
9.3:对信号的响应
- Page ID
- 202775

培养技能
- 描述信号通路如何引导蛋白表达、细胞代谢和细胞生长
- 识别 PKC 在信号转导通路中的功能
- 认识细胞凋亡在健康生物体发育和维持中的作用
在细胞内部,配体与其内部受体结合,使它们能够直接影响细胞的DNA和蛋白质产生机制。 使用信号转导途径,质膜中的受体会对细胞产生各种影响。 信号通路的结果千差万别,取决于所涉及的细胞类型以及外部和内部条件。 下面介绍了一小部分答复样本。
基因表达
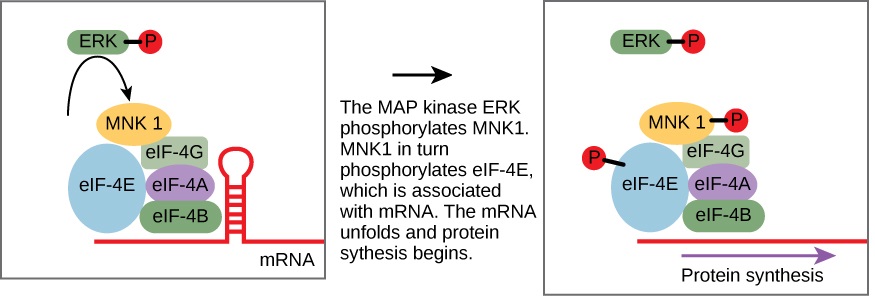
一些信号转导途径调节 RNA 的转录。 其他人则调节来自mRNA的蛋白质的翻译。 调节细胞核翻译的蛋白质的一个例子是 MAP 激酶 ERK。 当表皮生长因子(EGF)结合表皮生长因子受体时,ERK在磷酸化级联中被激活(见图9.2.1)。 磷酸化后,ERK 进入细胞核并激活蛋白激酶,该蛋白激酶反过来又调节蛋白质翻译(图\(\PageIndex{1}\))。
PKC 可以与之相互作用的第二种蛋白质是充当抑制剂的蛋白质。 抑制剂是一种与蛋白质结合并阻止其发挥作用或降低其功能的分子。 在这种情况下,抑制剂是一种叫做 Iμ-B 的蛋白质,它与调节蛋白 NF-μB 结合。 (符号代表希腊字母 kappa。) 当 Iμ-B 与 NF-μB 结合时,该复合物无法进入细胞核,但是当 PKC 磷酸化时,它就无法再结合 NF-μB,而 NF-μB(一种转录因子)可以进入细胞核并启动 RNA 转录。 在这种情况下,磷酸化的作用是使抑制剂失活,从而激活转录过程。
增加细胞新陈代谢
另一种信号通路的结果会影响肌肉细胞。 肾上腺素激活肌肉细胞中的β-肾上腺素能受体会导致细胞内环AMP(cAMP)的增加。 肾上腺素也被称为肾上腺素,是一种激素(由附着在肾脏上的肾上腺产生),可使人体为短期紧急情况做好准备。 Cyclic AMP 激活 PKA(蛋白激酶 A),后者反过来又磷酸化两种酶。 第一种酶通过激活中间糖原磷酸化酶激酶(GPK)来促进糖原降解,后者反过来激活糖原磷酸化酶(GP),将糖原分解为葡萄糖。 (回想一下,你的身体会将多余的葡萄糖转化为糖原以便短期储存。 当需要能量时,糖原会迅速重新转化为葡萄糖。) 第二种酶糖原合酶(GS)的磷酸化抑制了其从葡萄糖中形成糖原的能力。 通过这种方式,肌肉细胞通过糖原降解激活葡萄糖的形成,抑制葡萄糖形成糖原的使用,从而防止糖原降解和合成的徒劳循环,从而获得现成的葡萄糖池。 然后,葡萄糖可供肌肉细胞使用,以应对肾上腺素的突然激增,即 “战斗或逃跑” 反射。
细胞生长
