
8.3: التنفس الخلوي
- Page ID
- 194744

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)أهداف التعلم
- قارن وقارن بين موقع ووظيفة نظام نقل الإلكترون في خلية بدائية النواة وخلية حقيقية النواة
- قارن وقارن بين الاختلافات بين مستوى الركيزة والفسفرة التأكسدية
- شرح العلاقة بين التناضح الكيميائي والقوة المحركة للبروتون
- وصف وظيفة وموقع سينثاز ATP في خلية بدائية النواة مقابل خلية حقيقية النواة
- قارن وقارن بين التنفس الهوائي واللاهوائي
لقد ناقشنا للتو مسارين في هدم الجلوكوز - تحلل السكر ودورة كريبس - اللذان يولدان ATP عن طريق الفسفرة على مستوى الركيزة. ومع ذلك، يتم إنتاج معظم ATP خلال عملية منفصلة تسمى الفسفرة التأكسدية، والتي تحدث أثناء التنفس الخلوي. يبدأ التنفس الخلوي عندما تنتقل الإلكترونات من NADH و FADH 2 - المصنوع في تحلل السكر، والتفاعل الانتقالي، ودورة كريبس - من خلال سلسلة من التفاعلات الكيميائية إلى متقبل الإلكترون غير العضوي النهائي (إما الأكسجين في التنفس الهوائي أو الجزيئات غير العضوية غير الأكسجين في التنفس اللاهوائي). تتم عمليات نقل الإلكترون هذه في الجزء الداخلي من غشاء الخلية للخلايا بدائية النواة أو في مجمعات بروتينية متخصصة في الغشاء الداخلي للميتوكوندريا للخلايا حقيقية النواة. يتم حصاد طاقة الإلكترونات لتوليد تدرج كهروكيميائي عبر الغشاء، والذي يستخدم لصنع ATP عن طريق الفسفرة التأكسدية.
نظام نقل الإلكترون
نظام نقل الإلكترون (ETS) هو المكون الأخير الذي يشارك في عملية التنفس الخلوي؛ وهو يتألف من سلسلة من مجمعات البروتين المرتبطة بالغشاء وناقلات الإلكترون الملحقة المتنقلة المرتبطة بها. نقل الإلكترون عبارة عن سلسلة من التفاعلات الكيميائية التي تشبه لواء الجرافة حيث يتم تمرير الإلكترونات من NADH و FADH 2 بسرعة من حامل إلكتروني ETS إلى التالي. يمكن لهذه الناقلات تمرير الإلكترونات في ETS بسبب إمكانات الأكسدة والاختزال. لكي يقبل البروتين أو المادة الكيميائية الإلكترونات، يجب أن يكون لها قدرة أكسدة أكثر إيجابية من المتبرع بالإلكترون. لذلك، تنتقل الإلكترونات من حاملات الإلكترون ذات إمكانات الأكسدة والاختزال السلبية الأكبر إلى تلك التي لديها إمكانات الأكسدة والاختزال الأكثر إيجابية. الفئات الأربع الرئيسية لناقلات الإلكترون المشاركة في كل من أنظمة نقل الإلكترون حقيقية النواة وبدائية النواة هي السيتوكرومات والبروتينات المفلورة وبروتينات الحديد والكونونات.
في التنفس الهوائي، يكون مستقبل الإلكترون النهائي (أي الذي يتمتع بقدرة الأكسدة والاختزال الأكثر إيجابية) في نهاية ETS هو جزيء الأكسجين (O 2) الذي يتحول إلى ماء (H 2 O) بواسطة حامل ETS النهائي. يختلف ناقل الإلكترون هذا، السيتوكروم أوكسيديز، بين أنواع البكتيريا ويمكن استخدامه للتمييز بين البكتيريا ذات الصلة الوثيقة للتشخيص. على سبيل المثال، يستخدم دواء Pseudomonas aeruginosa الانتهازي الذي لا يحتوي على الجرام وداء الكوليريا المسبب للكوليرا الذي يسبب الجرام أوكسيديز السيتوكروم C، الذي يمكن اكتشافه عن طريق اختبار الأوكسيداز، في حين أن البكتيريا المعوية الأخرى التي لا تحمل الجرام، مثل E. coli، تعتبر سلبية لهذا الغرض. اختبار لأنها تنتج أنواعًا مختلفة من أوكسيديز السيتوكروم.
