌豆植物,以及一种以紫罗兰花为 Vv 的杂合豌豆植物的基因型。
单杂交十字架的 Punnett Square 方法
当两个真正育种的父母之间发生受精时,这些亲本只有一个特征不同,则该过程称为单杂交杂交,由此产生的后代是单杂交种。 孟德尔进行了七次单杂交杂交,涉及每个特征的对比特征。 根据他在F 1 和F 2 代中的结果,孟德尔假设单杂交杂交中的每个父母都为每个后代贡献了两个配对单位因子中的一个,每种可能的单位因子组合的可能性都相同。
要演示单杂交杂交,可以考虑使用黄豌豆种子和绿豌豆种子的真正繁殖豌豆植物的案例。 主要种子颜色为黄色;因此,黄色种子的植物的亲本基因型分别为 Y Y和具有绿色种子的植物的亲本基因型为yy。 可以绘制一个由英国遗传学家雷金纳德·庞内特设计的 Punnett 方块,它运用概率规则来预测遗传交叉或交配的可能结果及其预期频率。 为了准备一个 Punnett 方块,所有可能的亲本等位基因组合都列在网格的顶部(对于一个亲本)和侧面(对于另一个亲本),表示它们的减数分离为单倍体配子。 然后在表格的方框中组合出卵子和精子的组合,以显示哪些等位基因正在结合。 然后,每个盒子代表这种交配可能产生的合子或受精卵的二倍体基因型。 由于每种可能性相同,因此可以从 Punnett 方块中确定基因型比率。 如果已知遗传模式(显性或隐性),则也可以推断出表型比率。 对于由两个真正繁殖的亲本组成的单杂交杂交,每个亲本贡献一种等位基因。 在这种情况下,只有一种基因型是可能的。 所有后代都是 Yy 并且有黄色的种子(图\(\PageIndex{1}\))。
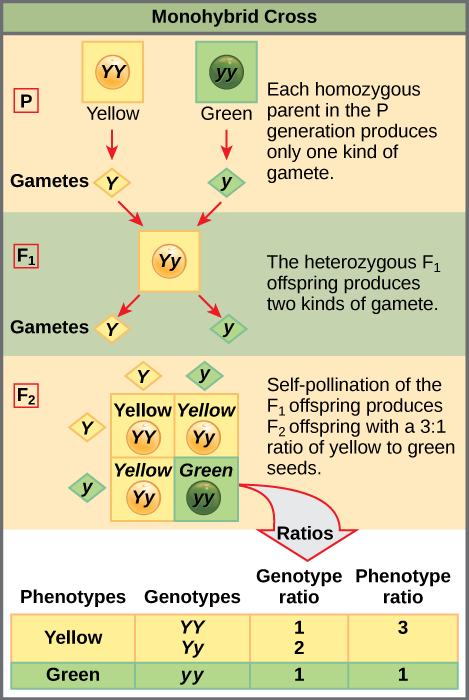
其中一个 Y y杂合子后代的自交可以用2×2 Punnett方块表示,因为每个父母都可以捐赠两个不同的等位基因中的一个。 因此,后代可能有四种等位基因组合中的一种:YY、Yy、yy 或 yy(图\(\PageIndex{1}\))。 请注意,有两种获得 Y y 基因型的方法:a Y 来自卵子,a y 来自精子,或 a y 来自卵子,一个 Y 来自精子。 必须将这两种可能性都考虑在内。 回想一下,孟德尔的豌豆植物特征在倒数交叉中的表现方式相同。 因此,这两种可能的杂合组合产生的后代尽管显性和隐性等位基因来自不同的父母,但它们在基因型和表型上是相同的。 它们组合在一起。 由于受精是一个随机事件,我们预计每种组合的可能性相同,并且后代的 YY: Y y: y y基因型比例为 1:2:1(图\(\PageIndex{1}\))。 此外,由于 YY 和 Yy 后代的种子是黄色的,表型上是相同的,应用概率求和法则,我们预计后代的表型比为 3 黄色:1 绿色。 事实上,在样本量较大的情况下,孟德尔观察到每代 F 2 中大约有这个比率,这是个体特征的交叉产生的。
