
15.6: מיטוזיס
- Page ID
- 208500

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מיטוזיס מורכב מפרופאזה, מטאפאזה, אנפאזה וטלופאז, עם פעילויות סלולריות מובהקות המאפיינות כל שלב. זה משלים את שכפול הגרעין, ואחריו ציטוקינזיס, בו התא מתחלק לייצור שני תאי בת.
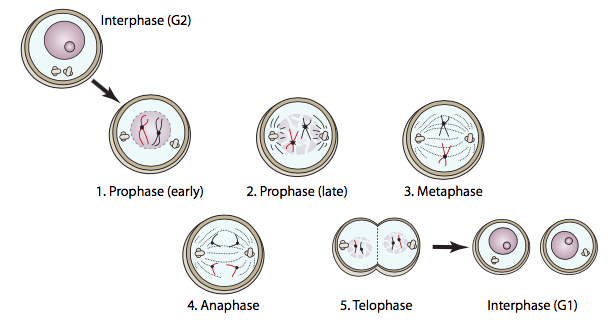
Prophase הוא הכנת כל רכיב לריקוד סלולרי מורכב זה. ה-DNA מתעבה (הוא עטוף סביב עצמו בחוזקה כדי להפוך אותו לאריזה קטנה וחזקה יותר) כך שהוא פחות רגיש לשבירה במהלך התנועה על פני התא. בכך, רוב ה- DNA הופך ללא פעיל מבחינה תעתיק. גופי הגולגי והרטיקולום האנדופלזמי מתחילים להתפרק לשלפוחיות קרומיות שניתן לפזר ביתר קלות ובאופן שווה על פני התא כך ששני תאי הבת יקבלו בערך אותו דבר. הצנטרוזומים (בתאי בעלי חיים) נעים ממיקומם המקורי ליד הגרעין לכיוון צדדים מנוגדים של התא, כדי לבסס את הקטבים של הציר המיטוטי.
MPF מזרחן חלבונים מוטוריים של מיקרו-צינוריות וחלבונים הקשורים למיקרו-צינוריות (MAPs) כדי לשנות את הדינמיקה הרגילה של המיקרוטובוליות ולאפשר להתרחש ארגון מחדש מסיבי לציר מיטוטי. לדוגמה, יעד אחד של MPF הוא PRC1, חלבון צרור שאינו מופעל על ידי זרחון, ובכך מאפשר למיקרוטובולים בודדים לעבור למיקומים חדשים בקלות רבה יותר מאשר צרור גדול יכול. השפעות אחרות הן השבתה של MAPs מייצבים, מה שמוביל ללקות רבה יותר של מיקרו-צינורות עקב שכיחות מוגברת של קטסטרופה. יעדי החלבון המוטורי של MPF נמצאים במשפחת הקינזינים והזרחון נחוץ כדי שחלקם ייקשרו לציר המיטוטי.
פרומטאפזה נחשבת לעיתים לשלב נפרד אך מכונה גם פרופאזה מאוחרת, והיא מוגדרת בעיקר על ידי פירוק המעטפת הגרעינית. תהליך זה נגרם על ידי זרחון MPF של הלמינים הגרעיניים. המעוטרים במטענים שליליים מהפוספטים, הלמינים מסרבים להתחבר זה לזה עוד, מה שמוביל להתמוטטות הלמינה הגרעינית. כאשר הלמינים מתנתקים, המעטפת הגרעינית נשארת קשורה אליהם, ושברים. הפיצול הגרעיני הזה חייב לקרות כדי שהציר המיטוטי יוכל להגיע פנימה ולהיצמד לכרומוזומים.