هناك العديد من الحالات التي لا يمكن فيها التنفس الهوائي، بما في ذلك أي واحد أو أكثر مما يلي:
- تفتقر الخلية إلى جينات ترميز أوكسيديز السيتوكروم المناسب لنقل الإلكترونات إلى الأكسجين في نهاية نظام نقل الإلكترون.
- تفتقر الخلية إلى الجينات التي تقوم بترميز الإنزيمات لتقليل الآثار الضارة للغاية لجذور الأكسجين الخطرة الناتجة أثناء التنفس الهوائي، مثل بيروكسيد الهيدروجين (H 2 O 2) أو الأكسيد الفائق\((\ce{O2-})\).
- تفتقر الخلية إلى كمية كافية من الأكسجين لإجراء التنفس الهوائي.
أحد البدائل الممكنة للتنفس الهوائي هو التنفس اللاهوائي، باستخدام جزيء غير عضوي غير الأكسجين كمستقبل للإلكترون النهائي. توجد أنواع عديدة من التنفس اللاهوائي في البكتيريا والعتيقة. تعتبر أجهزة إزالة النتروجين من بكتيريا التربة المهمة التي تستخدم النترات\((\ce{NO3-})\) والنتريت\((\ce{NO2-})\) كمستقبلات نهائية للإلكترون، وتنتج غاز النيتروجين (N 2). تتحول العديد من البكتيريا التي تتنفس هوائيًا، بما في ذلك E. coli، إلى استخدام النترات كمستقبل نهائي للإلكترون وإنتاج النتريت عند استنفاد مستويات الأكسجين.
عادةً ما تحتوي الميكروبات التي تستخدم التنفس اللاهوائي على دورة كريبس سليمة، لذلك يمكن لهذه الكائنات الحية الوصول إلى طاقة جزيئات NADH و FADH 2 المتكونة. ومع ذلك، تستخدم أجهزة التنفس اللاهوائية حاملات ETS المعدلة المشفرة بواسطة جينوماتها، بما في ذلك المجمعات المميزة لنقل الإلكترون إلى متقبل الإلكترون النهائي. يتم إنشاء تدرجات كهروكيميائية أصغر من أنظمة نقل الإلكترون هذه، لذلك يتم تكوين كميات أقل من ATP من خلال التنفس اللاهوائي.
التمارين الرياضية\(\PageIndex{1}\)
هل يستخدم كل من التنفس الهوائي والتنفس اللاهوائي سلسلة نقل إلكترونية؟
التناضح الكيميائي والقوة المحركة للبروتون والفسفرة التأكسدية
في كل عملية نقل للإلكترون عبر ETS، يفقد الإلكترون الطاقة، ولكن مع بعض عمليات النقل، يتم تخزين الطاقة كطاقة محتملة باستخدامها لضخ أيونات الهيدروجين (H +) عبر الغشاء. في الخلايا بدائية النواة، يتم ضخ H + إلى خارج الغشاء السيتوبلازمي (يسمى الفضاء المحيط بالبكتيريا سالبة الجرام وإيجابية الجرام)، وفي الخلايا حقيقية النواة، يتم ضخها من مصفوفة الميتوكوندريا عبر غشاء الميتوكوندريا الداخلي إلى الفضاء بين الأغشية. يوجد توزيع غير متساوٍ لـ H + عبر الغشاء الذي ينشئ تدرجًا كهروكيميائيًا لأن أيونات H + مشحونة بشكل إيجابي (كهربائي) وهناك تركيز أعلى (كيميائي) على جانب واحد من الغشاء. يشار إلى هذا التدرج الكهروكيميائي المتكون من تراكم H + (المعروف أيضًا باسم البروتون) على جانب واحد من الغشاء مقارنة بالآخر باسم القوة المحركة للبروتون (PMF). نظرًا لأن الأيونات المعنية هي H +، يتم أيضًا تحديد تدرج درجة الحموضة، حيث يكون جانب الغشاء الذي يحتوي على تركيز أعلى من H+ أكثر حمضية. بالإضافة إلى استخدام PMF لصنع ATP، كما تمت مناقشته في هذا الفصل، يمكن أيضًا استخدام PMF لقيادة عمليات أخرى غير مواتية للطاقة، بما في ذلك نقل المغذيات ودوران السوط من أجل الحركة.