孟德尔通过进行 F 3 交叉来验证这些结果,他在交叉中自我交叉表达占主导地位和隐性的 F 2 植物。 当他自我交叉表达绿色种子的植物时,所有后代都有绿色种子,这证实了所有绿色种子的纯合基因型均为 yy。 当他自我交叉表达黄色种子的 F 2 植物时,他发现三分之一的植物是真实繁殖的,三分之二的植物以 3:1 的黄绿色种子比例分离。 在这种情况下,真正的育种植物具有纯合子(YY)基因型,而分离植物对应于杂合子(Yy)基因型。 当这些植物自我受精时,结果就像 F 1 自受精的十字架一样。
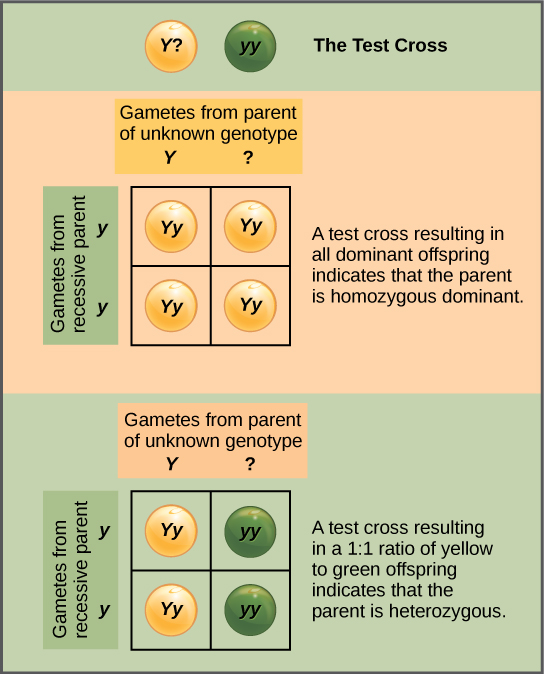
测试十字架区分显性表型
除了预测已知纯合子或杂合父母之间杂交的后代外,孟德尔还开发了一种方法来确定表达显性特征的生物是杂合子还是纯合子。 这种技术被称为测试十字架,至今仍被动植物育种者使用。 在测试十字架中,表达显性的生物体与具有相同特征的纯合隐性生物交叉。 如果表达显性特征的生物体是纯合子,则所有 F 1 后代都是表达显性特征的杂合子(图\(\PageIndex{2}\))。 或者,如果显性表达生物是杂合子,则 F 1 后代的杂合子和隐性纯合子的比例将为 1:1(图\(\PageIndex{2}\))。 检验交叉进一步证实了孟德尔的假设,即成对的单位因子平均分离。
练习
在豌豆植物中,圆豌豆(R)比起皱豌豆(r)占主导地位。 你在带有皱纹豌豆(基因型 rr)的豌豆植物和具有圆豌豆的未知基因型植物之间进行交叉测试。 你最终会得到三株植物,它们都有圆豌豆。 从这些数据中,你能分辨出圆豌豆亲本植物是纯合占主导地位还是杂合子的? 如果圆豌豆亲本植物是杂合子,那么随机抽取的 3 个后代豌豆全部变圆的概率是多少?
- 回答
-
你无法确定植物是纯合还是杂合子,因为数据集太小:偶然地,即使存在隐性基因,所有三种植物也可能只获得了显性基因。 如果圆豌豆亲本是杂合子,则三个后代豌豆的随机样本全部为圆形的概率为八分之一。
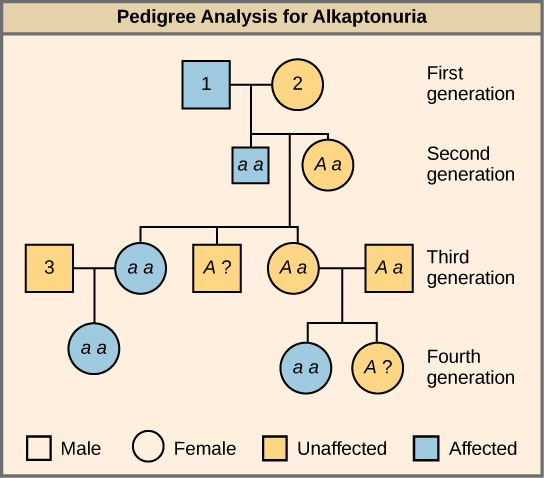
许多人类疾病都是遗传性的。 家庭中有些人患有隐性遗传疾病,健康的人可能想知道他或她是否有致病基因,以及将这种疾病传染给他或她的后代的风险有多大。 当然,在人体中进行交叉测试是不道德的,也是不切实际的。 相反,遗传学家使用血统分析来研究人类遗传疾病的遗传模式(图\(\PageIndex{3}\))。
练习
标记为 1、2 和 3 的个体的基因型是什么?
