
9.1: 信号分子和细胞受体
- Page ID
- 202760

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)培养技能
- 描述在多细胞生物体中发现的四种信号传导类型
- 比较内部受体和细胞表面受体
- 认识配体的结构及其作用机制之间的关系
活细胞世界中有两种交流。 细胞之间的通信称为细胞间信号,细胞内的通信称为细胞内信号。 记住区别的一种简单方法是理解前缀的拉丁语起源:inter-表示 “之间”(例如,相交线是相互交叉的线),intra-表示 “内部”(如静脉注射)。
化学信号由信号传导细胞以称为配体的小、通常是挥发性或可溶性分子的形式释放。 配体是结合另一个特定分子的分子,在某些情况下,在此过程中传递信号。 因此,配体可以被视为信号分子。 配体与靶细胞中的蛋白质相互作用,靶细胞是受化学信号影响的细胞;这些蛋白质也被称为受体。 配体和受体分为多个变种;但是,特定的配体将具有通常仅结合该配体的特定受体。
信号形式
在多细胞生物中发现了四类化学信号:旁分泌信号传导、内分泌信号传导、自分泌信号传导和间隙连接处的直接信号(图\(\PageIndex{1}\))。 不同类别的信号传导之间的主要区别在于信号通过生物体到达目标细胞的距离。 并非所有细胞都受到相同信号的影响。
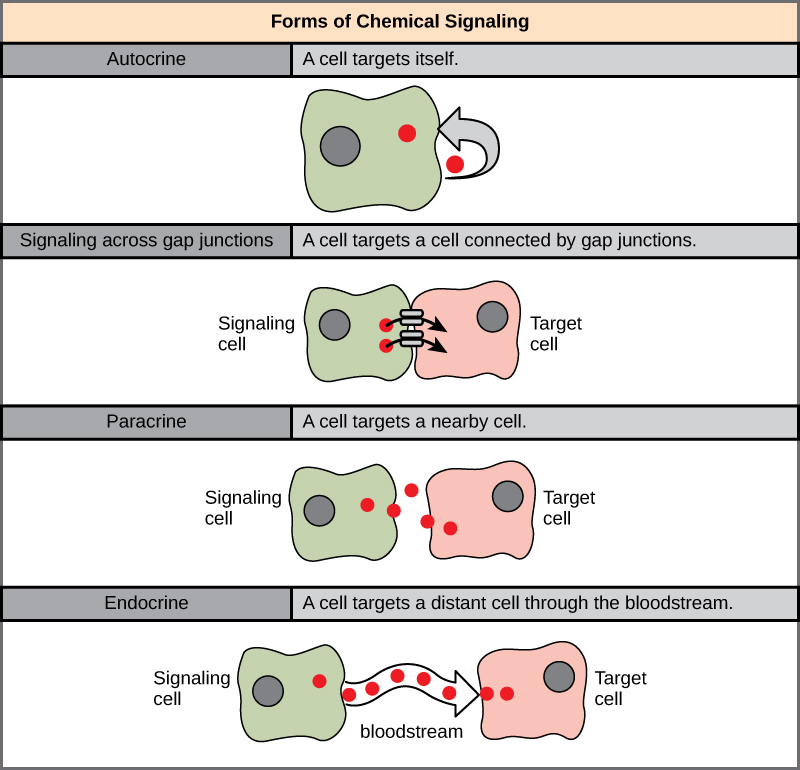
旁分泌信号传导
在靠近的细胞之间局部作用的信号称为旁分泌信号。 旁分泌信号通过扩散在细胞外基质中移动。 这些类型的信号通常会引起仅持续很短时间的快速响应。 为了保持反应局部化,旁分泌配体分子通常会被酶快速降解或被邻近细胞去除。 移除信号将重新建立信号的浓度梯度,使它们在再次释放时能够在细胞内空间快速扩散。
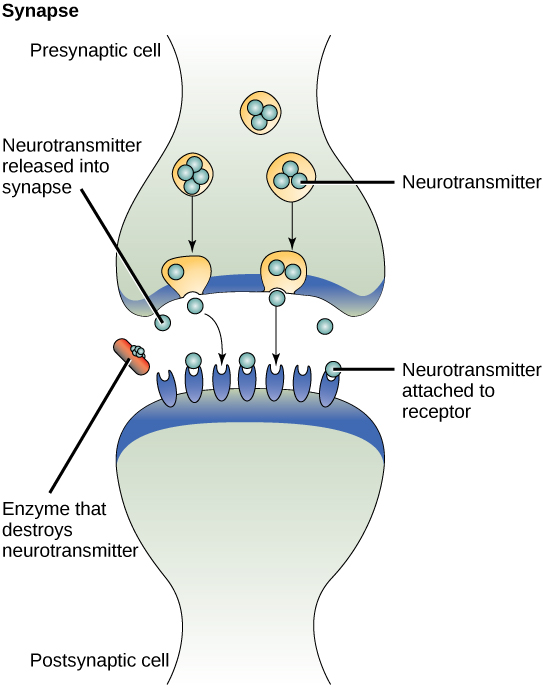
旁分泌信号传导的一个例子是信号通过神经细胞之间的突触传输。 神经细胞由细胞体、几个接受刺激的称为树突的短分支延伸部分和一个称为轴突的长延伸部分组成,后者将信号传输到其他神经细胞或肌肉细胞。 发生信号传输的神经细胞之间的交界处称为突触。 突触信号是在神经细胞之间传播的化学信号。 神经细胞内的信号通过快速移动的电脉冲传播。 当这些脉冲到达轴突末端时,通过突触前细胞(发出信号的细胞)释放称为神经递质的化学配体,信号会继续传递到下一个细胞的树突上。 神经递质在神经细胞之间的很短距离内传输,神经细胞被称为化学突触(图\(\PageIndex{2}\))。 神经细胞之间的小距离允许信号快速传播;这样可以立即做出反应,例如,把手从火炉上移开!