تتسبب الطاقة الكامنة لهذا التدرج الكهروكيميائي الناتج عن ETS في انتشار H + عبر الغشاء (غشاء البلازما في الخلايا بدائية النواة والغشاء الداخلي في الميتوكوندريا في الخلايا حقيقية النواة). يجب أن يحدث هذا التدفق لأيونات الهيدروجين عبر الغشاء، والذي يسمى التناضح الكيميائي، من خلال قناة في الغشاء عبر مركب إنزيم مرتبط بالغشاء يسمى ATP synthase (الشكل\(\PageIndex{1}\)). إن الميل إلى الحركة بهذه الطريقة يشبه إلى حد كبير المياه المتراكمة على جانب واحد من السد، والتي تتحرك عبر السد عند فتحه. إن سينثاز ATP (مثل مزيج من مدخل ومولد السد الكهرومائي) هو بروتين معقد يعمل كمولد صغير، ويقلب بقوة H + المنتشر عبر الإنزيم، ويخفض تدرجه الكهروكيميائي من حيث يوجد العديد من الصد المتبادل لـ H + إلى حيث يوجد عدد أقل من H +. في الخلايا بدائية النواة، يتدفق H + من خارج الغشاء السيتوبلازمي إلى السيتوبلازم، بينما في الميتوكوندريا حقيقية النواة، يتدفق H + من الفضاء بين الأغشية إلى مصفوفة الميتوكوندريا. يؤدي تدوير أجزاء هذه الآلة الجزيئية إلى تجديد ATP من ADP والفوسفات غير العضوي (P i) عن طريق الفسفرة التأكسدية، وهي آلية ثانية لصنع ATP الذي يحصد الطاقة الكامنة المخزنة في التدرج الكهروكيميائي.
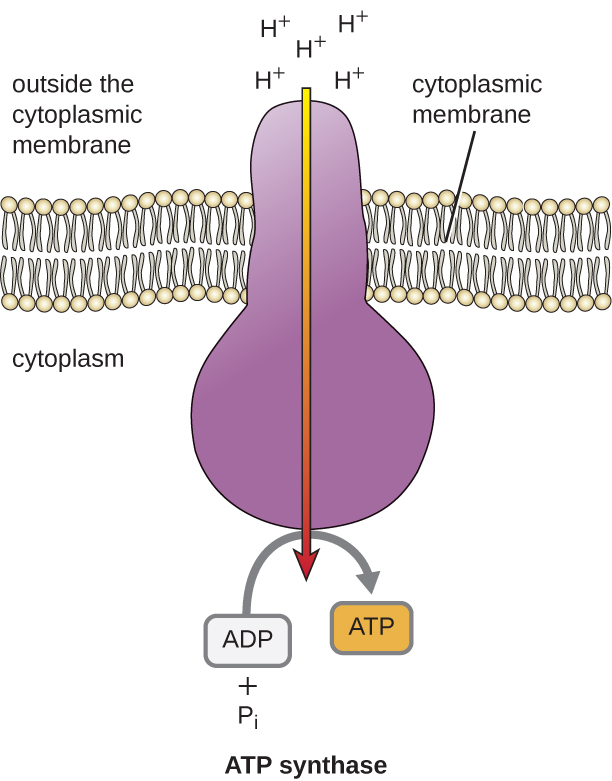
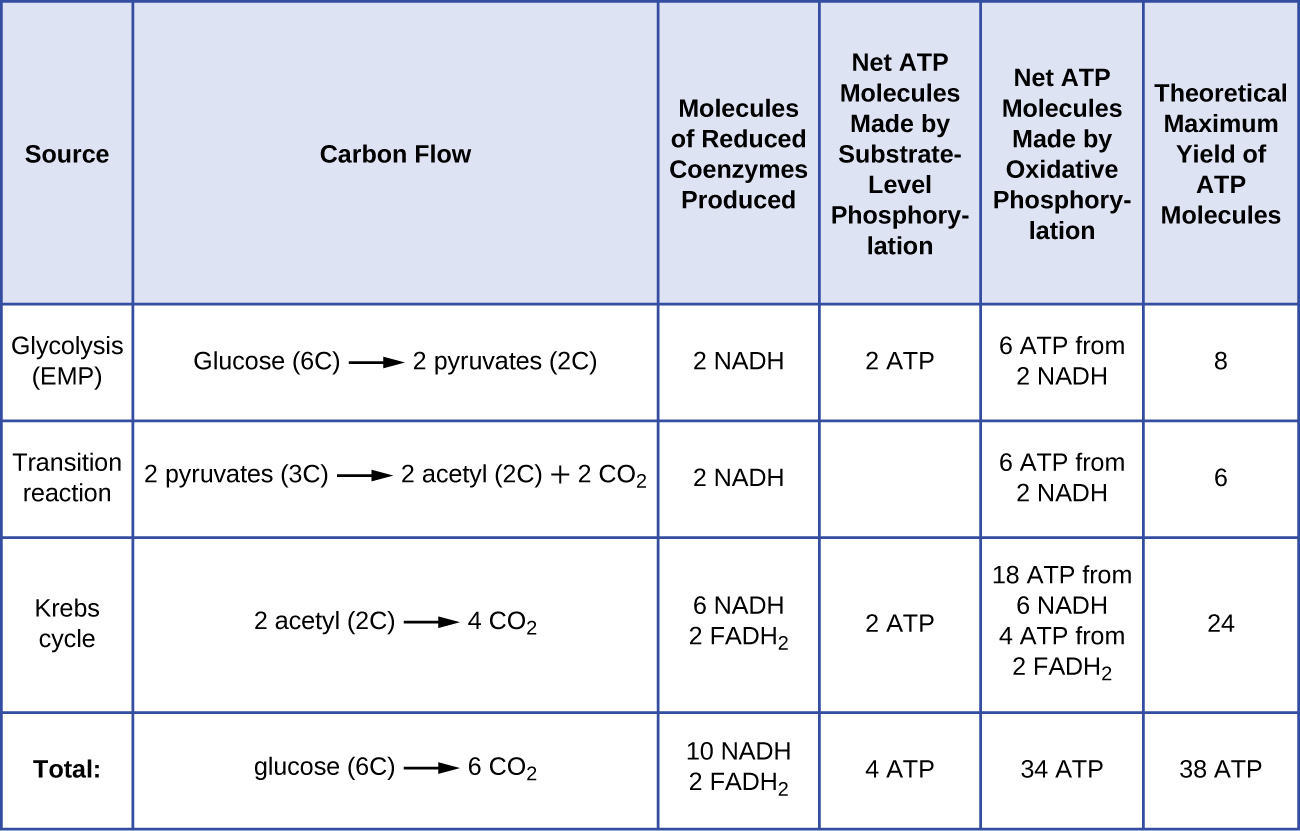