- 回答
-
个体 1 的基因型为 aa。 个体 2 的基因型为 Aa。 个体 3 的基因型为 Aa。
支配和退缩的替代方案
孟德尔对豌豆植物的实验表明:(1)每个基因存在两个 “单位” 或等位基因;(2)等位基因在每一代中都保持其完整性(不混合);(3)在存在显性等位基因的情况下,隐性等位基因是隐藏的,对表型没有任何贡献。 因此,隐性等位基因可以 “携带”,而不能由个体表达。 这种杂合个体有时被称为 “携带者”。 对其他动植物的进一步遗传研究表明,复杂性要大得多,但孟德尔遗传学的基本原理仍然成立。 在接下来的章节中,我们将讨论孟德尔主义的一些扩展。 如果孟德尔选择了一个表现出这些遗传复杂性的实验系统,他可能不明白他的结果意味着什么。
统治不完整
孟德尔的研究结果,即特征是作为显性遗传和隐性对遗传,这与当时的后代表现出父母特征混合的观点相矛盾。 但是,杂合子表型偶尔看起来确实介于两个父母之间。 例如,在 snapdragon Antirrhinum majus(图\(\PageIndex{4}\))中,有白花的纯合父母(C W C W)和有红花的纯合亲本(C R C R)之间的杂交将产生粉红色的后代花(C R C W)。 (请注意,孟德尔扩展使用不同的基因型缩写,以区分这些模式与简单的优势和隐性。) 这种遗传模式被描述为不完全的优势,表示两个对比的等位基因的表达,使个体表现出中间表型。 与白花的等位基因相比,红花的等位基因不完全占主导地位。 但是,杂合子自交叉的结果仍然可以预测,就像孟德尔显性交叉和隐性交叉一样。 在这种情况下,基因型比率为 1 C R C R: 2 C R C W: 1 C W C W,红色:pink: w hite 的表型比率为 1:2:1。

共同支配地位
不完全优势的一种变体是共显性,其中具有相同特征的两个等位基因在杂合子中同时表达。 共同优势的一个例子是人类的明尼苏达血型。 M 和 N 等位基因以存在于红细胞表面的 M 或 N 抗原的形式表达。 纯合子(L M L M 和 L N L N)表达 M 或 N 等位基因,杂合子(L M L N)平等表达两个等位基因。 在表达共显性状的杂合子之间的自我交叉中,三种可能的后代基因型在表型上是不同的。 但是,孟德尔单杂交杂交的 1:2:1 基因型比率特征仍然适用。
多个等位基因
孟德尔暗示,给定基因只能存在两个等位基因,一个显性等位基因,一个是隐性基因。 我们现在知道这太简单了。 尽管个体人类(以及所有二倍体生物)只能有给定基因的两个等位基因,但在种群层面上可能存在多个等位基因,因此可以观察到两个等位基因的许多组合。 请注意,当同一个基因存在许多等位基因时,惯例是将野生动物中最常见的表型或基因型表示为野生型(通常缩写为 “+”);这被认为是标准或规范。 所有其他表型或基因型均被视为该标准的变体,这意味着它们与野生型有所不同。 该变异体可能隐性或显性于野生型等位基因。
