
12.3: قوانين الميراث
- Page ID
- 196491

المهارات اللازمة للتطوير
- شرح قانون مندل للفصل والتشكيلة المستقلة من حيث علم الوراثة وأحداث الانقسام الاختزالي
- استخدم طريقة الخط المتشعب وقواعد الاحتمالات لحساب احتمالية الأنماط الجينية والأنماط الظاهرية من تقاطعات الجينات المتعددة
- شرح تأثير الربط وإعادة التركيب على الأنماط الجينية المشيجية
- شرح النتائج المظهرية للتأثيرات الشرباطية بين الجينات
قام ميندل بتعميم نتائج تجاربه على نبات البازلاء في أربعة افتراضات، يُطلق على بعضها أحيانًا اسم «القوانين»، والتي تصف أساس الوراثة السائدة والمتنحية في الكائنات ثنائية الصبغيات. كما تعلمت، توجد امتدادات أكثر تعقيدًا من Mendelism لا تظهر نفس النسب المظهرية لـ F 2 (3:1). ومع ذلك، تلخص هذه القوانين أساسيات علم الوراثة الكلاسيكي.
أزواج من عوامل الوحدة أو الجينات
اقترح مندل أولاً أن عوامل الوحدة المزدوجة للوراثة تنتقل بأمانة من جيل إلى جيل عن طريق تفكك العوامل المزدوجة وإعادة ربطها أثناء تكوين الأمشاج والتخصيب، على التوالي. بعد أن تخطى البازلاء بصفات متناقضة ووجد أن السمة المتنحية عادت إلى الظهور في جيل F 2، استنتج مندل أن العوامل الوراثية يجب أن تكون موروثة كوحدات منفصلة. تناقضت هذه النتيجة مع الاعتقاد في ذلك الوقت بأن الصفات الأبوية كانت مختلطة في النسل.
يمكن أن تكون الأليلات مهيمنة أو متنحية
ينص قانون مندل للهيمنة على أنه في الزيجوت غير المتجانس، ستخفي إحدى السمات وجود سمة أخرى لنفس الخاصية. بدلاً من أن يساهم كلا الأليلين في النمط الظاهري، سيتم التعبير عن الأليل السائد حصريًا. سيظل الأليل المتنحي «كامنًا» ولكنه سينتقل إلى النسل بنفس الطريقة التي ينتقل بها الأليل السائد. لن يتم التعبير عن السمة المتنحية إلا من خلال النسل الذي يحتوي على نسختين من هذا الأليل (الشكل\(\PageIndex{1}\))، وسوف تتكاثر هذه النسل بشكل حقيقي عند التزاوج الذاتي.
منذ تجارب مندل على نباتات البازلاء، وجد باحثون آخرون أن قانون الهيمنة لا يصح دائمًا. بدلاً من ذلك، تم العثور على عدة أنماط مختلفة من الميراث.

الفصل المتساوي للأليلات
لاحظ مندل أن نباتات البازلاء ذات التكاثر الحقيقي ذات الصفات المتناقضة أدت إلى ظهور أجيال F 1 التي عبرت جميعها عن السمة السائدة وأجيال F 2 التي عبرت عن السمات السائدة والمتنحية بنسبة 3:1، اقترح Mendel قانون الفصل. ينص هذا القانون على أن عوامل الوحدة المزدوجة (الجينات) يجب أن تنفصل بالتساوي في الأمشاج بحيث يكون لدى النسل احتمال متساوٍ لوراثة أي عامل. بالنسبة للجيل F 2 من الصليب أحادي الهجين، يمكن أن ينتج عن ذلك التوليفات الثلاثة الممكنة التالية من الأنماط الجينية: متماثل الزيجوت السائد أو غير المتجانس أو متنحي متماثل الزيجوت. نظرًا لأن الزيجوت غير المتجانسة يمكن أن تنشأ من مسارين مختلفين (تلقي أليل مهيمن وآخر متنحي من أي من الوالدين)، ولأن الزيجوت غير المتجانسة والأفراد المهيمنين متماثلين متماثلون ظاهريًا، فإن القانون يدعم نسبة النمط الظاهري التي لاحظها مندل 3:1. الفصل المتساوي للأليلات هو السبب في أننا نستطيع تطبيق مربع Punnett للتنبؤ بدقة بنسل الآباء ذوي الأنماط الجينية المعروفة. الأساس المادي لقانون ميندل للفصل هو القسم الأول من الانقسام الاختزالي، حيث يتم فصل الكروموسومات المتماثلة بإصداراتها المختلفة من كل جين إلى نوى ابنة. لم يفهم المجتمع العلمي دور الفصل الميولوجي للكروموسومات في التكاثر الجنسي خلال حياة مندل.