细胞信号通路在细胞分裂中也起着重要作用。 除非受到来自其他细胞的信号的刺激,否则细胞通常不会分裂。 促进细胞生长的配体被称为生长因子。 大多数生长因子与与酪氨酸激酶相关的细胞表面受体结合。 这些细胞表面受体被称为受体酪氨酸激酶(RTK)。 激活 RTK 会启动信号通路,该信号通路包括一种叫做 RAS 的 G 蛋白,它会激活前面描述的 MAP 激酶通路。 然后,酶 MAP 激酶刺激与其他细胞成分相互作用的蛋白质的表达,从而启动细胞分裂。
职业联系:癌症生物学家
癌症生物学家研究癌症的分子起源,目的是开发新的预防方法和治疗策略,在不损害人体正常细胞的情况下抑制肿瘤的生长。 如前所述,信号通路控制细胞生长。 这些信号通路由信号蛋白控制,而信号蛋白反过来又由基因表达。 这些基因的突变可能导致信号蛋白失效。 这会阻止细胞调节其细胞周期,从而触发不受限制的细胞分裂和癌症。 调节信号蛋白的基因是一种癌基因,它是一种有可能导致癌症的基因。 编码 RAS 的基因是一种癌基因,最初是在 RAS 蛋白突变与癌症有关时发现的。 进一步的研究表明,30%的癌细胞在RAS基因中有突变,导致生长失控。 如果不加以控制,不受控制的细胞分裂可能导致肿瘤的形成和转移,即癌细胞在体内新部位的生长。
癌症生物学家已经能够识别出许多其他导致癌症发展的癌基因。 例如,HER2 是一种细胞表面受体,在 20% 的人类乳腺癌中过量存在。 癌症生物学家意识到基因复制导致25%的乳腺癌患者HER2过度表达,因此开发了一种名为赫赛汀(曲妥珠单抗)的药物。 赫赛汀是一种单克隆抗体,靶向 HER2,由免疫系统清除。 赫赛汀疗法有助于通过HER2控制信号传导。 赫赛汀与化疗联合使用有助于提高转移性乳腺癌患者的总体存活率。
细胞死亡
当细胞受损、多余或对生物体具有潜在危险时,细胞可以启动触发程序性细胞死亡或凋亡的机制。 细胞凋亡允许细胞以受控的方式死亡,从而防止潜在的有害分子从细胞内部释放。 有许多内部检查点可以监测细胞的健康状况;如果观察到异常,细胞可以自发启动细胞凋亡过程。 但是,在某些情况下,例如病毒感染或癌症导致的细胞分裂失控,细胞的正常制衡会失败。 外部信号也可以引发细胞凋亡。 例如,大多数正常动物细胞都有与细胞外基质相互作用的受体,细胞外基质是一种糖蛋白网络,为生物体中的细胞提供结构支持。 细胞受体与细胞外基质的结合在细胞内启动信号级联。 但是,如果细胞远离细胞外基质,则信号传导停止,细胞会发生凋亡。 该系统可防止细胞在人体中传播和增殖失控,就像转移的肿瘤细胞一样。
导致细胞凋亡的外部信号传导的另一个例子发生在 T 细胞发育中。 T 细胞是与外来大分子和颗粒结合的免疫细胞,并将其靶向以免疫系统破坏。 通常,T细胞不靶向 “自身” 蛋白(自身生物体的蛋白),这种过程可能导致自身免疫性疾病。 为了培养区分自身和非自我的能力,对未成熟的T细胞进行筛查,以确定它们是否与所谓的自身蛋白结合。 如果 T 细胞受体与自身蛋白质结合,则该细胞会启动细胞凋亡以移除潜在的危险细胞。