多个等位基因的一个例子是兔子的外套颜色(图\(\PageIndex{5}\))。 在这里,c 基因存在四个等位基因。 野生型版本 C + C + 表示为棕色毛皮。 龙猫表型 c c h 表示为黑尖的白色毛皮。 喜马拉雅表型 c h h 四肢有黑色毛皮,其他地方有白色皮毛。 最后,白化病或 “无色” 表型 cc 表示为白色毛皮。 如果有多个等位基因,则可能存在优势层次结构。 在这种情况下,野生型等位基因比所有其他等位基因占主导地位,龙猫在喜马拉雅和白化病上占主导地位,喜马拉雅比白化病占主导地位。 通过观察每个可能的杂合子后代的表型,揭示了这种层次结构或等位基因系列。
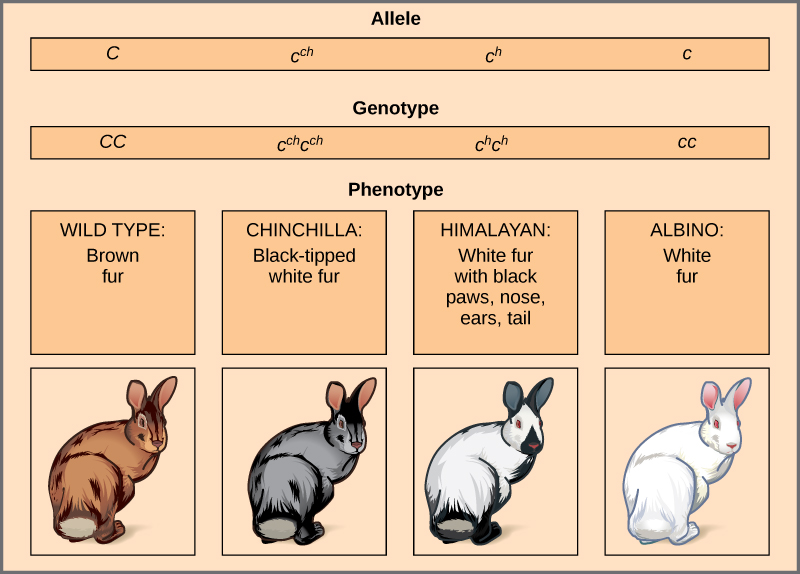
野生型表型相对于所有其他突变体的完全优势通常是特定基因产物 “剂量” 的影响,因此野生型等位基因提供正确数量的基因产物,而突变等位基因不能。 对于兔子的等位基因系列,野生型等位基因可能提供给定剂量的毛皮色素,而突变体提供的剂量较少或根本不提供。 有趣的是,喜马拉雅表型是等位基因的结果,该等位基因产生温度敏感的基因产物,这种产物只能在兔子身体较冷的四肢产生色素。
或者,一个突变等位基因可以比所有其他表型(包括野生型)占主导地位。 当突变等位基因以某种方式干扰遗传信息时,可能会发生这种情况,因此,即使是具有一个野生型等位基因拷贝的杂合子也能表达突变表型。 突变等位基因可以干扰的一种方法是增强野生型基因产物的功能或改变其在体内的分布。 这方面的一个例子是果蝇中的 Antennapedia 突变(图\(\PageIndex{6}\))。 在这种情况下,突变等位基因扩大了基因产物的分布,因此,Anten napedia h eterozygote 在触角应位于的头上长出腿。

进化联系:多个等位基因使疟疾寄生虫具有耐药性
疟疾是一种人类寄生虫病,由受感染的雌性蚊子传播,包括冈比亚按蚊(图\(\PageIndex{7}\) a),其特征是周期性高烧、发冷、流感样症状和严重贫血。 恶性疟原虫和间日疟原虫是疟疾最常见的病原体,恶性疟原虫是最致命的(图\(\PageIndex{7}\) b)。 如果得到及时和正确的治疗,恶性疟原虫疟疾的死亡率为0.1%。 但是,在世界某些地区,这种寄生虫已经对常用的疟疾治疗产生了抗药性,因此最有效的疟疾治疗方法可能因地理区域而异。

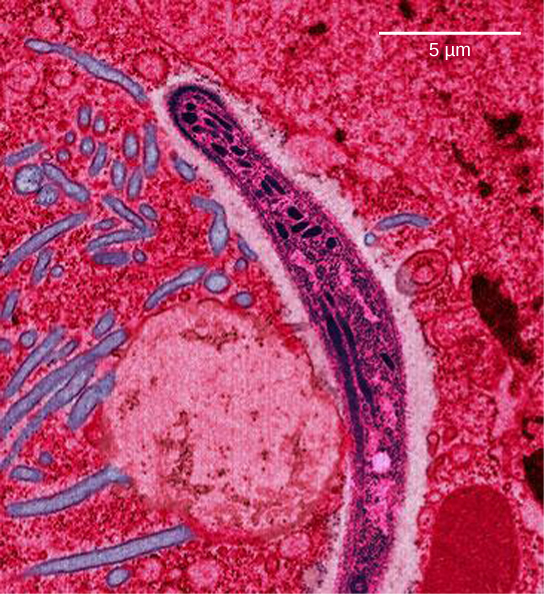
