
7.3: أكسدة البيروفات ودورة حمض الستريك
- Page ID
- 196294

المهارات اللازمة للتطوير
- اشرح كيف يختلف المسار الدائري، مثل دورة حمض الستريك، اختلافًا جوهريًا عن المسار الخطي، مثل تحلل السكر
- وصف كيفية تحضير البيروفات، وهو منتج تحلل السكر، للدخول في دورة حمض الستريك
إذا كان الأكسجين متاحًا، فسيمضي التنفس الهوائي إلى الأمام. في الخلايا حقيقية النواة، يتم نقل جزيئات البيروفات المنتجة في نهاية تحلل السكر إلى الميتوكوندريا، وهي مواقع التنفس الخلوي. هناك، سيتم تحويل البيروفات إلى مجموعة أسيتيل يتم التقاطها وتنشيطها بواسطة مركب حامل يسمى الإنزيم المساعد A (CoA). يسمى المركب الناتج أسيتيل CoA. CoA مصنوع من فيتامين B5 وحمض البانتوثيني. يمكن استخدام Acetyl CoA بعدة طرق من قبل الخلية، ولكن وظيفته الرئيسية هي توصيل مجموعة الأسيتيل المشتقة من البيروفات إلى المرحلة التالية من المسار في تقويض الجلوكوز.
انهيار البيروفات
لكي يدخل البيروفات، منتج تحلل السكر، المسار التالي، يجب أن يخضع لعدة تغييرات. التحويل عبارة عن عملية من ثلاث خطوات (الشكل\(\PageIndex{1}\)).
الخطوة 1. تتم إزالة مجموعة الكربوكسيل من البيروفات، مما يؤدي إلى إطلاق جزيء من ثاني أكسيد الكربون في الوسط المحيط. نتيجة هذه الخطوة هي مجموعة هيدروكسي إيثيل ثنائية الكربون مرتبطة بالإنزيم (ديهيدروجيناز البيروفات). هذا هو الأول من الكربونات الستة من جزيء الجلوكوز الأصلي الذي تتم إزالته. تستمر هذه الخطوة مرتين (تذكر: هناك جزيئان من البيروفات يتم إنتاجهما في نهاية الجليكولسيس) لكل جزيء من الجلوكوز يتم استقلابه؛ وبالتالي، ستتم إزالة اثنين من الكربونات الستة في نهاية كلتا الخطوتين.
الخطوة 2. تتأكسد مجموعة هيدروكسي إيثيل إلى مجموعة أسيتيل، ويتم التقاط الإلكترونات بواسطة NAD+، لتشكيل NADH. سيتم استخدام الإلكترونات عالية الطاقة من NADH لاحقًا لتوليد ATP.
الخطوة 3. يتم نقل مجموعة الأسيتيل المرتبطة بالإنزيم إلى CoA، مما ينتج جزيئًا من الأسيتيل CoA.
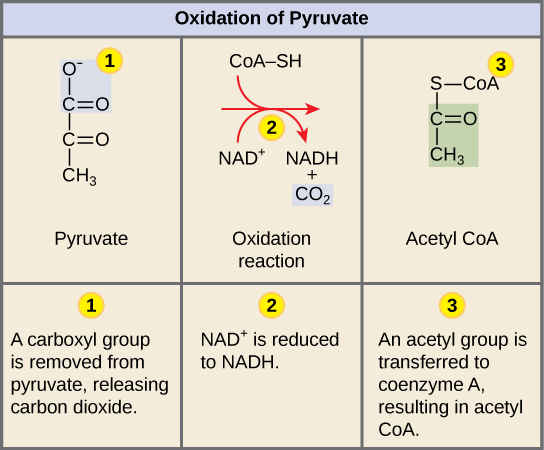
لاحظ أنه خلال المرحلة الثانية من استقلاب الجلوكوز، عندما تتم إزالة ذرة الكربون، فإنها ترتبط بذرتين من الأكسجين، مما ينتج ثاني أكسيد الكربون، وهو أحد المنتجات النهائية الرئيسية للتنفس الخلوي.