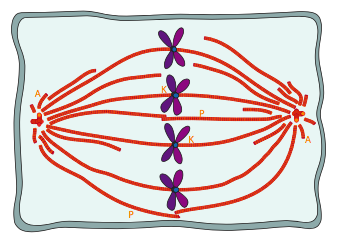
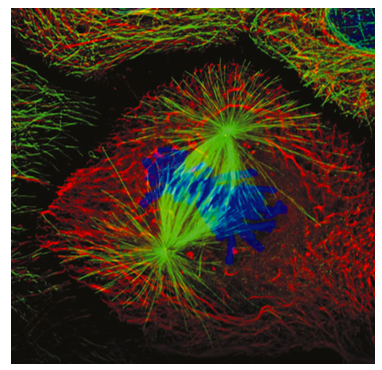
חלק מהמיקרוטובולים של הציר המיטוטי מתחברים לכרומוזומים דרך חלבוני הקינטוכור, המקשרים את המיקרוטובולים של הציר לאזור הצנטרומרים של כל כרומוזום. אלה ידועים בשם מיקרו-צינורות קינטוכור (איור). \(\PageIndex{8}\) ישנם שני סוגים נוספים של מיקרוטובולים בציר המיטוטי (איור\(\PageIndex{7}\)): המיקרוטובולים הקוטביים המגיעים על פני התא ומקיימים אינטראקציה זה עם זה כדי לסייע בשמירה על הפרדת הצנטרוזומים והגדרת האורך הכולל של הציר, והמיקרוטובולים של אסטר שהם בדרך כלל קצרים, מקרינים החוצה ומייצבים את הצנטרוזום. זכור כי ה- DNA שוכפל מוקדם יותר בשלב S, ולכן כרומטידות אחיות עדיין מחוברות חלקית. מבחינה ויזואלית, אזור הצנטרומרים נראה צר יותר או דחוס יותר משאר הכרומוזום, ובדרך כלל שוכן ליד האמצע. הצנטרומר מכיל רצפים חוזרים ונשנים המעורבים בקשירה והרכבה של קינטוכור.
בפרימטים, המוטיב החוזר ידוע בשם DNA לווין אלפא, אשר עשוי ממקרים מרובים של חזרות טנדם של רצף ליבה ~170bp על פני טווח DNA צנטרומרי לאורך מגה-בסיס. חזרות דומות נמצאות גם בבעלי חוליות שונים אחרים. באיקריוטים אחרים, הגודל והרצף עשויים להשתנות; לדוגמה, חזרות קצרות בהרבה של ~5bp נמצאות ב-DNA צנטרומרי בגודל 200-600kb בכרומוזומי תסיסנית, ו-S. pombe יש DNA צנטרומרי הרבה מתחת ל-10kb.
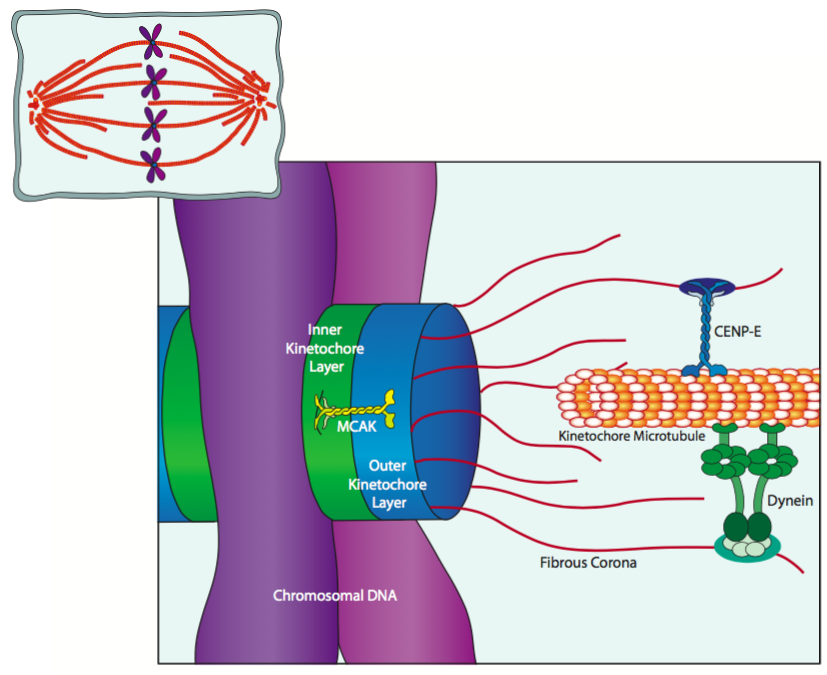
הקינטוכורים הנצמדים ל-DNA הצנטרומר הם מבני חלבון טרילמינריים המורכבים משכבה פנימית, שכבה חיצונית וקורונה סיבית. המיקרוטובולים הקינטוכורים של הציר המיטוטי מחוברים בעיקר לקורונה הסיבית. כפי שמתואר באיור, הוא מחובר דרך CENP-E, קינסין וחלבונים מוטוריים דינין הנקשרים לאורך חבית המיקרוטובולה. למעשה, לפעמים המגע הראשון בין כרומוזום (דרך הקינטוכור) למיקרו-צינורית ציר נמצא אי שם באמצע המיקרוטובולה, ושילוב של דינמיקה של מיקרו-צינוריות ופעילות חלבון מוטורי מעביר את הכרומוזום לקצה הדיסטלי של המיקרוטובולה. זה מתאפשר על ידי MCAK (קינסין הקשור לצנטרומר מיטוטי), הקשור לחלבוני הליבה של הקינטוכור וממלא תפקיד בדפולימריזציה של מיקרוטובולים ליד הקצה (+).