تشكيلة مستقلة
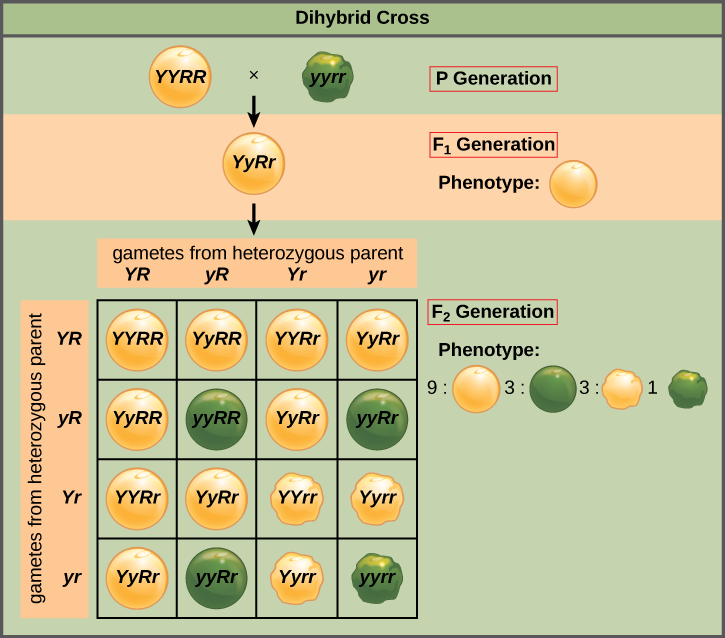
ينص قانون مندل للتشكيلة المستقلة على أن الجينات لا تؤثر على بعضها البعض فيما يتعلق بفرز الأليلات إلى الأمشاج، ومن المرجح أن يحدث كل مزيج ممكن من الأليلات لكل جين بنفس القدر. يمكن توضيح التشكيلة المستقلة للجينات من خلال الصليب ثنائي الهجين، وهو عبارة عن تقاطع بين أبوين يتكاثران حقيقيان يعبران عن سمات مختلفة لخاصيتين. ضع في اعتبارك خصائص لون البذور وملمس البذور لنباتين من البازلاء، أحدهما يحتوي على بذور خضراء مجعدة (yyrr) والآخر يحتوي على بذور صفراء مستديرة (YYRR). نظرًا لأن كل من الوالدين متماثل الزيجوت، فإن قانون الفصل يشير إلى أن الأمشاج الخاصة بالنبات الأخضر/المجعد كلها سنوات، وأن الأمشاج الخاصة بالنبات الأصفر/المستدير كلها YR. لذلك، فإن جيل F 1 من النسل جميعًا هو yyrr (الشكل\(\PageIndex{2}\)).
التمارين
في نباتات البازلاء، تهيمن الأزهار الأرجوانية (P) على الزهور البيضاء (p) وتهيمن البازلاء الصفراء (Y) على البازلاء الخضراء (y). ما هي الأنماط الجينية والأنماط الظاهرية المحتملة للتقاطع بين نباتات البازلاء PpYy و PpYy؟ ما عدد المربعات التي تحتاج إليها لإجراء تحليل مربع بونيت لهذا الصليب؟
- إجابة
-
الأنماط الجينية المحتملة هي PpyY و PpyY و PpyY. سيؤدي النمطين الوراثيين السابقين إلى نباتات ذات أزهار أرجوانية وبازلاء صفراء، في حين أن النمطين الوراثيين الأخيرين سينتج عنهما نباتات ذات أزهار بيضاء مع البازلاء الصفراء، بنسبة 1:1 لكل نمط ظاهري. تحتاج فقط إلى مربع Punnett 2 × 2 (إجمالي أربعة مربعات) لإجراء هذا التحليل لأن اثنين من الأليلات متجانسان.
بالنسبة لجيل F2، يتطلب قانون الفصل أن يتلقى كل مشيج إما أليل R أو أليل R جنبًا إلى جنب مع أليل Y أو أليل y. ينص قانون التشكيلة المستقلة على أن المشيج الذي يتم فرز الأليل فيه من المحتمل أيضًا أن يحتوي إما على أليل Y أو أليل y. وبالتالي، هناك أربعة أنواع من الأمشاج ذات الاحتمالية المتساوية يمكن تشكيلها عندما يتم عبور الزيجوت غير المتجانس Yyrr ذاتيًا، على النحو التالي: YR و Yr و yr. يمنحنا ترتيب هذه الأمشاج على طول الجزء العلوي واليسار من مربع بونيت 4 × 4 (الشكل\(\PageIndex{2}\)) 16 تركيبة جينية متساوية الاحتمال. من هذه الأنماط الجينية، نستنتج نسبة ظاهرية تبلغ 9 مستدير/أصفر: 3 دائري/أخضر: 3 متجعد/أصفر:1 متجعد/أخضر (الشكل\(\PageIndex{2}\)). هذه هي نسب النسل التي نتوقعها، بافتراض أننا أجرينا عمليات التهجين بحجم عينة كبير بما يكفي.
بسبب التنوع المستقل والهيمنة، يمكن تقسيم النسبة المظهرية ثنائية الهجين 9:3:3:1 إلى نسبتين 3:1، وهي سمة مميزة لأي تقاطع أحادي الهجين يتبع نمطًا مهيمنًا ومتنحًا. بتجاهل لون البذور والنظر فقط في نسيج البذور في الصليب ثنائي الهجين أعلاه، نتوقع أن يكون ثلاثة أرباع نسل جيل F 2 مستديرًا، وربعه سيكون مجعدًا. وبالمثل، عند عزل لون البذور فقط، نفترض أن ثلاثة أرباع نسل F 2 سيكون أصفر وربعه سيكون أخضر. يعد فرز الأليلات من حيث الملمس واللون أحداثًا مستقلة، لذا يمكننا تطبيق قاعدة المنتج. لذلك، من المتوقع أن تكون نسبة نسل F 2 المستدير والأصفر (3/4) × (3/4) = 9/16، ومن المتوقع أن تكون نسبة النسل المجعد والأخضر (1/4) × (1/4) = 1/16. هذه النسب مماثلة لتلك التي تم الحصول عليها باستخدام مربع Punnett. يمكن أيضًا حساب النسل المستدير والأخضر والمجعد والأصفر باستخدام قاعدة المنتج، حيث يشتمل كل من هذه الأنماط الجينية على نمط ظاهري واحد مهيمن وآخر متنحي. لذلك، يتم حساب نسبة كل منها على النحو (3/4) × (1/4) = 3/16.