在东南亚、非洲和南美,恶性疟原虫对抗疟疾药物氯、甲氟喹和磺胺多辛-乙胺嘧啶产生了耐药性。 恶性@@ 疟原虫在对人类具有传染性的生命阶段是单倍体,它已经进化出了 dhps 基因的多个耐药突变等位基因。 不同程度的磺胺多辛耐药性与每个等位基因有关。 作为单倍体,恶性疟原虫只需要一个耐药等位基因即可表达这种特征。
在东南亚,dhp s基因的不同耐磺胺多辛等位基因分布在不同的地理区域。 这是一种常见的进化现象,之所以发生这种现象,是因为耐药突变体出现在人群中,并且与其他恶性疟原虫分离株近距离杂交。 在磺胺多辛被广泛用作非处方疟疾药物的地区,耐磺胺多辛的寄生虫造成了巨大的人类苦难。 与在感染周期内大量繁殖的病原体一样,恶性疟原虫在常用抗疟药物的选择性压力下进化相对较快(超过十年左右)。 出于这个原因,科学家必须不断努力开发新的药物或药物组合来对抗全球疟疾负担。 1
X 关联特征
在人类以及许多其他动物和某些植物中,个体的性别由性染色体决定。 性染色体是一对非同源染色体。 到目前为止,我们只考虑了非性染色体或常染色体之间的遗传模式。 除了 22 对同源的常染色体外,人类雌性有一对同源的 X 染色体,而人类雄性有一对 XY 染色体。 尽管 Y 染色体包含与 X 染色体相似的小区域,因此它们可以在减数分裂期间配对,但 Y 染色体要短得多,包含的基因也少得多。 当正在检查的基因存在于X染色体上但不存在于Y染色体上时,据说该基因与X染色体相连。
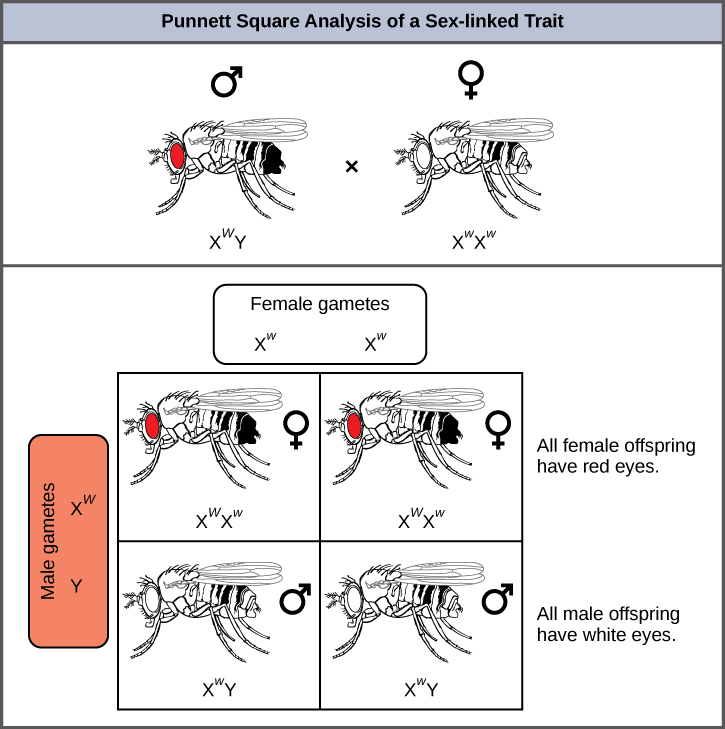
果蝇的眼睛颜色是最早被鉴定的X连锁特征之一。 托马斯·亨特·摩根在 1910 年将这个特征映射到 X 染色体上。 和人类一样,雄性果蝇有一对 XY 染色体,雌性是 XX。 在苍蝇中,野生型的眼睛颜色为红色(X W),以白眼色(X w)为主(图\(\PageIndex{8}\))。 由于眼睛颜色基因的位置,互惠杂交不会产生相同的后代比例。 据说雄性是 hemizygous,因为它们对任何 X 连锁特征都只有一个等位基因。 Hemizygosity 使得对支配地位和后退的描述对于 XY 男性来说无关紧要。 雄性@@ 果蝇在 Y 染色体上缺少第二个等位基因拷贝;也就是说,它们的基因型只能是 X W Y 或 X w Y。相比之下,雌性有这个基因的两个等位基因拷贝,可以是 X W X W 、X W X w 或 X w X w。