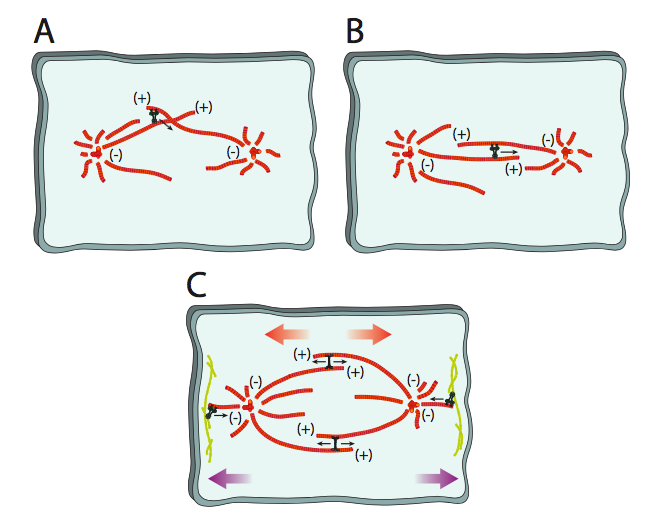
המעבר מהציטו-שלד המיקרוטובולי הבין-פאזי לציר מיטוטי דורש מספר מנועים מולקולריים כדי להזיז את הצנטרוזומים, ליישר את המיקרוטובולים ולהרחיב את הציר. אלה מתוארים באיור\(\PageIndex{9}\). בתחילה, כאשר הצנטרוזומים המשוכפלים מתרחקים זה מזה יחד עם חלק מהמיקרוטובולים הציטו-שלד, המיקרוטובולים יתקשרו בזוויות שונות. מכיוון שהמיקרוטובולים הקוטביים שעוזרים להרחיב או לשמור על רוחב הציר חייבים לפעול במקביל, דינינים ציטופלסמיים נקשרים למיקרוטובולים הקוטביים בסופו של דבר ועל ידי הזזת אחד לאורך השני, מביאים אותם במקביל (9a). ברגע שהם נמצאים במצב זה, BiMC וקינסינים אחרים משתלטים כמנועים העיקריים לאורך מיקרו-צינורות קוטביים. הם יוצרים כוח דחיפה כלפי חוץ על ידי אחיזה במיקרו-צינורית הפונה לכיוון אחד, ונהיגה לאורך MT מקביל הפונה לכיוון ההפוך לכיוון הקצה (+) (9b). לבסוף, דינינים ציטוזוליים המחוברים לציטוס שלד קליפת המוח מושכים את המיקרוטובולים האסטרליים, מה שמושך את קצות הציר רחוק יותר מהמרכז (9c).
למעשה, נראה שיש שני מנגנונים בעבודה: מערכת bub1/bub2 פועלת במסלול חישת המתח, בעוד שחלבון מטאפאזה אחר, mad2 נראה חשוב בהשעיית המיטוזה עם ניתוק הקינטוכור עם המיקרוטובולה של הציר.