يشير قانون التشكيلة المستقلة أيضًا إلى أن التقاطع بين الوالدين الأصفر المتجعد (YyRr) والأخضر المستدير (yyRr) سينتج نفس نسل F 1 و F 2 كما هو الحال في صليب YYRR x yyrr.
يكمن الأساس المادي لقانون التشكيلة المستقلة أيضًا في الانقسام الاختزالي الأول، حيث تصطف الأزواج المتماثلة المختلفة في اتجاهات عشوائية. يمكن أن يحتوي كل مشيج على أي مزيج من الكروموسومات الأبوية والأمومية (وبالتالي الجينات الموجودة عليها) لأن اتجاه الرباعيات على مستوى الميتافيز عشوائي.
طريقة الخط المتشعب
عندما يتم النظر في أكثر من جينين، تصبح طريقة Punnett-square غير عملية. على سبيل المثال، يتطلب فحص الصليب الذي يتضمن أربعة جينات شبكة 16 × 16 تحتوي على 256 صندوقًا. سيكون إدخال كل نمط وراثي يدويًا أمرًا مرهقًا للغاية. بالنسبة للتقاطعات الأكثر تعقيدًا، تُفضل طرق الخط المتشعب والاحتمالية.
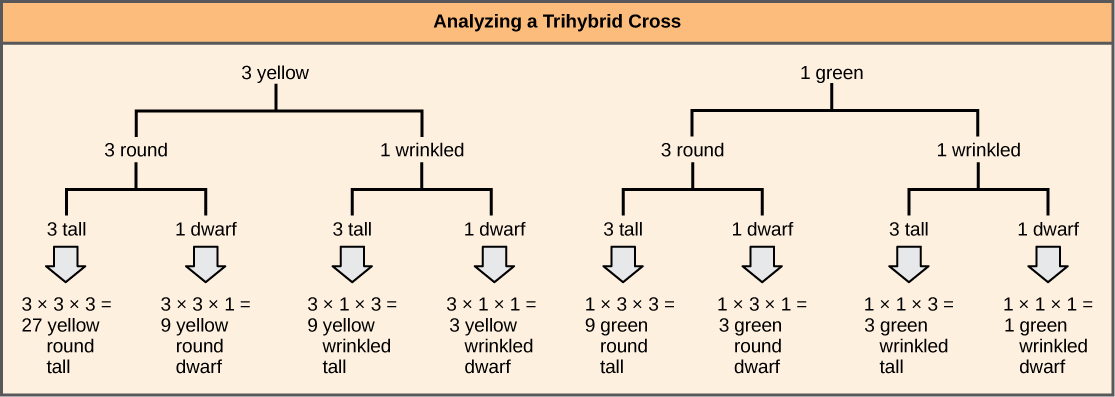
لإعداد رسم تخطيطي للخط المتشعب للتقاطع بين الزيجوت غير المتجانس F 1 الناتج عن التقاطع بين الوالدين AABBCC و aabbcc، نقوم أولاً بإنشاء صفوف تساوي عدد الجينات التي يتم أخذها في الاعتبار، ثم نفصل الأليلات في كل صف على الخطوط المتشعبة وفقًا لـ احتمالات التقاطعات الفردية أحادية الهجين (الشكل\(\PageIndex{3}\)). نقوم بعد ذلك بضرب القيم على طول كل مسار متشعب للحصول على احتمالات ذرية F 2. لاحظ أن هذه العملية هي نسخة تخطيطية من قاعدة المنتج. يمكن ضرب القيم على طول كل مسار متشعب لأن كل جين يتنوع بشكل مستقل. بالنسبة إلى الصليب الثلاثي الهجين، تكون النسبة المظهرية لـ F 2 هي 27:9:9:3:3:3:1.
طريقة الاحتمالية
في حين أن طريقة الخط المتشعب هي طريقة تخطيطية لتتبع الاحتمالات في الصليب، فإن طريقة الاحتمالية تعطي نسب النسل المتوقع أن يُظهر كل نمط ظاهري (أو نمط جيني) دون المساعدة البصرية المضافة. تستخدم كلتا الطريقتين قاعدة المنتج وتأخذان في الاعتبار الأليلات لكل جين على حدة. في وقت سابق، قمنا بفحص النسب المظهرية للتقاطع الثلاثي الهجين باستخدام طريقة الخط المتشعب؛ والآن سنستخدم طريقة الاحتمال لفحص النسب الجينية للتقاطع مع المزيد من الجينات.
بالنسبة إلى الصليب ثلاثي الهجين، تعد كتابة طريقة الخط المتشعب أمرًا شاقًا، وإن لم يكن شاقًا مثل استخدام طريقة Punnett-square. ومع ذلك، لإثبات قوة طريقة الاحتمالية بشكل كامل، يمكننا التفكير في حسابات جينية محددة. على سبيل المثال، بالنسبة للتقاطع رباعي الهجين بين الأفراد غير المتجانس الزيجوت لجميع الجينات الأربعة، والذي يتم فيه فرز جميع الجينات الأربعة بشكل مستقل وبنمط سائد ومتنحي، ما هي نسبة النسل المتوقع أن تكون متنحية متجانسة الزيجوت لجميع الأليلات الأربعة؟ بدلاً من كتابة كل نمط وراثي ممكن، يمكننا استخدام طريقة الاحتمال. نحن نعلم أنه بالنسبة لكل جين، سيكون جزء النسل المتنحي متماثل الزيجوت 1/4. لذلك، بضرب هذا الكسر لكل من الجينات الأربعة، (1/4) × (1/4) × (1/4) × (1/4)، نحدد أن 1/256 من النسل سيكون متنحًا متجانسًا أربع مرات.