يختلف عدد جزيئات ATP الناتجة عن تقويض الجلوكوز. على سبيل المثال، يختلف عدد أيونات الهيدروجين التي يمكن لمجمعات نظام نقل الإلكترون ضخها عبر الغشاء بين الأنواع المختلفة من الكائنات الحية. في التنفس الهوائي في الميتوكوندريا، يولد مرور الإلكترونات من جزيء واحد من NADH قوة دافعة كافية للبروتون لتكوين ثلاثة جزيئات ATP عن طريق الفسفرة التأكسدية، في حين أن مرور الإلكترونات من جزيء واحد من FADH 2 يولد قوة دافعة كافية للبروتون لصنع اثنين فقط جزيئات ATP. وبالتالي، فإن جزيئات NADH العشرة التي يتم إنتاجها لكل جلوكوز أثناء تحلل السكر، والتفاعل الانتقالي، ودورة كريبس تحمل طاقة كافية لصنع 30 جزيء ATP، في حين أن جزيئي FADH 2 المصنوع من كل جلوكوز خلال هذه العمليات يوفران طاقة كافية لصنع أربعة جزيئات ATP. بشكل عام، يبلغ الحد الأقصى للعائد النظري لـ ATP الذي يتم تحقيقه أثناء التنفس الهوائي الكامل للجلوكوز 38 جزيئًا، أربعة منها يتم تصنيعها عن طريق الفسفرة على مستوى الركيزة و 34 بواسطة الفسفرة التأكسدية (الشكل\(\PageIndex{2}\)). في الواقع، عادة ما يكون إجمالي إنتاج ATP أقل، حيث يتراوح من جزيء واحد إلى 34 جزيء ATP، اعتمادًا على ما إذا كانت الخلية تستخدم التنفس الهوائي أو التنفس اللاهوائي؛ في الخلايا حقيقية النواة، يتم إنفاق بعض الطاقة لنقل المواد الوسيطة من السيتوبلازم إلى الميتوكوندريا، مما يؤثر على عائد ATP.