כאשר המעטפת הגרעינית מתפרקת, המיקרוטובולים של הציר המיטוטי עוברים אי יציבות דינמית מוגברת, עוברים בין תקופות של קפיצות גדילה (פילמור) וקיצור מהיר (פירוק קטסטרופלי), ומחפשים כרומוזומים להתחבר אליהם. ברגע שהמיקרוטובולים הקינטוכורים מתחברים לכרומוזומים, הדינמיקה המיקרוטובולית משתנה. המיקרוטובולה תעבור בעיקר קיצור אם היא נמצאת מעבר למרכז הציר ובעיקר מתארכת אם היא קצרה מהמרכז. מכיוון שבסופו של דבר כל קבוצה של כרומטידות אחיות מחוברת למיקרוטובולים בשני הקינטוכורים, כל כרומטיד מחובר לקיצור אחד ולמיקרוטובולה אחת מתארכת. כאשר הכרומוזומים מתקרבים למרכז הציר המיטוטי, קצב הקיצור/התארכות המיקרוטובוליות מאט. הכרומטידות האחיות נדחפות ונמשכות על ידי המיקרו-צינורות של הציר עד שכולן מסודרות לאורך קו האמצע של הציר המיטוטי, שברוב המקרים (אך לא בכולם) הוא גם קו האמצע של התא. ברגע שכולם מסודרים בשורה, התא נחשב כמי שהגיע למטאפאזה. בניגוד לשלבים האחרים, מטאפאזה היא שלב סטטי יחסית - זהו מחסום ליישור הכרומוזומים.
הכרומוזומים חייבים להיות מיושרים כראוי כדי להבטיח ששני תאי הבת יקבלו את ההשלמה הנכונה של הכרומוזומים. כיצד התא יודע מתי הכרומוזומים הגיעו למרכז הציר? ניסוי פשוט באלגנטיות הוכיח שהמנגנון הכללי הוא בדיקת מתח - אם שני המיקרוטובולים המתחברים לזוג הכרומטידות האחיות מכל צד הם באותו אורך, עליהם להפעיל מתח שווה על הכרומוזומים. אם חיבור המיקרוטובול-קינטוכור מנותק במטאפאזה, התא יימנע מלהתקדם (Nicklas, R.B., et al, J Cell Biol. 130: 929-39, 1995). עם זאת, אם מופעל מתח שווה ערך על ידי משיכה בכרומוזום בעזרת מיקרו-חוט זכוכית, התקדמות המיטוזה משוחזרת!
בנוסף לבדיקת המתח, יש תנאי נוסף שיש לעמוד בו להמשך המיטוזה: יש להשבית את ה- MPF. כפי שתואר קודם לכן, MPF מוביל בחלקו לאי-אקטיבציה משלו על ידי הפעלת המתחם המקדם אנפאזה (APC), אשר פוליוביקוויטינט את הציקלין, מה שמוביל להרס שלו ובכך להשבתת MPF-Cdk. APC מתייג גם את Securin להשמדה. Securin הוא חלבון הקושר ומעכב את האנזים הפרוטאוליטי, separase, אשר הפעלתו נדרשת כדי לאפשר לכרומטידות האחות להיפרד, אשר בתורו, הכרחי כדי להמשיך באנפאזה.
חסימת מצבים פתולוגיים, אם ורק אם הכרומוזומים כולם מסתדרים בצלחת המטאפאזה, התא ימשיך לשלב הבא של המיטוזה: אנאפאזה. הכרומטידות האחיות נפרדות ונמשכות לעבר קטבים מנוגדים של הציר המיטוטי. קצת סוטה, גם כשהכרומוזומים נעים לכיוון קטבי הציר, הקטבים עצמם נעים מעט החוצה. הפרדת הכרומטידות האחיות מחייבת ניתוק של ה"דבק "המולקולרי המחזיק אותם יחד: חלבוני הקוהזין. הקוהזינים נקשרים לשתי מולקולות ה-DNA ומחזיקים אותן יחד זמן קצר לאחר השכפול בחזרה בשלב S. ככל שמתקרב אנפאזה, מופעל הפרדת האנזים, אשר לאחר מכן חותך את מולקולות הקוהזין. ברגע שכל מולקולות הקוהזין נחתכות, ניתן סוף סוף להפריד את הכרומטידות האחיות. הסרת הקוהזינים נמשכת בערך פנימה מהנקודות הדיסטליות של הכרומוזומים לצנטרומר, שהוא בדרך כלל אזור ההתקשרות האחרון.