بالنسبة لنفس الصليب رباعي الهجين، ما النسبة المتوقعة من النسل الذي له النمط الظاهري السائد في جميع المواقع الأربعة؟ يمكننا الإجابة على هذا السؤال باستخدام النسب المظهرية، ولكن دعونا نفعل ذلك بالطريقة الصعبة - باستخدام النسب الجينية. يسأل السؤال عن نسبة النسل التي هي 1) متماثل الزيجوت السائد عند A أو غير المتجانس عند A، و 2) متماثل الزيجوت عند B أو غير المتجانس عند B، وما إلى ذلك. توضح ملاحظة «أو» و «و» في كل ظرف من الظروف مكان تطبيق قواعد المبلغ والمنتج. احتمال وجود متماثل الزيجوت عند A هو 1/4 واحتمال وجود زيجوت متغاير عند A هو 1/2. احتمال تماثل الزيجوت أو الزيجوت المتغاير هو 1/4 + 1/2 = 3/4 باستخدام قاعدة المجموع. يمكن الحصول على نفس الاحتمال بنفس الطريقة لكل من الجينات الأخرى، بحيث يكون احتمال النمط الظاهري السائد عند A و B و C و D، باستخدام قاعدة المنتج، يساوي 3/4 × 3/4 × 3/4 × 3/4 × 3/4 أو 27/64. إذا لم تكن متأكدًا أبدًا من كيفية الجمع بين الاحتمالات، فإن العودة إلى طريقة الخط المتشعب يجب أن توضح الأمر.
قواعد التسميد متعدد الهجينة
إن التنبؤ بالأنماط الجينية والأنماط الظاهرية للنسل من صلبان معينة هو أفضل طريقة لاختبار معرفتك بعلم الوراثة المندلية. نظرًا لوجود صليب متعدد الهجينة يخضع لتشكيلة مستقلة ويتبع نمطًا مهيمنًا ومتنحًا، توجد العديد من القواعد المعممة؛ يمكنك استخدام هذه القواعد للتحقق من نتائجك أثناء عملك من خلال الحسابات الجينية (الجدول\(\PageIndex{1}\)). لتطبيق هذه القواعد، يجب عليك أولاً تحديد n، عدد أزواج الجينات غير المتجانسة (عدد الجينات التي تفصل بين أليلين لكل منهما). على سبيل المثال، يحتوي التقاطع بين AaBb و AabB غير المتجانسة على n من 2. في المقابل، يحتوي التقاطع بين AaBaB و AaBb على n of 1 لأن A ليس متغاير الزيجوت.
| قاعدة عامة | عدد أزواج الجينات غير المتجانسة |
|---|---|
| عدد الأمشاج المختلفة لـ F1 | 2 ن |
| عدد الأنماط الجينية المختلفة لـ F 2 | رقم 3 |
| بالنظر إلى الميراث السائد والمنحي، فإن عدد الأنماط الظاهرية المختلفة لـ F 2 | 2 ن |
الجينات المرتبطة تنتهك قانون التشكيلة المستقلة
على الرغم من أن جميع خصائص البازلاء في Mendel تصرفت وفقًا لقانون التشكيلة المستقلة، إلا أننا نعلم الآن أن بعض تركيبات الأليل لا يتم توريثها بشكل مستقل عن بعضها البعض. الجينات الموجودة على كروموسومات منفصلة غير متماثلة ستقوم دائمًا بالفرز بشكل مستقل. ومع ذلك، يحتوي كل كروموسوم على مئات أو آلاف الجينات، المنظمة خطيًا على الكروموسومات مثل الخرز على الخيط. يمكن أن يتأثر فصل الأليلات إلى الأمشاج بالارتباط، حيث تكون الجينات الموجودة جسديًا بالقرب من بعضها البعض على نفس الكروموسوم أكثر عرضة للوراثة كزوج. ومع ذلك، بسبب عملية إعادة التركيب، أو «التقاطع»، من الممكن أن يتصرف جينان على نفس الكروموسوم بشكل مستقل، أو كما لو أنهما غير مرتبطين. لفهم هذا، دعونا ننظر في الأساس البيولوجي لربط الجينات وإعادة التركيب.
تمتلك الكروموسومات المتماثلة نفس الجينات في نفس الترتيب الخطي. قد تختلف الأليلات في أزواج الكروموسومات المتماثلة، لكن الجينات التي تتوافق معها لا تختلف. استعدادًا للقسم الأول من الانقسام الاختزالي، تتكاثر الكروموسومات المتماثلة وتشبك. مثل الجينات الموجودة على المتجانسات التي تتوافق مع بعضها البعض. في هذه المرحلة، تتبادل شرائح الكروموسومات المتماثلة شرائح خطية من المواد الوراثية (الشكل\(\PageIndex{4}\)). هذه العملية تسمى إعادة التركيب، أو التقاطع، وهي عملية وراثية شائعة. نظرًا لأن الجينات تتم محاذاتها أثناء إعادة التركيب، لا يتم تغيير ترتيب الجينات. بدلاً من ذلك، فإن نتيجة إعادة التركيب هي دمج الأليلات الأمومية والأبوية على نفس الكروموسوم. عبر كروموسوم معين، قد تحدث العديد من أحداث إعادة التركيب، مما يتسبب في خلط واسع النطاق للأليلات.