当神经递质结合突触后细胞表面的受体时,靶细胞的电化学电位发生变化,并启动下一个电脉冲。 释放到化学突触中的神经递质会迅速降解或被突触前细胞重新吸收,因此受体神经细胞可以快速恢复,并做好对下一个突触信号快速反应的准备。
内分泌信号
来自远处细胞的信号被称为内分泌信号,它们来自内分泌细胞。 (在体内,许多内分泌细胞位于内分泌腺体中,例如甲状腺、下丘脑和垂体。) 这些类型的信号通常会产生较慢的响应,但效果更持久。 内分泌信号中释放的配体被称为激素,这些信号分子在身体的一部分产生,但会影响一段距离之外的其他身体部位。
激素通过血液在内分泌细胞和靶细胞之间传播很远的距离,这是一种在全身移动的相对较慢的方式。 由于其运输形式,激素作用于靶细胞时会被稀释并以低浓度存在。 这与旁分泌信号传导不同,在旁分泌信号传导中,配体的局部浓度可能非常高。
自分泌信号
自分泌信号由信号传导细胞产生,这些信号细胞也可以与释放的配体结合。 这意味着信号单元和目标小区可以是相同或相似的小区(前缀 a uto- 表示自我,提醒信号单元向自身发送信号)。 这种类型的信号传导通常发生在生物体的早期发育期间,以确保细胞发育成正确的组织并发挥适当的功能。 自分泌信号还调节疼痛感和炎症反应。 此外,如果细胞感染了病毒,则该细胞可以发出自身信号,表明其会经历程序性细胞死亡,在此过程中杀死病毒。 在某些情况下,相同类型的相邻细胞也会受到释放配体的影响。 在胚胎发育中,这种刺激一组相邻细胞的过程可能有助于引导相同的细胞分化为相同的细胞类型,从而确保正确的发育结果。
跨间隙连接处的直接信令
动物体内的间隙连接和植物中的 plasmodesmata 是相邻细胞的质膜之间的连接。 这些充满水的通道允许称为细胞内介质的小信号分子在两个细胞之间扩散。 小分子,例如钙离子(Ca 2+),能够在细胞之间移动,但蛋白质和DNA等大分子无法通过通道。 信道的特异性确保细胞保持独立,但可以快速轻松地传输信号。 信号分子的转移传递直接与靶细胞相邻的细胞的当前状态;这允许一组细胞协调它们对其中只有一个可能接收到的信号的反应。 在植物中,plasmodesmata 无处不在,使整个工厂变成了一个巨大的通信网络。