在 X 连锁交叉中,F 1 和 F 2 后代的基因型取决于隐性特征在 P 1 代中由雄性还是雌性表达。 关于果蝇的眼睛颜色,当 P 1 雄性表示白眼表型而雌性呈纯合红眼时,F 1 代的所有成员都表现出红眼睛(图\(\PageIndex{9}\))。 F 1 雌性是杂合子(X W X w),雄性都是 X W Y,它们从纯合显性 P 1 雌性那里获得了 X 染色体,从 P 1 雄性那里获得了 Y 染色体。 随后 X W X w 雌性和 X W Y 雄性之间的交叉只会产生红眼雌性(X W X W 或 X WX w 基因型)以及红眼和白眼雄性(具有 X W Y 或 X w Y 基因型)。 现在,考虑一下纯合的白眼雌性和红眼的雄性之间的交叉。 F 1 一代将只展示杂合红眼雌性(X W X w)和白眼雄性(X w Y)。 F 2 雌性中有一半是红眼的(X W X w),一半是白眼的(X w X w)。 同样,F 2 雄性中有一半是红眼(X W Y),一半是白眼(X w Y)。
练习
白眼雄性和雌性之间的杂合红眼颜色会产生多大比例的后代?
- 回答
-
一半的雌性后代是杂合子(X W X w),有红眼睛,一半是纯合隐性遗传(X w X w),有白眼睛。 一半的雄性后代将是 hemizygous 占主导地位(X W Y),是的,红色,一半是白眼睛的 hemizygous 隐性遗传(X w Y)。
果蝇遗传学的发现可以应用于人类遗传学。 当女性父母因隐性 X 连锁特征纯合时,她会将该特征传递给百分之百的后代。 因此,她的雄性后代注定要表现出这种特征,因为他们将继承父亲的Y染色体。 在人类中,某些疾病(某些形式的色盲、血友病和肌肉萎缩症)的等位基因是X连锁的。 据说因这些疾病而杂合的女性是携带者,可能不会表现出任何表型效应。 这些雌性会将这种疾病传染给一半的儿子,并将携带者身份传给一半的女儿;因此,隐性 X 连锁特征在男性中出现的频率要高于雌性。
在一些具有性染色体的生物体中,具有非同源性染色体的性别是雌性而不是雄性。 所有鸟类都是如此。 在这种情况下,与性别相关的特征更有可能出现在女性身上,因为它们是半精的。
人类性相关疾病
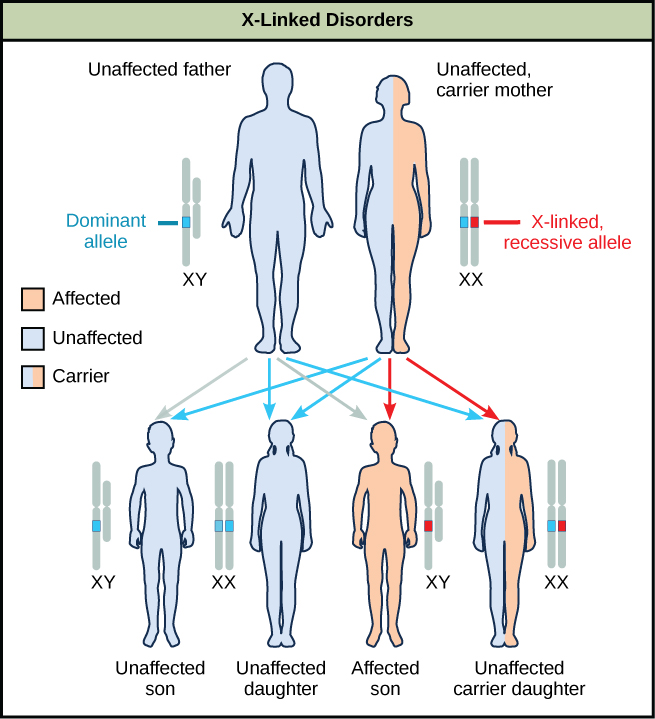
摩根实验室的性别连锁研究为了解人类X连锁隐性疾病(包括红绿色色盲以及A型和B型血友病)提供了基础知识。 由于人类男性只需要继承一个隐性突变体 X 等位基因即可受到影响,因此在男性中观察到 X 连锁疾病的比例不成比例。 雌性必须从父母双方那里继承隐性 X 连锁等位基因才能表达这种特征。 当他们继承一个隐性 X 连锁突变等位基因和一个显性的 X 连锁野生型等位基因时,它们是该特征的携带者,通常不受影响。 由于位于其中一条 X 染色体上的显性等位基因失活,携带者雌性可能表现出轻微的特征。 但是,女性携带者可以将该特征贡献给儿子,从而使儿子表现出该特征,或者她们可以将隐性等位基因贡献给女儿,从而使女儿成为该特征的携带者(图\(\PageIndex{10}\))。 尽管存在一些 Y 型连锁隐性疾病,但它们通常与男性的不育有关,因此不会传播给后代。