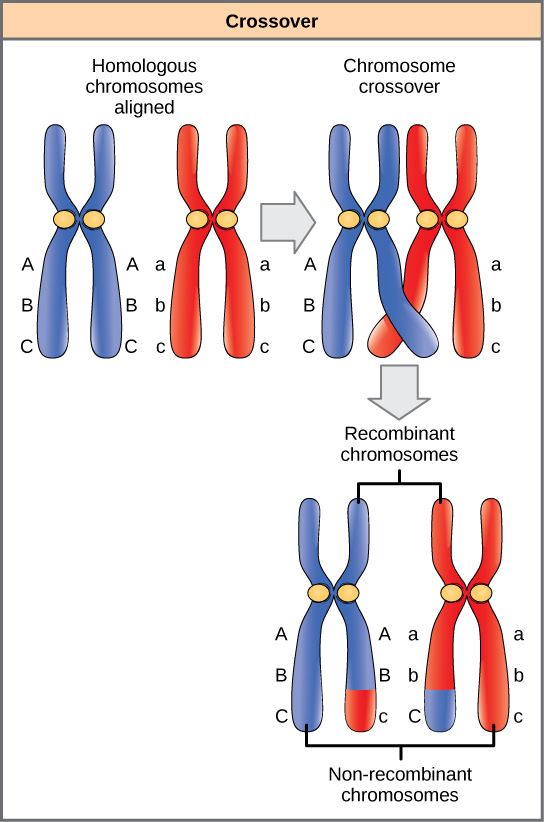
عندما يوجد جينان على مقربة من نفس الكروموسوم، يتم اعتبارهما مرتبطين، وتميل أليلاتهما إلى الانتقال عبر الانقسام الاختزالي معًا. ولتوضيح ذلك، تخيل تقاطعًا ثنائي الهجين يتضمن لون الزهرة وارتفاع النبات حيث تكون الجينات بجوار بعضها البعض على الكروموسوم. إذا كان أحد الكروموسوم المتماثل يحتوي على أليلات للنباتات الطويلة والزهور الحمراء، وكان الكروموسوم الآخر يحتوي على جينات للنباتات القصيرة والزهور الصفراء، فعندما تتشكل الأمشاج، سوف تتجمع الأليلات الطويلة والحمراء معًا في الأمشاج وستنتقل الأليلات القصيرة والصفراء إلى الأمشاج الأخرى. تسمى هذه الأنماط الجينية الأبوية لأنها موروثة سليمة من آباء الفرد المنتج للأمشاج. ولكن على عكس ما إذا كانت الجينات موجودة على كروموسومات مختلفة، فلن تكون هناك أمشاج بأليلات طويلة وصفراء ولن تكون هناك أمشاج بأليلات قصيرة وحمراء. إذا قمت بإنشاء مربع Punnett باستخدام هذه الأمشاج، فسترى أن التنبؤ المندلي الكلاسيكي بنتيجة 9:3:3:1 للصليب المختلط لن ينطبق. مع زيادة المسافة بين جينين، يزداد احتمال حدوث انتقال واحد أو أكثر بينهما، وتتصرف الجينات كما لو كانت على كروموسومات منفصلة. استخدم علماء الوراثة نسبة الأمشاج المؤتلفة (تلك التي لا تشبه الوالدين) كمقياس لمدى تباعد الجينات على الكروموسوم. وباستخدام هذه المعلومات، قاموا بإنشاء خرائط مفصلة للجينات على الكروموسومات للكائنات الحية المدروسة جيدًا، بما في ذلك البشر.
لا يشير منشور مندل الأساسي إلى الربط، وقد تساءل العديد من الباحثين عما إذا كان قد واجه الربط ولكنهم اختاروا عدم نشر تلك المقاطع خشية أن تبطل افتراضاته المتنوعة المستقلة. تحتوي البازلاء في الحديقة على سبعة كروموسومات، وقد أشار البعض إلى أن اختياره لسبع خصائص لم يكن مصادفة. ومع ذلك، حتى لو لم تكن الجينات التي فحصها موجودة على كروموسومات منفصلة، فمن المحتمل أنه ببساطة لم يلاحظ الارتباط بسبب تأثيرات الخلط الواسعة لإعادة التركيب.
اتصال المنهج العلمي: اختبار فرضية التشكيلة المستقلة
ولتقدير مقدار العمل والإبداع الذي استغرقته تجارب مندل بشكل أفضل، تابع إحدى صلبان مندل ثنائية الهجين.
سؤال: ماذا سيكون نسل الصليب ثنائي الهجين؟
الخلفية: ضع في اعتبارك أن نباتات البازلاء تنضج في موسم نمو واحد، ويمكنك الوصول إلى حديقة كبيرة حيث يمكنك زراعة الآلاف من نباتات البازلاء. هناك العديد من نباتات التكاثر الحقيقية ذات الأزواج التالية من السمات: النباتات الطويلة ذات القرون المتضخمة والنباتات القزمة ذات القرون الضيقة. قبل أن تنضج النباتات، تقوم بإزالة الأعضاء المنتجة لحبوب اللقاح من النباتات الطويلة/المنتفخة في الصلبان لمنع الإخصاب الذاتي. عند نضج النبات، يتم تهجين النباتات يدويًا عن طريق نقل حبوب اللقاح من النباتات الأقزام/المقيدة إلى وصمات النباتات الطويلة/المنتفخة.
