
9.2: 信号的传播
- Page ID
- 202761

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 解释配体的结合如何启动整个细胞的信号转导
- 认识磷酸化在细胞内信号传输中的作用
- 评估第二信使在信号传输中的作用
一旦配体与受体结合,信号就会通过膜传递到细胞质中。 以这种方式延续信号称为信号转导。 信号转导仅发生在细胞表面受体上,因为内部受体能够直接与细胞核中的DNA相互作用以启动蛋白质合成。
当配体与其受体结合时,会发生构象变化,影响受体的细胞内域。 配体结合后细胞外结构域的构象变化可以在受体的膜区域传播,并导致细胞内结构域或其相关蛋白的激活。 在某些情况下,配体的结合会导致受体的二聚化,这意味着两个受体相互结合形成一种称为二聚体的稳定复合物。 二聚体是两个分子(通常是相同的)结合在一起时形成的化合物。 受体以这种方式的结合使它们的细胞内结构域能够密切接触并相互激活。
结合启动信号通路
配体与细胞表面受体结合后,受体细胞内成分的激活会引发一系列事件,称为信号通路或信号级联。 在信号通路中,第二信使、酶和活化蛋白与特定蛋白质相互作用,而特定的蛋白质反过来又在连锁反应中被激活,最终导致细胞环境的变化(图\(\PageIndex{1}\))。 瀑布中的事件连续发生,就像河流中的水流一样。 在某个点之前发生的交互被定义为上游事件,该点之后的事件称为下游事件。
艺术连接
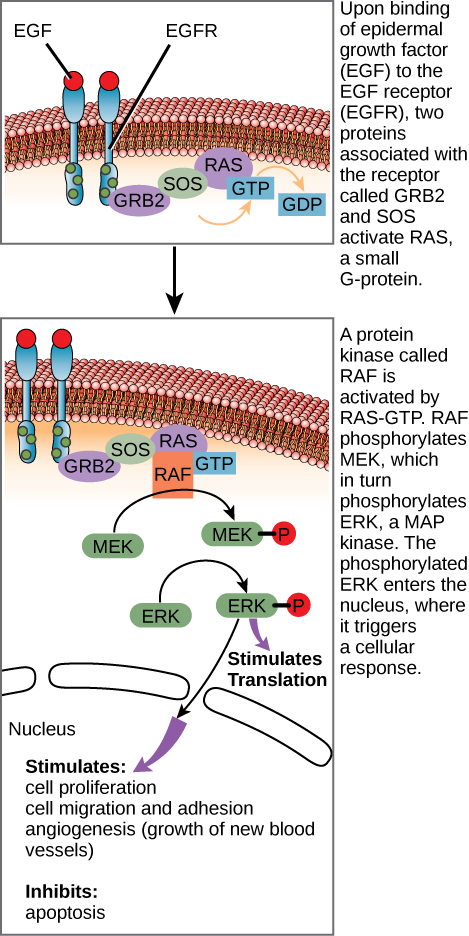
在某些癌症中,RAS G 蛋白的 GTPase 活性受到抑制。 这意味着 RAS 蛋白无法再将 GTP 水解成 GDP。 这会对下游细胞事件产生什么影响?
信号通路很快就会变得非常复杂,因为大多数细胞蛋白会影响不同的下游事件,具体取决于细胞内部的条件。 根据两个或多个信号通路之间的相互作用,单个通路可以向不同的端点分支,而相同的配体通常用于在不同的细胞类型中启动不同的信号。 这种反应变化是由于不同细胞类型中蛋白质表达的差异造成的。 另一个复杂的因素是通路的信号整合,其中来自两个或多个不同细胞表面受体的信号合并以激活细胞中的相同反应。 此过程可以确保在单元提交特定响应之前满足多个外部要求。
细胞外信号的影响也可以通过酶级联放大。 在信号启动时,单个配体与单个受体结合。 但是,激活受体连接酶可以激活信号级联成分的许多拷贝,从而放大信号。
细胞内信号传导方法
信号通路的诱导取决于酶对细胞成分的修饰。 可能发生许多酶促修饰,下游的下一个成分反过来识别它们。 以下是细胞内信号传导中一些比较常见的事件。