链接到学习
观看此视频,了解有关性别相关特征的更多信息。
杀伤力
个体基因组中的很大一部分基因对生存至关重要。 有时,必需基因的非功能性等位基因可以通过突变产生并在人群中传播,只要具有该等位基因的个体也有野生型的功能拷贝。 野生型等位基因的功能足以维持生命,因此被认为比非功能性等位基因占主导地位。 但是,请考虑两个杂合父母,他们的基因型为野生型/非功能突变体,这是假设的必需基因。 在他们四分之一的后代中,我们预计会观察到对非功能等位基因具有纯合隐性遗传的个体。 由于基因是必不可少的,这些人可能无法在受精后发育、在子宫内死亡或在晚年死亡,具体取决于哪个生命阶段需要这个基因。 一种遗传模式,即等位基因仅以纯合形式致命,杂合子可能正常或非致命表型发生某种改变,这种遗传模式被称为隐性致命。
对于具有隐性致死等位基因的杂合个体之间的杂合个体之间的杂交,该等位基因在纯合时会导致出生前死亡,只能观察到野生型纯合子和杂合子。 因此,基因型比率将为 2:1。 在其他情况下,隐性致死等位基因也可能在杂合子中表现出显性(但不是致命的)表型。 例如,果蝇中的隐性致命 Curly 等位基因会影响杂合子形式的翅膀形状,但在纯合子中却是致命的。
野生型等位基因的单个拷贝并不总是足以维持正常功能甚至存活。 占主导地位的致死遗传模式是等位基因在纯合子和杂合子中均具有致命性;只有在致死表型发生在育龄之后,该等位基因才能传播。 具有导致显性致命等位基因的突变的个体即使是杂合子形式也无法存活。 显性致命等位基因非常罕见,因为正如你所预料的那样,该等位基因只能持续一代并且不会传播。 但是,正如隐性致命等位基因可能不会立即表现出死亡表型一样,显性致死等位基因也可能要等到成年后才能表达。 一旦个体达到生育年龄,等位基因可能会在不知不觉中传播,从而导致两代人延迟死亡。 人类中的一个例子是亨廷顿氏病,在这种病中,神经系统逐渐消失(图\(\PageIndex{11}\))。 对占主导地位的亨廷顿等位基因(Hh)具有杂合性的人不可避免地会患上这种致命疾病。 但是,亨廷顿舞蹈症的发作可能要到40岁才会发生,那时患者可能已经将等位基因传给了50%的后代。
摘要
当在某种特征上存在差异的真正繁殖或纯合个体交叉时,所有后代都将成为该特征的杂合子。 如果这些特征被遗传为显性和隐性特征,则 F 1 后代都将表现出与显性特征的纯合亲本相同的表型。 如果这些杂合子后代是自交叉的,则由此产生的 F 2 后代同样有可能继承具有显性或隐性特征的配子,从而产生四分之一是纯合显性的,一半是杂合遗传,四分之一是纯合隐性遗传。 由于纯合显性个体和杂合个体在表型上是相同的,因此 F 2 后代中观察到的特征将表现出三个显性与一个隐性遗传的比例。