أسيتيل أوكا إلى ثاني أكسيد الكربون
في وجود الأكسجين، يقوم الأسيتيل CoA بتوصيل مجموعة الأسيتيل الخاصة به إلى جزيء رباعي الكربون، أوكسالوأسيتات، لتكوين السيترات، وهو جزيء من ستة كربون مع ثلاث مجموعات من الكربوكسيل؛ سيحصد هذا المسار ما تبقى من الطاقة القابلة للاستخراج مما بدأ كجزيء جلوكوز. يُطلق على هذا المسار الفردي أسماء مختلفة: دورة حمض الستريك (لأول وسيط يتكون - حمض الستريك، أو السيترات - عندما ينضم الأسيتات إلى أوكسكالوأسيتات)، ودورة TCA (نظرًا لأن حمض الستريك أو السيترات والإيزوسيترات عبارة عن أحماض ثلاثية الكربوكسيل)، ودورة كريبس ، على اسم هانز كريبس، الذي حدد الخطوات في المسار لأول مرة في ثلاثينيات القرن الماضي في عضلات طيران الحمام.
دورة حمض الستريك
مثل تحويل البيروفات إلى أسيتيل CoA، تحدث دورة حمض الستريك في مصفوفة الميتوكوندريا. جميع إنزيمات دورة حمض الستريك تقريبًا قابلة للذوبان، باستثناء وحيد هو إنزيم سكسينات ديهيدروجيناز، وهو جزء لا يتجزأ من الغشاء الداخلي للميتوكوندريا. على عكس تحلل السكر، فإن دورة حمض الستريك عبارة عن حلقة مغلقة: الجزء الأخير من المسار يجدد المركب المستخدم في الخطوة الأولى. الخطوات الثماني للدورة هي سلسلة من تفاعلات الأكسدة والاختزال والجفاف والترطيب ونزع الكربوكسيل التي تنتج جزيئين من ثاني أكسيد الكربون، واحد GTP/ATP، وأشكال مخفضة من NADH و FADH 2 (الشكل\(\PageIndex{2}\)). يعتبر هذا مسارًا هوائيًا لأن NADH و FADH 2 المنتجين يجب أن ينقلوا إلكتروناتهم إلى المسار التالي في النظام، والذي سيستخدم الأكسجين. في حالة عدم حدوث هذا النقل، لا تحدث خطوات الأكسدة لدورة حمض الستريك أيضًا. لاحظ أن دورة حمض الستريك تنتج القليل جدًا من ATP بشكل مباشر ولا تستهلك الأكسجين بشكل مباشر.
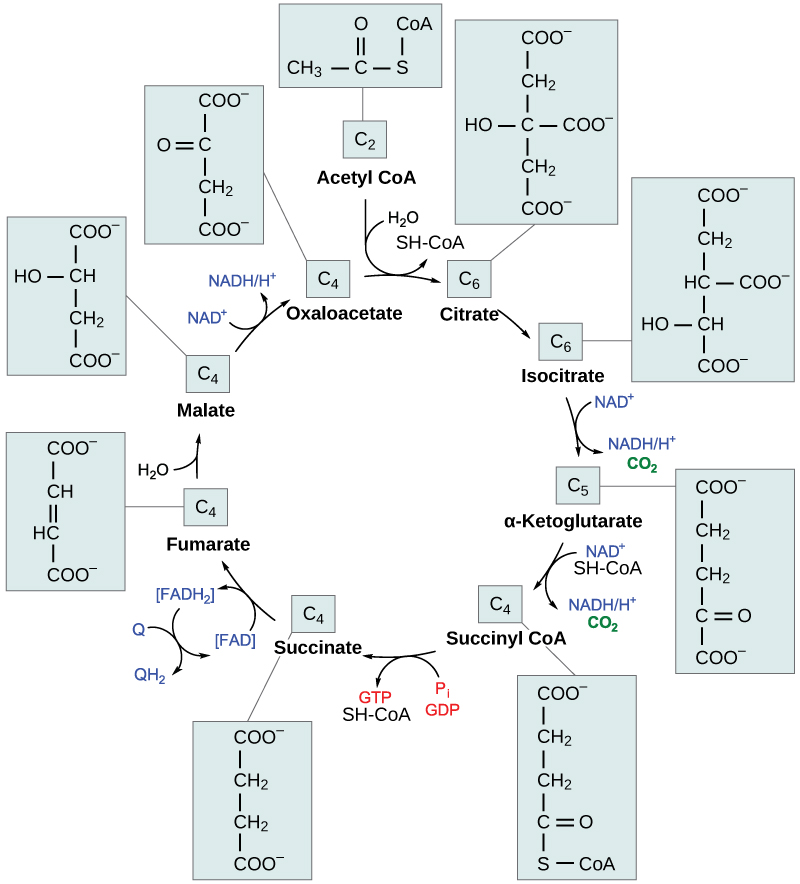
خطوات دورة حمض الستريك
الخطوة 1. قبل بدء الخطوة الأولى، تحدث مرحلة انتقالية يتم خلالها تحويل حمض البيروفيك إلى acetyl CoA. ثم تبدأ الخطوة الأولى من الدورة: هذه هي خطوة التكثيف، حيث تجمع بين مجموعة الأسيتيل ثنائي الكربون وجزيء أوكسالوأسيتات رباعي الكربون لتشكيل جزيء سداسي الكربون من السيترات. يرتبط CoA بمجموعة سلفهيدريل (-SH) وينتشر بعيدًا لينضم في النهاية مع مجموعة أسيتيل أخرى. هذه الخطوة لا رجعة فيها لأنها تتطلب مجهودًا كبيرًا. يتم التحكم في معدل هذا التفاعل من خلال ردود الفعل السلبية وكمية ATP المتاحة. في حالة زيادة مستويات ATP، ينخفض معدل هذا التفاعل. إذا كان هناك نقص في ATP، يزداد المعدل.