الفرضية: سيتم فرز كلا الزوجين من السمات بشكل مستقل وفقًا لقوانين المندلية. عندما يتم تهجين الوالدين الذين يتكاثرون حقًا، يكون كل نسل F 1 طويلًا وله قرون منتفخة، مما يشير إلى أن السمات الطويلة والمتضخمة تهيمن على الصفات القزمة والمقيدة، على التوالي. ينتج عن التقاطع الذاتي لـ F 1 غير المتجانسة ذرية 2,000 F 2.
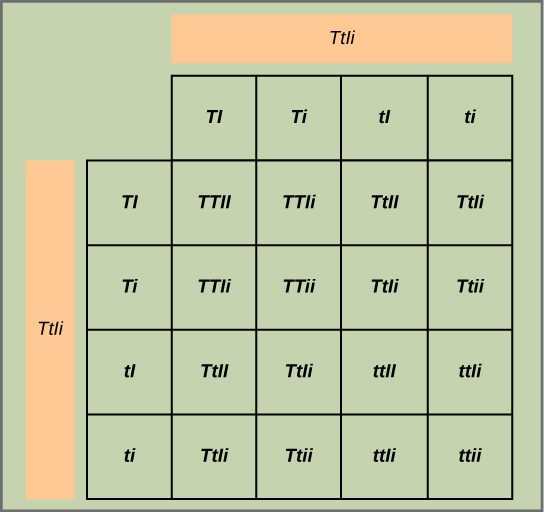
اختبر الفرضية: نظرًا لأن كل زوج من السمات يتم فرزه بشكل مستقل، فمن المتوقع أن تكون نسب الطول:القزم والمتضخم:المقيد 3:1. يُطلق على زوج السمات الطول/القزم اسم T/T، ويتم تعيين زوج السمات المتضخم/المقيد I/i. لذلك فإن كل عضو من جيل F 1 لديه نمط وراثي من tTiI. قم بإنشاء شبكة مماثلة للشكل\(\PageIndex{2}\)، حيث يمكنك عبور اثنين من أفراد TTiI. يمكن لكل فرد التبرع بأربع مجموعات من صفتين: TI أو Ti أو Ti أو Ti أو ti، مما يعني أن هناك 16 احتمالًا للأنماط الجينية للنسل. نظرًا لأن الأليلات T و I هي المهيمنة، فإن أي فرد لديه واحد أو اثنين من هذه الأليلات سيعبر عن الأنماط الظاهرية الطويلة أو المتضخمة، على التوالي، بغض النظر عما إذا كان لديهم أيضًا أليل t أو i. فقط الأفراد الذين هم tt أو ii سيعبرون عن الأليلات القزمية والمقيدة، على التوالي. كما هو موضح في الشكل\(\PageIndex{5}\)، تتوقع أنك ستلاحظ نسب النسل التالية: الطويل/المتضخم:الطويل/المضيق: القمز/المتضخم: القزام/المقيد بنسبة 9:3:3:1. لاحظ من الشبكة أنه عند النظر إلى أزواج السمات الطويلة/القزمية والمتضخمة/المقيدة بشكل منعزل، يتم توريث كل منها بنسب 3:1.
اختبر الفرضية: يمكنك عبور النباتات القزمة والطويلة ثم عبور النسل ذاتيًا. للحصول على أفضل النتائج، يتكرر ذلك مع مئات أو حتى الآلاف من نباتات البازلاء. ما هي الاحتياطات الخاصة التي يجب اتخاذها في الصلبان وفي زراعة النباتات؟
قم بتحليل بياناتك: ستلاحظ الأنماط الظاهرية النباتية التالية في جيل F 2: 2706 طول/منتفخ، 930 طويل/مقيد، 888 قمز/منتفخ، و 300 قمز/مقيد. قلل هذه النتائج إلى نسبة وحدد ما إذا كانت متوافقة مع قوانين Mendelian.
كوِّن استنتاجًا: هل كانت النتائج قريبة من النسبة المظهرية المتوقعة 9:3:3:1؟ هل تدعم النتائج التنبؤ؟ ما الذي يمكن ملاحظته إذا تم استخدام عدد أقل بكثير من النباتات، بالنظر إلى أن الأليلات تنفصل عشوائيًا في الأمشاج؟ حاول أن تتخيل زراعة العديد من نباتات البازلاء، وفكر في احتمال حدوث خطأ تجريبي. على سبيل المثال، ماذا سيحدث إذا كان الجو عاصفًا للغاية يومًا ما؟
الرعاف
أشارت دراسات مندل في نباتات البازلاء إلى أن مجموع النمط الظاهري للفرد يتم التحكم فيه بواسطة الجينات (أو كما سماها، عوامل الوحدة)، بحيث يتم التحكم في كل خاصية بشكل واضح وكامل بواسطة جين واحد. في الواقع، تكون الخصائص الفردية التي يمكن ملاحظتها دائمًا تحت تأثير جينات متعددة (لكل منها أليلين أو أكثر) تعمل في انسجام تام. على سبيل المثال، تساهم ثمانية جينات على الأقل في لون العين لدى البشر.