\(\PageIndex{2}\)يلخص الشكل الحد الأقصى للعائدات النظرية لـ ATP من العمليات المختلفة أثناء التنفس الهوائي الكامل لجزيء جلوكوز واحد.
التمارين الرياضية\(\PageIndex{1}\)
ما هي وظائف القوة المحركة للبروتون؟
ملخص
- يتم إنتاج معظم ATP الناتج أثناء التنفس الخلوي للجلوكوز عن طريق الفسفرة التأكسدية.
- يتكون نظام نقل الإلكترون (ETS) من سلسلة من مجمعات البروتين المرتبطة بالغشاء وناقلات الإلكترون الملحقة المتنقلة المرتبطة بها. تم تضمين ETS في الغشاء السيتوبلازمي لبدائيات النواة وغشاء الميتوكوندريا الداخلي لحقيقيات النوى.
- يحتوي كل مركب من مركبات ETS على إمكانات اختزال مختلفة، وتنتقل الإلكترونات من حاملات الإلكترون ذات إمكانات الأكسدة والاختزال السلبية إلى تلك التي تتمتع بإمكانيات الأكسدة والاختزال الأكثر إيجابية.
- لإجراء التنفس الهوائي، تتطلب الخلية الأكسجين باعتباره مستقبل الإلكترون النهائي. تحتاج الخلية أيضًا إلى دورة كريبس كاملة، وأكسيداز السيتوكروم المناسب، وإنزيمات إزالة السموم بالأكسجين لمنع الآثار الضارة لجذور الأكسجين الناتجة أثناء التنفس الهوائي.
- تستخدم الكائنات الحية التي تقوم بالتنفس اللاهوائي حاملات نظام نقل الإلكترون البديلة للنقل النهائي للإلكترونات إلى متقبل الإلكترون النهائي غير الأكسجين.
- تُظهر الميكروبات تباينًا كبيرًا في تكوين أنظمة نقل الإلكترون الخاصة بها، والتي يمكن استخدامها لأغراض التشخيص للمساعدة في تحديد بعض مسببات الأمراض.
- عندما يتم تمرير الإلكترونات من NADH و FADH 2 من خلال ETS، يفقد الإلكترون الطاقة. يتم تخزين هذه الطاقة من خلال ضخ H + عبر الغشاء، مما يولد قوة دافعة للبروتون.
- يمكن تسخير طاقة هذه القوة المحركة للبروتونات من خلال السماح لأيونات الهيدروجين بالانتشار مرة أخرى عبر الغشاء عن طريق التناضح الكيميائي باستخدام مركب ATP synthase. عندما تنتشر أيونات الهيدروجين عبر التدرج الكهروكيميائي، تدور مكونات سينثاز ATP، مما يجعل ATP من ADP و P i عن طريق الفسفرة التأكسدية.
- يشكل التنفس الهوائي المزيد من ATP (بحد أقصى 34 جزيء ATP) أثناء الفسفرة التأكسدية مقارنة بالتنفس اللاهوائي (بين جزيء واحد و 32 جزيء ATP).