קוהזין הוא מולטימר של ארבע יחידות משנה, Scc1, Scc3, Smc1 ו- Smc3 בשמרים. חלבון נוסף נצפה גם ב- Xenopus. חלבון SCC1 מבוקע על ידי הפרדה בשמרים, אך במטאזואנים, SCC1 עשוי להיות מוסר מכרומוזומים גם בשיטה אחרת. הוא זרחני, מה שמפחית את הזיקה שלו ל-DNA, ועלול לחשוף אתר להידרוליזה מזורזת הפרדה.
Separase גם מקדם אנאפאזה על ידי הפעלת Cdc14, פוספטאז הדרוש לדה-פוספורילציה של מצעי cdk שעברו זרחון על ידי מתחמי הציקלין-cdk של מיטוזה מוקדמת. בנוסף, Cdc14 נדרש גם לציטוקינזיס בשמרים S. cerevisiae ונמטודות C. elegans.
למעשה ניתן לחלק את האנאפאזה לשני שלבים, המכונים לפעמים מוקדם ומאוחר או A ו- B. בהתחלה, המיקרוטובולים הקינטוכורים מתקצרים משני הקצוות, ומנועים ממשפחת הקינזינים מושכים את המיקרוטובולים בחזרה לכיוון קטבי הציר. עם תחילת אנאפאזה מאוחרת, המיקרוטובולים הקוטביים מתארכים, וכוח נוסף המפריד בין כרומטידים מופעל על ידי חלבונים מוטוריים ממשפחת הקינזינים [קינזין-5] הדוחפים את המיקרוטובולים הקוטביים זה מול זה כדי להגביר את ההפרדה בין הקטבים. מנועים ממשפחת Dynein עוזרים לכוון תנועה של הקטבים גם כן, דרך הצמדתם למיקרו-צינורות האסטר ולשלד הקורטיקלי (היקפי).
כאשר שתי קבוצות הכרומוזומים מגיעות לקטבים שלהן, מתחיל הטלופאז. מבחינה טכנית, הוא הצטבר לאט לאט מאז האנאפאזה: כאשר MPF הושבת על ידי APC, הסתיימה יכולתו לזרחן למינים גרעיניים. פוספטאזות חלבון בתא מסירות את קבוצות הפוספט, ומאפשרות ללמינים לקיים אינטראקציה שוב זה עם זה, ועל ידי טלופאז הם משחזרים את הלמינה הגרעינית ואת המעטפת הגרעינית. מכיוון שהלמינים וחלבוני הממברנה הגרעינית האחרים מקיימים אינטראקציה גם עם DNA, שברי הממברנה הגרעינית התפזרו בחזרה בפרופזה מאוחרת מתלכדים כעת סביב כל קבוצת DNA ליצירת המעטפות הגרעיניות החדשות. שאר האברונים הממברניים המקוטעים (ER, Golgi) מתחילים להיווצר מחדש. בסוף הטלופאז, המוצר הוא תא גדול יחיד עם שני גרעינים שלמים בצדדים מנוגדים. השלב הבא והאחרון, ציטוקינזיס, מפצל את התא לשני תאי בת נפרדים ועצמאיים. בתאי בעלי חיים, ציטוקינזיס דומה להידוק שרוך באמצע התא, מושך את "המותניים" פנימה עד שכל הקצוות נפגשים, ונוצרים