受体的类型
受体是靶细胞中或其表面结合配体的蛋白质分子。 有两种类型的受体,即内部受体和细胞表面受体。
内部受体
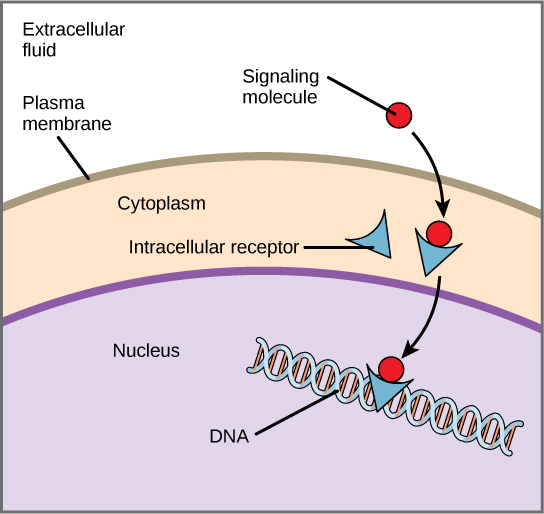
内部受体,也称为细胞内受体或细胞质受体,存在于细胞质中,对能够穿过质膜的疏水配体分子产生反应。 一旦进入细胞,这些分子中有许多会与作为 mRNA 合成(转录)调节剂的蛋白质结合,从而介导基因表达。 基因表达是将细胞 DNA 中的信息转化为氨基酸序列的细胞过程,氨基酸最终形成蛋白质。 当配体与内部受体结合时,会触发构象变化,暴露蛋白质上的 DNA 结合位点。 配体受体复合物进入细胞核,然后与染色体DNA的特定调节区域结合,促进转录的启动(图\(\PageIndex{3}\))。 转录是将细胞 DNA 中的信息复制成一种称为信使 RNA(mRNA)的特殊形式的 RNA 的过程;该细胞使用 mRNA(移出细胞质并与核糖体结合)中的信息以正确的顺序连接特定的氨基酸,从而产生蛋白质。 内部受体可以直接影响基因表达,无需将信号传递给其他受体或信使。
细胞表面受体
细胞表面受体,也称为跨膜受体,是与外部配体分子结合的细胞表面、膜锚定(整体)蛋白。 这种类型的受体跨越质膜并进行信号转导,其中细胞外信号被转换为细胞间信号。 与细胞表面受体相互作用的配体不必进入它们所影响的细胞。 细胞表面受体也被称为细胞特异性蛋白或标志物,因为它们对单个细胞类型具有特异性。
由于细胞表面受体蛋白是正常细胞功能的基础,因此其中任何一种蛋白质的故障都可能产生严重后果也就不足为奇了。 事实证明,某些受体分子的蛋白质结构错误在高血压(高血压)、哮喘、心脏病和癌症中起作用。
每个细胞表面受体都有三个主要成分:外部配体结合结构域、疏水膜跨越区和细胞内的细胞内结构域。 配体结合结构域也称为细胞外结构域。 每个结构域的大小和范围差异很大,具体取决于受体的类型。
进化连接:病毒如何识别主机
与活细胞不同,许多病毒没有质膜或维持生命所必需的任何结构。 有些病毒只是由含有 DNA 或 RNA 的惰性蛋白壳组成的。 要繁殖,病毒必须入侵作为宿主的活细胞,然后接管宿主的蜂窝设备。 但是病毒如何识别其宿主呢?