等位基因并不总是以显性和隐性模式表现。 不完全的优势描述了杂合子表现出介于纯合表型之间的表型的情况。 Codominance 描述了杂合子中两个等位基因的同时表达。 尽管二倍体生物对于任何给定基因只能有两个等位基因,但一个群体中存在一个基因的两个以上的等位基因是很常见的。 在人类中,就像在许多动物和某些植物中一样,雌性有两条X染色体,雄性有一条X和一条Y染色体。 据说存在于X染色体但不存在于Y染色体的基因是X连锁基因,因此雄性仅继承该基因的一个等位基因,而雌性继承两个等位基因。 最后,一些等位基因可能是致命的。 隐性致命等位基因仅在纯合子中具有致命性,但显性致命等位基因在杂合子中也是致命的。
脚注
- 1 Sumiti Vinayak 等人,“耐磺胺多辛恶性疟原虫的起源和演变”,公共科学图书馆病原体 6,第 3 期(2010):e1000830,doi: 10.1371/journal.ppat.1000830。
词汇表
- 等位基因
- 由突变产生并存在于同源染色体上相同相对位置的基因变异
- 常染色体
- 任何非性染色体
- 共同支配地位
- 在杂合子中,两个等位基因具有相同特征的完整同时表达
- 显性致命性
- 遗传模式,其中等位基因在纯合子和杂合子中都是致命的;只有在致死表型发生在育龄之后,该等位基因才能传播
- 基因型
- 生物体的潜在基因构成,包括物理上可见和未表达的等位基因
- hemizygous
- 一个特征只存在一个等位基因,比如在 X 连锁中;hemizygosity 使得对优势和隐性的描述变得无关紧要
- 杂合的
- 同源染色体上给定基因有两个不同的等位基因
- 纯合的
- 同源染色体上给定基因有两个相同的等位基因
- 统治不完整
- 在杂合子中,表达两个对比鲜明的等位基因,使个体表现出中间表型
- 单杂种
- 两个真正繁殖的父母之间交叉的结果,他们只对一个特征表现出不同的特征
- 表型
- 生物体表达的可观察特征
- Punnett 广场
- 两个个体之间交叉的视觉表现,其中每个个体的配子分别沿着网格的顶部和侧面表示,可能的合子基因型在网格的每个方框中重组
- 隐性致命
- 遗传模式,其中等位基因仅以纯合形式致命;杂合子可能正常或具有某些改变的非致命表型
- 与性有关的
- 性染色体上的任何基因
- 测试十字架
- 具有未知基因型的显性表达个体与纯合隐性遗传个体之间的交叉;后代表型表明未知亲本是显性特征的杂合子还是纯合子
- X 联动
- 基因存在于 X,但 Y 染色体上不存在