الخطوة 2. في الخطوة الثانية، يفقد السيترات جزيئًا مائيًا ويكسب جزيئًا آخر عندما يتم تحويل السيترات إلى أيزومر، أيزوسيترات.
الخطوة 3. في الخطوة الثالثة، يتأكسد الإيزوسيترات، وينتج جزيئًا من خمسة كربون، ألفا كيتوجلوتارات، جنبًا إلى جنب مع جزيء من ثاني أكسيد الكربون واثنين من الإلكترونات، مما يقلل NAD+ إلى NADH. يتم تنظيم هذه الخطوة أيضًا من خلال التعليقات السلبية من ATP و NADH، والتأثير الإيجابي لـ ADP.
الخطوات 3 و 4. الخطوتان الثالثة والرابعة هي خطوات الأكسدة ونزع الكربوكسيل، والتي تطلق الإلكترونات التي تقلل NAD+ إلى NADH وتطلق مجموعات الكربوكسيل التي تشكل جزيئات ثاني أكسيد الكربون. ألفا كيتوجلوتارات هي نتاج الخطوة الثالثة، ومجموعة السكسينيل هي نتاج الخطوة الرابعة. تُلزم Coa مجموعة السكسينيل بتشكيل CoA سكسينيل. يتم تنظيم الإنزيم الذي يحفز الخطوة الرابعة عن طريق تثبيط التغذية الراجعة لـ ATP و Succinyl CoA و NADH.
الخطوة 5. في الخطوة الخامسة، يتم استبدال مجموعة الفوسفات بالإنزيم المساعد A، ويتم تكوين رابطة عالية الطاقة. تُستخدم هذه الطاقة في الفسفرة على مستوى الركيزة (أثناء تحويل مجموعة السكسينيل إلى سكسينات) لتشكيل إما جوانين ثلاثي الفوسفات (GTP) أو ATP. هناك نوعان من الإنزيم، يُطلق عليهما الإيزوإنزيمات، لهذه الخطوة، اعتمادًا على نوع الأنسجة الحيوانية التي توجد فيها. يوجد أحد الأشكال في الأنسجة التي تستخدم كميات كبيرة من ATP، مثل القلب والعضلات الهيكلية. ينتج هذا النموذج ATP. يوجد الشكل الثاني من الإنزيم في الأنسجة التي تحتوي على عدد كبير من المسارات الابتنائية، مثل الكبد. ينتج هذا النموذج GTP. GTP يعادل بقوة ATP؛ ومع ذلك، فإن استخدامه أكثر تقييدًا. على وجه الخصوص، يستخدم تخليق البروتين في المقام الأول GTP.
الخطوة 6. الخطوة السادسة هي عملية الجفاف التي تحول السكسينات إلى فومارات. يتم نقل ذرتين من الهيدروجين إلى FAD لإنتاج FADH 2. الطاقة الموجودة في إلكترونات هذه الذرات غير كافية لتقليل NAD + ولكنها كافية لتقليل FAD. على عكس NADH، يظل هذا الناقل مرتبطًا بالإنزيم وينقل الإلكترونات إلى سلسلة نقل الإلكترون مباشرة. أصبحت هذه العملية ممكنة من خلال توطين الإنزيم الذي يحفز هذه الخطوة داخل الغشاء الداخلي للميتوكوندريا.