病毒通常与宿主细胞上的细胞表面受体结合。 例如,引起人类流感(流感)的病毒与呼吸系统细胞膜上的受体特异性结合。 宿主之间细胞表面受体的化学差异意味着感染特定物种(例如人类)的病毒无法感染其他物种(例如鸡)。
但是,与人类相比,病毒的DNA或RNA量非常少,因此,病毒可以迅速繁殖。 病毒繁殖总是会产生错误,从而导致新产生的病毒发生变化;这些变化意味着与细胞表面受体相互作用的病毒蛋白可能以可以与新宿主中的受体结合的方式进化。 这种变化是随机发生的,经常发生在病毒的繁殖周期中,但只有在具有新结合特性的病毒与合适的宿主接触时,这些变化才有意义。 就流感而言,这种情况可能发生在动物和人密切接触的环境中,例如家禽和养猪场。 1 一旦病毒跳转到新宿主上,它就会迅速传播。 科学家们密切关注新出现的病毒(称为新兴病毒),希望这种监测能够减少全球病毒流行的可能性。
细胞表面受体参与了多细胞生物的大部分信号传导。 细胞表面受体一般分为三类:离子通道连接受体、G 蛋白连接受体和酶联受体。
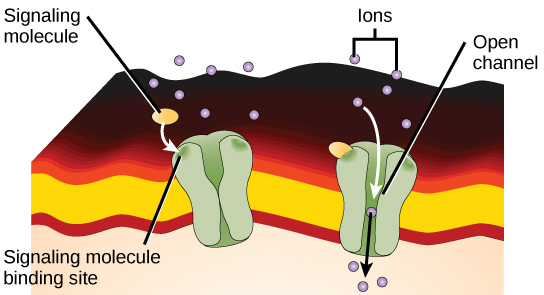
离子通道连接受体结合配体并打开穿过膜的通道,允许特定的离子通过。 为了形成通道,这种类型的细胞表面受体具有广泛的膜跨越区域。 为了与构成质膜中心的磷脂脂脂肪酸尾部相互作用,膜跨区域中的许多氨基酸本质上是疏水性的。 相反,排列在通道内部的氨基酸具有亲水性,允许水或离子通过。 当配体与通道的细胞外区域结合时,蛋白质结构会发生构象变化,允许钠、钙、镁和氢等离子通过(图\(\PageIndex{4}\))。
G 蛋白连接受体结合配体并激活一种称为 G 蛋白的膜蛋白。 然后,活化的 G 蛋白与离子通道或膜中的酶相互作用(图\(\PageIndex{5}\))。 所有 G 蛋白连接受体都有七个跨膜结构域,但每种受体都有自己特定的细胞外结构域和 G 蛋白结合位点。
使用 G 蛋白连接受体的细胞信号传导是一系列循环事件。 在配体结合之前,非活性 G 蛋白可以与受体上新发现的结合特异位点结合。 一旦 G 蛋白与受体结合,由此产生的形状变化就会激活 G 蛋白,从而释放 GDP 并提高 GTP。 然后,G 蛋白的亚基分裂为 α 亚单位和 βγ 亚单位。 因此,这些 G 蛋白片段中的一个或两个片段可能能够激活其他蛋白质。 过了一会儿,G 蛋白的活性 α 亚单位上的 GTP 被水解为 GDP,βγ 亚单位被失活。 亚单位重新结合,形成非活性 G 蛋白,循环重新开始。