שני תאים נפרדים. טבעת התכווצות זו מורכבת מיחידות משנה של אקטין (מבני) ומיוסין (מניע). חלבונים אלה, המשתמשים ב- ATP לאנרגיה, מחגרים את עצמם קרוב יותר ויותר זה לזה בדומה ל"שבץ הכוח "של אקטין-מיוסין המתואר לסרקומרים של תאי שריר, העשוי בעיקר מאקטין ומיוסין. מנגנון זה הוא אוניברסלי לתאי בעלי חיים, אך מיקום הטבעת אינו תמיד במרכז התא. הטבעת עולה בקנה אחד עם מרכז התא, אך למעשה ממוקמת על ידי צלחת המטאפאזה (כלומר מרכז הציר המיטוטי). הדוגמה הברורה ביותר לצלחת מטאפאזה שאינה עולה בקנה אחד עם מרכז התא נמצאת ביצירת תאי ביצה. מכיוון שמטרתו של תא ביצית היא לספק את כל החומר הדרוש ליצירת אורגניזם חדש בר-קיימא עם ההפריה (הזרע תורם ביומסה זניחה מעבר לחומר הגנטי), הוא מתחלק בצורה לא סימטרית, כאשר הציר המיטוטי ממוקם הרחק לצד אחד של התא (איור). \(\PageIndex{11}\)
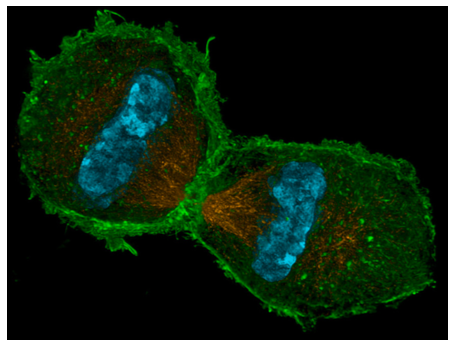
כאשר מתרחשת ציטוקינזיס, תא בת אחד, הביצית המשוערת, גדול מאוד, ואילו לתא השני, הנקרא גוף קוטבי, יש חומר ציטופלזמי מינימלי המקיף את הגרעין. הטבעת המתכווצת פועלת בתאי בעלי חיים מכיוון שקרום התא גמיש. בתאי צמחים, קרום התא מחובר היטב לדופן תא נוקשה, ולכן לא ניתן למשוך אותו פנימה. אז, תא הצמח בונה בצורה גאונית קיר באמצע התא באמצעות שלפוחיות מיוחדות שמקורן בחלק מהגולגי, ואשר מכילות את החומרים הדרושים ליצירת דופן תא. השלפוחיות נעות לאורך הפרגמופלסט, מבנה הבנוי מהמיקרוטובולים של הציר המיטוטי, וככל שהשלפוחיות מסתדרות לאורך אמצע התא, הן מתחילות להתמזג ליצירת שלפוחיות גדולות יותר ואז שלפוחית גדולה דמוית דיסק, צלחת התא. בסופו של דבר הם מגיעים לקרום התא עצמו, והתמזגות עם זה מובילה להיווצרות דופן תא חדשה, ושני תאים שלמים ועצמאיים.
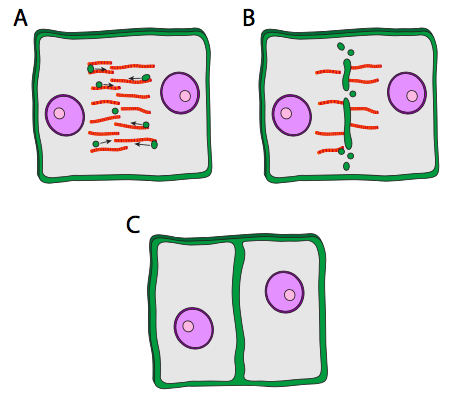
תוכן השלפוחיות הנוסעות לאורך הפרגמופלסט אינו מתואר היטב. Callose, פוליסכריד גלוקוז עם קישורי β1-3 ידוע כקיים בצלחת התא המתפתחת, אך לא נמצא בגולגי או בשלפוחיות. מעניין שברגע שצלחת התא התמזגה לחלוטין עם דפנות התא הקיימות, הקאלוזה נעלמת בהדרגה. הוא חשב שאותה מערכת אנזימים שסינתזה קאלוזה עשויה לעבור לסינתזה של תאית עם התבגרות צלחת התא.