في بعض الحالات، يمكن أن تساهم العديد من الجينات في جوانب النمط الظاهري الشائع دون أن تتفاعل منتجاتها الجينية بشكل مباشر. في حالة تطور الأعضاء، على سبيل المثال، يمكن التعبير عن الجينات بالتتابع، مع إضافة كل جين إلى تعقيد وخصوصية العضو. قد تعمل الجينات بأشكال تكميلية أو تآزرية، بحيث يلزم التعبير عن جينين أو أكثر في وقت واحد للتأثير على النمط الظاهري. قد تعارض الجينات بعضها البعض أيضًا، حيث يقوم أحد الجينات بتعديل التعبير عن الآخر.
في حالة الرعاف، يكون التفاعل بين الجينات عدائيًا، بحيث يخفي أحد الجينات التعبير عن الآخر أو يتداخل معه. «Epistasis» هي كلمة تتكون من جذور يونانية تعني «الوقوف». يُقال إن الأليلات التي يتم حجبها أو إسكاتها تميل إلى الإيليلات الرسغية التي تقوم بعملية الإخفاء. غالبًا ما يكون الأساس الكيميائي الحيوي للرعاف هو مسار جيني يعتمد فيه التعبير عن جين واحد على وظيفة الجين الذي يسبقه أو يتبعه في المسار.
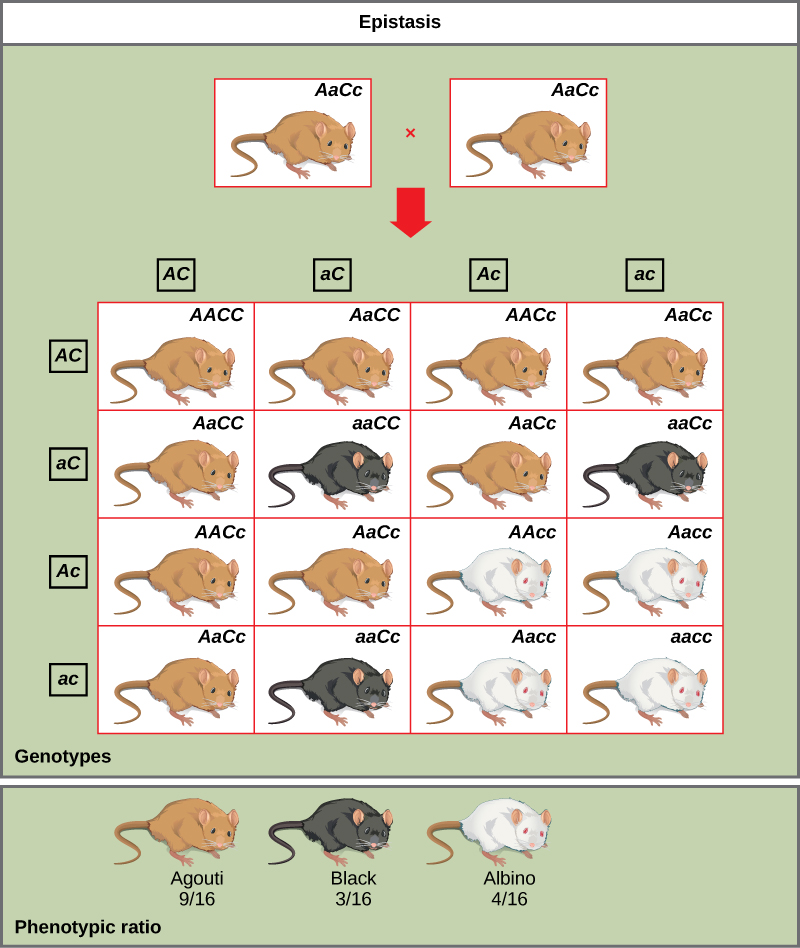
مثال على الرعاف هو التصبغ في الفئران. يسود لون المعطف من النوع البري، agouti (AA)، على الفراء ذي اللون الصلب (aa). ومع ذلك، فإن الجين المنفصل (C) ضروري لإنتاج الصباغ. الفأر الذي يحتوي على أليل متنحي في هذا المكان غير قادر على إنتاج الصبغة وهو ألبينو بغض النظر عن الأليل الموجود في الموضع A (الشكل\(\PageIndex{6}\)). لذلك، تنتج جميع الأنماط الجينية Aacc و Aacc و acc نفس النمط الظاهري للألبينو. سيؤدي التقاطع بين الزيجوت غير المتجانسة لكلا الجينين (AACc x AACC) إلى توليد ذرية بنسبة ظاهرية تبلغ 9 أغوتي: 3 اللون الصلب: 4 ألبينو (الشكل\(\PageIndex{6}\)). في هذه الحالة، يتحول الجين C إلى الجين A.
يمكن أن يحدث الرعاف أيضًا عندما يخفي الأليل السائد التعبير في جين منفصل. يتم التعبير عن لون الفاكهة في القرع الصيفي بهذه الطريقة. يؤدي التعبير المتنحي المتجانس للجين W (ww) إلى جانب التعبير المنحي المتجانس أو غير المتجانس للجين Y (YY أو Yy) إلى توليد فاكهة صفراء، والطريقة التي ينتج بها النمط الجيني الفاكهة الخضراء. ومع ذلك، في حالة وجود نسخة سائدة من جين W في الشكل المتجانس أو غير المتجانس، فإن القرع الصيفي سينتج ثمارًا بيضاء بغض النظر عن الأليلات Y. سيؤدي التقاطع بين الزيجوت البيضاء غير المتجانسة لكلا الجينين (WwYy × WyY) إلى إنتاج ذرية بنسبة ظاهرية تبلغ 12 أبيض:3 أصفر:1 أخضر.