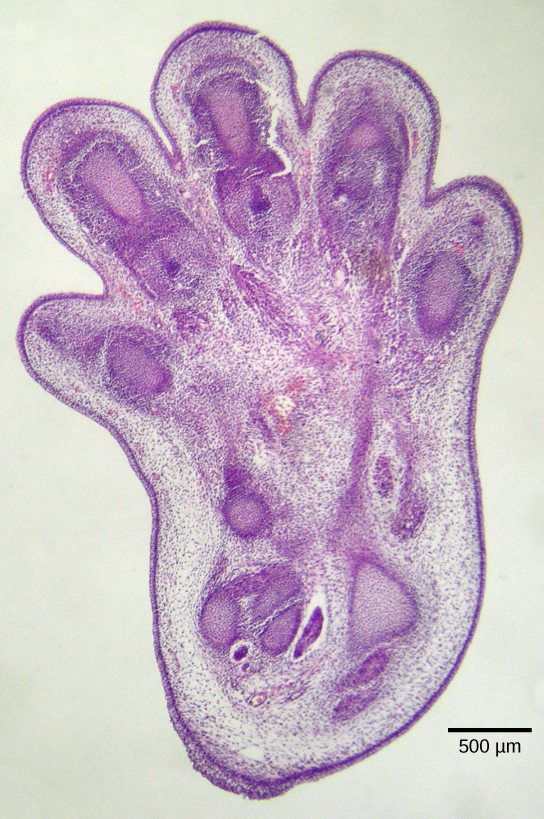
细胞凋亡对于正常的胚胎发育也至关重要。 例如,在脊椎动物中,发育的早期阶段包括在单个手指和脚趾之间形成网状组织(图\(\PageIndex{2}\))。 在正常发育过程中,必须消除这些不需要的细胞,从而形成完全分离的手指和脚趾。 细胞信号传导机制会触发细胞凋亡,细胞凋亡会破坏发育数字之间的细胞。
信号级联的终止
肿瘤细胞中经常出现的异常信号传导证明,在适当的时间终止信号与信号的启动同样重要。 停止特定信号的一种方法是降解配体或将其移除,使其无法再访问其受体。 雌激素和睾丸激素等疏水性激素引发长期事件的原因之一是因为它们结合了载体蛋白。 这些蛋白质使不溶性分子溶于血液,但它们也可以保护激素免受循环酶的降解。
在细胞内部,许多不同的酶逆转了信号级联产生的细胞修饰。 例如,磷酸酶是在称为脱磷酸化的过程中通过激酶去除附着在蛋白质上的磷酸基团的酶。 环状AMP(cAMP)被磷酸二酯酶降解为AMP,而位于细胞外膜和内膜中的Ca 2+ 泵会逆转钙储存的释放。
摘要
信号通路的启动是对外部刺激的反应。 这种反应可以采取许多不同的形式,包括蛋白质合成、细胞新陈代谢的变化、细胞生长甚至细胞死亡。 许多途径通过启动基因表达来影响细胞,所使用的方法相当多。 一些途径激活与 DNA 转录因子相互作用的酶。 其他人则修改蛋白质并诱导它们改变其在细胞中的位置。 根据生物体的状态,细胞可以通过以糖原或脂肪的形式储存能量,或者以葡萄糖的形式提供能量来做出反应。 信号转导途径允许肌肉细胞以葡萄糖的形式对即时能量需求做出反应。 细胞生长几乎总是受到称为生长因子的外部信号的刺激。 不受控制的细胞生长会导致癌症,编码信号通路蛋白质成分的基因突变通常存在于肿瘤细胞中。 程序性细胞死亡或细胞凋亡对于去除受损或不必要的细胞很重要。 使用细胞信号来组织细胞的拆解,可确保细胞质中的有害分子不会像在不受控制的死亡和坏死中那样被释放到细胞之间的空间中。 细胞凋亡还确保了死细胞成分的有效回收。 终止蜂窝信号级联非常重要,这样对信号的响应在时间和强度上都是适当的。 信号分子的降解和磷酸酶对通路中磷酸化中间体的脱磷酸化是终止细胞内信号的两种方法。
词汇表
- 细胞凋亡
- 程序性细胞死亡
- 生长因子
- 与细胞表面受体结合并刺激细胞生长的配体
- 抑制剂
- 与蛋白质(通常是酶)结合并使其无法发挥作用的分子
- 磷酸酶
- 从先前经过磷酸化的分子中去除磷酸基团的酶
- 磷酸二酯酶
- 降解 cAMP、产生 AMP 以终止信号传导的酶