磷酸化
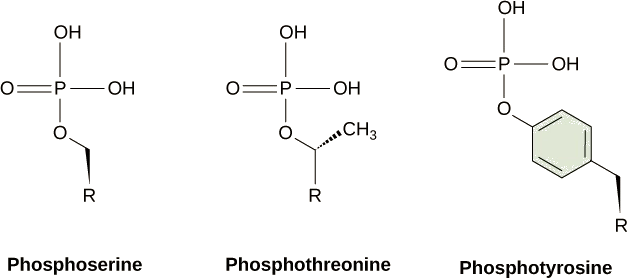
信号通路中最常见的化学修饰之一是在称为磷酸化的过程中向蛋白质等分子中添加磷酸基团(PO 4 —3)。 磷酸盐可以添加到诸如GMP之类的核苷酸中以形成GDP或GTP。 磷酸盐还经常被添加到蛋白质的丝氨酸、苏氨酸和酪氨酸残基中,它们取代氨基酸的羟基(图\(\PageIndex{2}\))。 磷酸盐的转移是由一种叫做激酶的酶催化的。 各种激酶以其磷酸化的底物命名。 丝氨酸和苏氨酸残基的磷酸化通常会激活酶。 酪氨酸残基的磷酸化既可以影响酶的活性,也可以产生与信号级联中下游成分相互作用的结合位点。 磷酸化可能会激活或失活酶,而磷酸化的逆转,即磷酸酶的脱磷酸化,将逆转这种影响。
第二信使
第二个信使是小分子,在信号分子与受体的结合引发信号后,它们会传播信号。 这些分子通过改变某些细胞蛋白的行为来帮助通过细胞质传播信号。
钙离子是一种广泛使用的第二信使。 细胞内钙离子(Ca 2+)的游离浓度非常低,因为质膜中的离子泵持续使用腺苷-5'-三磷酸(ATP)将其去除。 出于信号传导目的,Ca 2+ 储存在细胞质囊泡中,例如内质网,或者从细胞外部获取。 当信号传导发生时,配体门控的钙离子通道允许存在于细胞外(或细胞内储存室中)的较高水平的 Ca 2+ 流入细胞质,从而提高细胞质 Ca 2+ 的浓度。 对钙 2+ 增加的反应因所涉及的细胞类型而异。 例如,在胰腺的β细胞中,Ca 2+ 信号传导导致胰岛素的释放,而在肌肉细胞中,Ca 2+ 的增加会导致肌肉收缩。
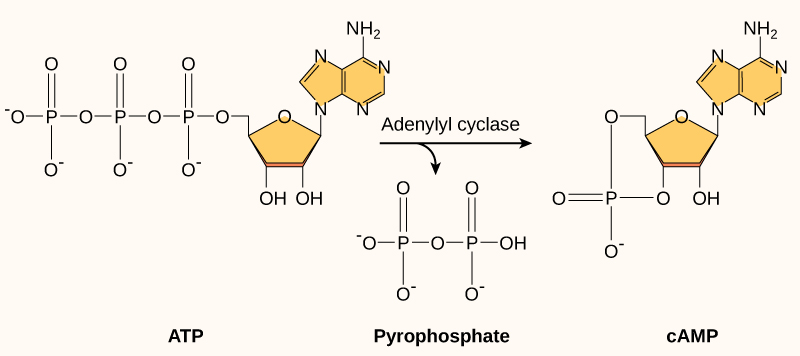
许多不同细胞类型中使用的另一个第二信使是循环AMP(cAMP)。 Cyclic AMP 由来自 ATP 的腺苷环化酶合成(图\(\PageIndex{3}\))。 cAMP 在细胞中的主要作用是结合并激活一种叫做 CAMP 依赖性激酶(a-激酶)的酶。 A-激酶调节许多重要的代谢途径:它磷酸化其靶蛋白的丝氨酸和苏氨酸残基,并在此过程中激活它们。 A-激酶存在于许多不同类型的细胞中,每种细胞中的靶蛋白也不同。 差异会导致不同细胞对 cAMP 的反应发生变化。
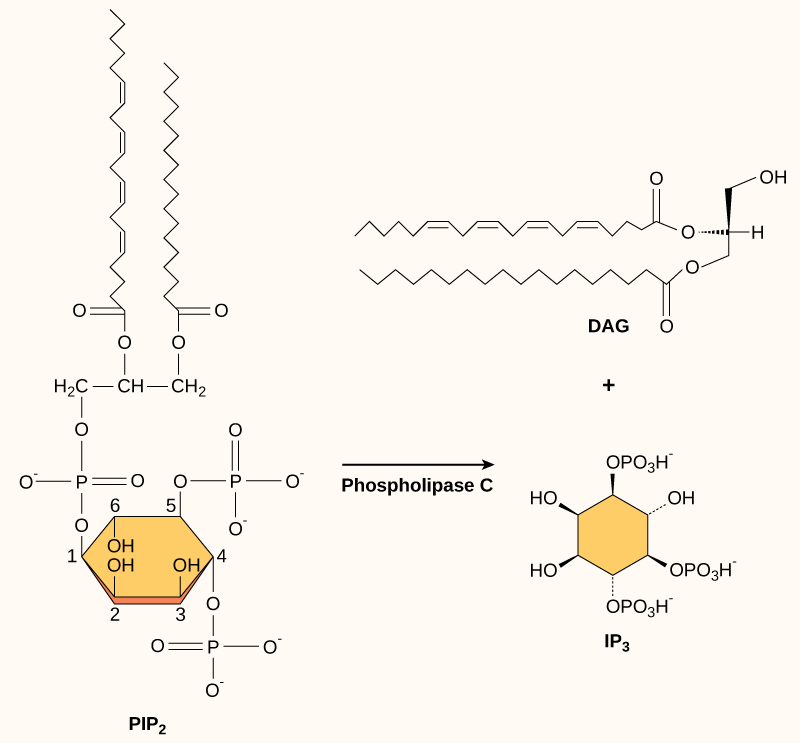
肌醇磷脂以低浓度存在于质膜中,是脂质,也可以转化为第二信使。 因为这些分子是膜成分,所以它们位于膜结合受体附近,很容易与它们相互作用。 磷脂酰肌醇(PI)是在细胞信号传导中起作用的主要磷脂。 被称为激酶的酶磷酸化 PI 以形成 PI-磷酸 (PIP) 和 PI-二磷酸 (PIP 2)。