أخيرًا، يمكن أن يكون الرعاف متبادلاً بحيث يعبر أي من الجين، عند وجوده في الشكل المهيمن (أو المتنحي)، عن نفس النمط الظاهري. في نبات محفظة الراعي (Capsella bursa-pastoris)، يتم التحكم في خاصية شكل البذور بواسطة جينين في علاقة رسطية سائدة. عندما يكون كلا الجينين A و B متنحين متماثلين (aabb)، تكون البذور بيضاوية الشكل. في حالة وجود الأليل السائد لأي من هذه الجينات، تكون النتيجة بذور مثلثة. أي أن كل نمط وراثي ممكن بخلاف aabb ينتج عنه بذور مثلثة، وسيؤدي التقاطع بين الزيجوت غير المتجانس لكلا الجينين (AABb x AABB) إلى نسل بنسبة ظاهرية تبلغ 15 مثلثًا: 1 بيضوي.
أثناء التعامل مع المشكلات الوراثية، ضع في اعتبارك أن أي خاصية منفردة تؤدي إلى نسبة نمطية تبلغ 16 هي سمة نموذجية للتفاعل بين جينين. تذكر نمط الوراثة المظهرية لصليب مندل ثنائي الهجين، والذي أخذ في الاعتبار جينين غير متفاعلين - 9:3:3:1. وبالمثل، نتوقع أن تُظهر أزواج الجينات المتفاعلة أيضًا نسبًا معبرًا عنها بـ 16 جزءًا. لاحظ أننا نفترض أن الجينات المتفاعلة غير مرتبطة؛ فهي لا تزال تنقسم بشكل مستقل إلى الأمشاج.
ملخص
افترض مندل أن الجينات (الخصائص) موروثة كأزواج من الأليلات (الصفات) التي تتصرف بنمط مهيمن ومتنحي. تنقسم الأليلات إلى أمشاج بحيث يكون من المرجح أن يتلقى كل مشيج بنفس القدر أيًا من الأليلين الموجودين في فرد ثنائي الشكل. بالإضافة إلى ذلك، يتم تصنيف الجينات في الأمشاج بشكل مستقل عن بعضها البعض. أي أنه من غير المرجح عمومًا أن تنفصل الأليلات إلى مشيج بأليل معين من جين آخر. يُظهر الصليب ثنائي الهجين تنوعًا مستقلاً عندما تكون الجينات المعنية على كروموسومات مختلفة أو بعيدة عن بعضها البعض على نفس الكروموسوم. بالنسبة لعمليات التقاطعات التي تشمل أكثر من جينين، استخدم الخط المتشعب أو طرق الاحتمالات للتنبؤ بالأنماط الجينية والأنماط الظاهرية للنسل بدلاً من مربع Punnett.
على الرغم من أن الكروموسومات تنقسم بشكل مستقل إلى الأمشاج أثناء الانقسام الاختزالي، فإن قانون مندل للتشكيلة المستقلة يشير إلى الجينات وليس الكروموسومات، وقد يحمل كروموسوم واحد أكثر من 1000 جين. عندما تكون الجينات قريبة من نفس الكروموسوم، تميل أليلاتها إلى أن تكون موروثة معًا. ينتج عن هذا نسب ذرية تنتهك قانون مندل للتشكيلة المستقلة. ومع ذلك، فإن إعادة التركيب تعمل على تبادل المواد الجينية على الكروموسومات المتماثلة بحيث يمكن إعادة تجميع الأليلات الأمومية والأبوية على نفس الكروموسوم. هذا هو السبب في أن الأليلات الموجودة على كروموسوم معين لا يتم توريثها دائمًا معًا. إعادة التركيب هي حدث عشوائي يحدث في أي مكان على الكروموسوم. لذلك، من المرجح أن تستمر الجينات المتباعدة في نفس الكروموسوم في الفرز بشكل مستقل بسبب أحداث إعادة التركيب التي حدثت في الفضاء الكروموسومي المتداخل.
سواء كانت الجينات تقوم بالفرز بشكل مستقل أم لا، فقد تتفاعل الجينات على مستوى المنتجات الجينية بحيث يؤدي التعبير عن الأليل لجين واحد إلى إخفاء أو تعديل تعبير الأليل لجين مختلف. هذا يسمى الرعاف.
مسرد المصطلحات
- منزوع الهجين
- نتيجة التقاطع بين أبوين يتكاثران حقيقيان يعبران عن سمات مختلفة لخاصيتين
- الرعاف
- التفاعل العدائي بين الجينات بحيث يخفي أحد الجينات أو يتداخل مع التعبير عن الآخر
- قانون الهيمنة
- في الزيجوت غير المتجانس، ستخفي إحدى السمات وجود سمة أخرى لنفس الخاصية
- قانون التشكيلة المستقلة
- لا تؤثر الجينات على بعضها البعض فيما يتعلق بفرز الأليلات إلى الأمشاج؛ من المرجح أن يحدث كل مزيج ممكن من الأليلات بنفس القدر
- قانون الفصل
- تنقسم عوامل الوحدة المزدوجة (أي الجينات) بالتساوي إلى الأمشاج بحيث يكون لدى النسل احتمال متساوٍ لوراثة أي مجموعة من العوامل
- الربط
- ظاهرة تكون فيها الأليلات الموجودة بالقرب من بعضها البعض على نفس الكروموسوم أكثر عرضة للتوريث معًا