الخطوة 7. يتم إضافة الماء إلى الفومارات خلال الخطوة السابعة، ويتم إنتاج المالات. تعمل الخطوة الأخيرة في دورة حمض الستريك على تجديد أوكسالوأسيتات عن طريق أكسدة المالات. يتم إنتاج جزيء آخر من NADH في هذه العملية.

منتجات دورة حمض الستريك
تدخل ذرتان من الكربون في دورة حمض الستريك من كل مجموعة أسيتيل، مما يمثل أربعة من الكربونات الستة لجزيء جلوكوز واحد. يتم إطلاق جزيئين من ثاني أكسيد الكربون في كل دورة؛ ومع ذلك، لا تحتوي هذه الجزيئات بالضرورة على أحدث ذرات الكربون المضافة. سيتم إطلاق ذرتي الكربون الأسيتيل في نهاية المطاف في دورات لاحقة من الدورة؛ وبالتالي، يتم دمج جميع ذرات الكربون الست من جزيء الجلوكوز الأصلي في نهاية المطاف في ثاني أكسيد الكربون. كل دورة من الدورة تشكل ثلاثة جزيئات NADH وجزيء واحد FADH 2. سوف تتصل هذه الناقلات بالجزء الأخير من التنفس الهوائي لإنتاج جزيئات ATP. يتم أيضًا إجراء GTP أو ATP واحد في كل دورة. يمكن استخدام العديد من المركبات الوسيطة في دورة حمض الستريك في تصنيع الأحماض الأمينية غير الأساسية؛ لذلك، تكون الدورة أمفيبوليك (تقويضية وبنائية على حد سواء).
ملخص
في وجود الأكسجين، يتحول البيروفات إلى مجموعة أسيتيل مرتبطة بجزيء حامل من الإنزيم المساعد A. يمكن أن يدخل الأسيتيل CoA الناتج عدة مسارات، ولكن في أغلب الأحيان، يتم تسليم مجموعة الأسيتيل إلى دورة حمض الستريك لمزيد من الهدم. أثناء تحويل البيروفات إلى مجموعة الأسيتيل، تتم إزالة جزيء من ثاني أكسيد الكربون واثنين من الإلكترونات عالية الطاقة. يمثل ثاني أكسيد الكربون اثنين (تحويل جزيئين من البيروفات) من الكربونات الستة لجزيء الجلوكوز الأصلي. يتم التقاط الإلكترونات بواسطة NAD+، ويحمل NADH الإلكترونات إلى مسار لاحق لإنتاج ATP. في هذه المرحلة، تم أكسدة جزيء الجلوكوز الذي دخل التنفس الخلوي في الأصل تمامًا. تم نقل الطاقة الكامنة الكيميائية المخزنة داخل جزيء الجلوكوز إلى ناقلات الإلكترون أو تم استخدامها لتجميع عدد قليل من ATPs.
دورة حمض الستريك عبارة عن سلسلة من تفاعلات الأكسدة والاختزال التي تزيل الإلكترونات عالية الطاقة وثاني أكسيد الكربون. يتم استخدام الإلكترونات المخزنة مؤقتًا في جزيئات NADH و FADH 2 لتوليد ATP في مسار لاحق. يتم إنتاج جزيء واحد من GTP أو ATP عن طريق الفسفرة على مستوى الركيزة في كل دورة من الدورة. لا توجد مقارنة بين المسار الدوري والمسار الخطي.
مسرد المصطلحات
- أسيتيل أوكا
- مزيج من مجموعة أسيتيل مشتقة من حمض البيروفيك والإنزيم المساعد A، وهو مصنوع من حمض البانتوثانيك (فيتامين مجموعة B)
- دورة حمض الستريك
- (أيضًا دورة كريبس) سلسلة التفاعلات الكيميائية المحفزة بالإنزيم ذات الأهمية المركزية في جميع الخلايا الحية
- دورة كريبس
- (أيضًا، دورة حمض الستريك) اسم بديل لدورة حمض الستريك، سمي على اسم هانز كريبس الذي حدد لأول مرة خطوات المسار في الثلاثينيات في عضلات طيران الحمام؛ انظر دورة حمض الستريك
- دورة TCA
- (أيضًا، دورة حمض الستريك) اسم بديل لدورة حمض الستريك، سمي على اسم المجموعة لحمض الستريك، حمض التريكاربوكسيليك (TCA)؛ انظر دورة حمض الستريك