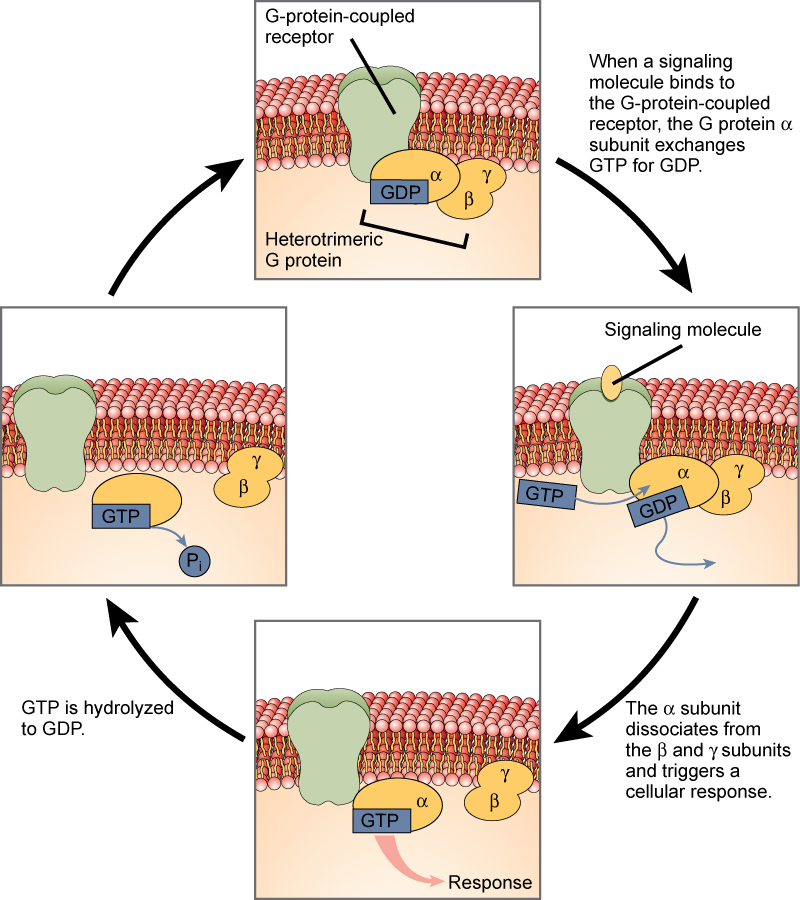
G蛋白连接受体已被广泛研究,关于它们在维持健康中的作用已经学到了很多。 对人类致病的细菌会释放毒物,从而中断特定的G蛋白连接受体功能,从而导致百日咳、肉毒中毒和霍乱等疾病。 例如,在霍乱(图\(\PageIndex{6}\))中,水传播的细菌霍乱弧菌会产生一种毒素——霍乱激素,它与小肠内的细胞结合。 然后,毒素进入这些肠道细胞,在那里它会修饰控制氯化物通道开放的 G 蛋白,使其保持持续活跃,从而导致体内大量液体流失,从而可能致命的脱水。

酶联受体是具有与酶相关的细胞内结构域的细胞表面受体。 在某些情况下,受体本身的细胞内结构域是一种酶。 其他酶联受体具有一个小的细胞内结构域,可直接与酶相互作用。 酶联受体通常具有较大的细胞外和细胞内结构域,但膜跨区由肽链的单个α-螺旋区组成。 当配体与细胞外结构域结合时,信号通过膜传输,激活酶。 该酶的激活在细胞内引发了一系列事件,最终导致反应。 这种酶联受体的一个例子是酪氨酸激酶受体(图\(\PageIndex{7}\))。 激酶是一种将磷酸基团从 ATP 转移到另一种蛋白质的酶。 酪氨酸激酶受体将磷酸基团转移到酪氨酸分子(酪氨酸残基)。 首先,信号分子与附近两个酪氨酸激酶受体的细胞外结构域结合。 然后,两个相邻的受体结合在一起或二聚化。 然后将磷酸盐添加到受体细胞内结构域的酪氨酸残基中(磷酸化)。 然后,磷酸化残留物可以将信号传输到细胞质中的下一个信使。
艺术连接
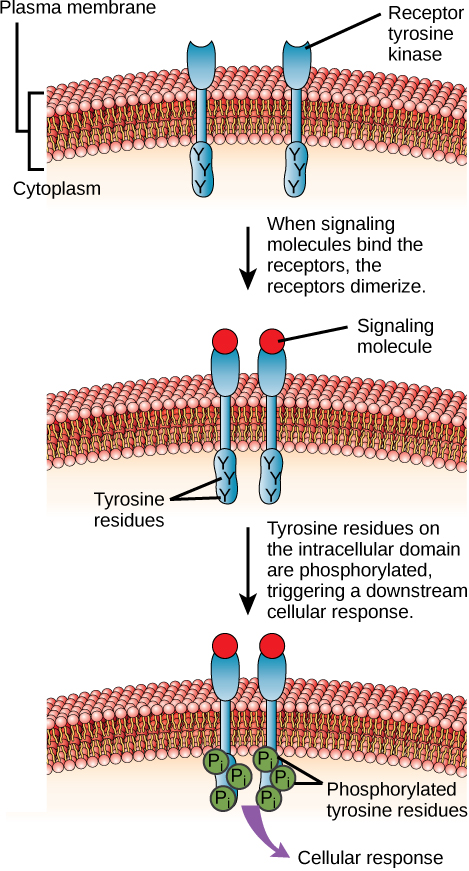
HER2 是一种受体酪氨酸激酶。 在30%的人类乳腺癌中,HER2被永久激活,导致细胞分裂不受控制。 拉帕替尼是一种用于治疗乳腺癌的药物,它抑制HER2受体酪氨酸激酶自动磷酸化(受体向自身添加磷酸盐的过程),从而使肿瘤生长减少50%。 除了自磷酸化外,拉帕替尼还会抑制以下哪个步骤?