磷脂酶 C 分解 PIP 2 形成二酰基甘油 (DAG) 和三磷酸肌醇 (IP 3)(图\(\PageIndex{4}\))。 PIP 2 分裂的这些产物充当了第二个信使。 二酰基甘油 (DAG) 残留在质膜中并激活蛋白激酶 C (PKC),蛋白激酶 C (PKC) 随后磷酸化其靶蛋白中的丝氨酸和苏氨酸残基。 IP 3 扩散到细胞质中并与内质网中的配体门控钙通道结合,释放出延续信号级联的 Ca 2+。
摘要
与受体结合的配体允许通过细胞进行信号转导。 通过细胞传递信号的事件链称为信号通路或级联。 由于不同蛋白质之间的相互作用,信号通路通常非常复杂。 细胞信号级联的一个主要组成部分是被称为激酶的酶对分子进行磷酸化。 磷酸化在蛋白质中的丝氨酸、苏氨酸和酪氨酸残基中添加磷酸基团,改变其形状,激活或失活蛋白质。 像核苷酸这样的小分子也可以被磷酸化。 第二个信使是小型的非蛋白质分子,用于在细胞内传输信号。 第二个信使的一些例子是钙离子(Ca 2+)、环状AMP(cAMP)、二酰基甘油(DAG)和三磷酸肌醇(IP 3)。
艺术联系
图\(\PageIndex{1}\):在某些癌症中,RAS G 蛋白的 GTPase 活性受到抑制。 这意味着 RAS 蛋白无法再将 GTP 水解成 GDP。 这会对下游细胞事件产生什么影响?
- 回答
-
ERK 会被永久激活,导致细胞增殖、迁移、粘附和新血管的生长。 细胞凋亡会受到抑制。
词汇表
- 循环放大器 (cAMP)
- 源自 ATP 的第二个信使
- 循环 AMP 依赖性激酶
- (也称为蛋白激酶 A 或 PKA)通过与 cAMP 结合而激活的激酶
- 二酰基甘油 (DAG)
- 用于质膜内信号传导的 PIP 2 的分解产物
- 二聚体
- 两个分子结合在一起时形成的化合物
- 二聚化
- (指受体蛋白)两种受体蛋白相互作用形成一种叫做二聚体的功能复合物
- 肌醇磷脂
- 脂质以少量浓度存在于质膜中,然后转化为第二个信使;它以肌醇(一种碳水化合物)作为其亲水性头组
- 三磷酸肌醇 (IP 3)
- 用于细胞内信号传导的 PIP 2 的分解产物
- 激酶
- 催化磷酸基团从 ATP 转移到另一个分子的酶
- 第二个信使
- 小型的非蛋白质分子,在受体激活导致其释放后在细胞内传播信号
- 信号集成
- 来自两个或多个不同细胞表面受体的信号相互作用,这些受体合并以激活细胞中的相同反应
- 信号转导
- 信号通过细胞的细胞质(有时还有细胞核)传播
- 信号通路
- (也是信号级联)细胞质中发生的一系列事件,这些事件发生在细胞质中,目的是传播来自质膜的信号以产生响应