- 信号传导分子结合、二聚化和下游细胞反应
- 二聚化和下游细胞反应
- 下游细胞反应
- 磷酸酶活性、二聚化和下游细胞反应
信号分子
配体由信号传导细胞产生,随后与靶细胞中的受体结合,充当化学信号,传播到靶细胞以协调反应。 作为配体的分子类型千差万别,从小蛋白质到钙(Ca 2+)等小离子不等。
小型疏水配体
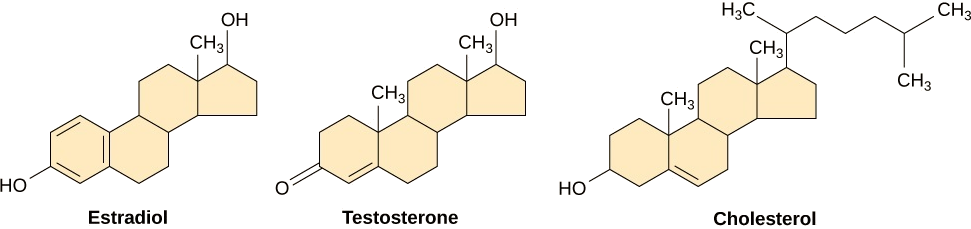
小型疏水配体可以直接通过质膜扩散并与内部受体相互作用。 这类配体的重要成员是类固醇激素。 类固醇是具有碳氢化合物骨架和四个熔合环的脂质;不同的类固醇具有附着在碳骨架上的不同官能团。 类固醇激素包括雌性激素雌二醇,这是一种雌激素;雄性激素,睾丸激素;以及胆固醇,胆固醇是生物膜的重要结构成分和类固醇激素的前体(图\(\PageIndex{8}\))。 其他疏水性激素包括甲状腺激素和维生素 D。为了溶于血液,疏水配体在血液中运输时必须与载体蛋白结合。
水溶性配体
水溶性配体是极性的,因此无法单独穿过质膜;有时,它们太大而根本无法穿过质膜。 相反,大多数水溶性配体与细胞表面受体的细胞外结构域结合。 这组配体非常多样化,包括小分子、肽和蛋白质。
其他配体
一氧化氮(NO)是一种同时充当配体的气体。 它能够直接在质膜上扩散,其作用之一是与平滑肌中的受体相互作用并诱导组织松弛。 NO 的半衰期很短,因此只能在很短的距离内起作用。 硝酸甘油是一种治疗心脏病的方法,其作用是触发一氧化氮的释放,这会导致血管扩张(扩张),从而恢复流向心脏的血液。 NO 最近已广为人知,因为它所影响的途径是治疗勃起功能障碍的处方药的靶向,例如伟哥(勃起涉及血管扩张)。
摘要
细胞通过细胞间和细胞内信号传导进行交流。 信号传导细胞分泌与靶细胞结合的配体,并在靶细胞内引发一系列事件。 多细胞生物中的四类信号传导是旁分泌信号传导、内分泌信号传导、自分泌信号和间隙连接处的直接信号。 旁分泌信号传导发生在短距离内。 内分泌信号由荷尔蒙在血液中长距离传输,自分泌信号由发送信号的同一个细胞或附近其他同类细胞接收。 间隙连接允许包括信号分子在内的小分子在相邻细胞之间流动。
内部受体存在于细胞质中。 在这里,它们结合穿过质膜的配体分子;这些受体-配体复合物移动到细胞核并直接与细胞 DNA 相互作用。 细胞表面受体将信号从细胞外部传递到细胞质。 离子通道连接的受体与它们的配体结合时,会在质膜中形成一个孔,某些离子可以穿过该孔隙。 G 蛋白连接受体与质膜细胞质侧的 G 蛋白相互作用,促进结合的 GDP 与 GTP 的交换,并与其他酶或离子通道相互作用以传递信号。 酶联受体将信号从细胞外部传递到膜结合酶的细胞内结构域。 配体结合导致酶的激活。 小型疏水配体(如类固醇)能够穿透质膜并与内部受体结合。 水溶性亲水配体无法通过膜;相反,它们与细胞表面受体结合,细胞表面受体将信号传递到细胞内部。
艺术联系
图\(\PageIndex{7}\):HER2 是一种受体酪氨酸激酶。 在30%的人类乳腺癌中,HER2被永久激活,导致细胞分裂不受控制。 拉帕替尼是一种用于治疗乳腺癌的药物,它抑制HER2受体酪氨酸激酶自动磷酸化(受体向自身添加磷酸盐的过程),从而使肿瘤生长减少50%。 除了自磷酸化外,拉帕替尼还会抑制以下哪个步骤?
- 信号传导分子结合、二聚化和下游细胞反应。
- 二聚化和下游细胞反应。
- 下游细胞反应。
- 磷酸酶活性、二聚化和下游细胞反应。
- 回答
-
C. 下游细胞反应将受到抑制。
脚注
- 1 A.B. Sigalov,《自然学院》。 IV。 向病毒学习,Self/Nonself 1,第 4 期(2010):282-298。 Y. Cao、X. Koh、L. Dong、X. Du、A. Wu、X. Ding、H. Deng、Y. Shu、J. Chen、T. Jiang,《流感病毒血凝素与人类和禽类受体的结合活性的快速估计》,PloS On e 6,第 4 期(2011):e18664。
词汇表
- 自分泌信号
- 由附近相同或相似的小区发送和接收的信号
- 细胞表面受体
- 细胞表面蛋白,即使配体没有进入细胞,也可以将信号从细胞外部传输到内部
- 化学突触
- 轴突末端和神经递质起作用的神经细胞树突之间的空间很小
- 内分泌细胞
- 释放参与内分泌信号传导的配体(激素)的细胞
- 内分泌信号
- 由配体(激素)通过生物体循环系统从信号细胞传递到靶细胞的长距离信号
- 酶联受体
- 具有与膜结合酶相关的细胞内结构域的细胞表面受体
- 细胞外域
- 位于细胞表面的细胞表面受体的区域
- G 蛋白连接受体
- 细胞表面受体,它激活膜结合的 G 蛋白,将信号从受体传递到附近的膜成分
- 细胞间信号传导
- 细胞之间的通信
- 内部接收器
- (也称为细胞内受体)受体蛋白,位于细胞的细胞溶胶中,与穿过质膜的配体结合
- 细胞内介质
- (也是第二个信使)在细胞内传输信号的小分子
- 细胞内信号传导
- 细胞内通信
- 离子通道连接受体
- 形成质膜通道的细胞表面受体,当配体与细胞外结构域(配体门控通道)结合时,质膜通道打开
- 配体
- 由信号传导细胞产生的分子,该分子与特定受体结合,在此过程中传递信号
- 神经递质
- 将信号从一个神经细胞传送到另一个神经细胞的化学配体
- 旁分泌信号
- 附近细胞之间的信号,由在细胞间空间的液体介质中传播的配体传递
- 接收器
- 与配体结合的靶细胞内或之上的蛋白质
- 信号细胞
- 释放信号分子的细胞,允许与另一个细胞通信
- 突触信号
- 在神经细胞之间传播的化学信号(神经递质)
- 靶细胞
- 具有来自信号细胞的信号或配体的受体的细胞