
10.7: תרגום אוקריוטי
- Page ID
- 208437
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)תרגום אוקריוטי, כמו בתעתיק, דומה באופן מספק (מנקודת מבט של סטודנט הלומד, או משימור אבולוציוני) למקרה הפרוקריוטי. תהליך ההתחלה מעט יותר מסובך, אך תהליכי ההתארכות והסיום זהים, אך עם הומולוגים אוקריוטיים של גורמי ההתארכות והשחרור המתאימים.
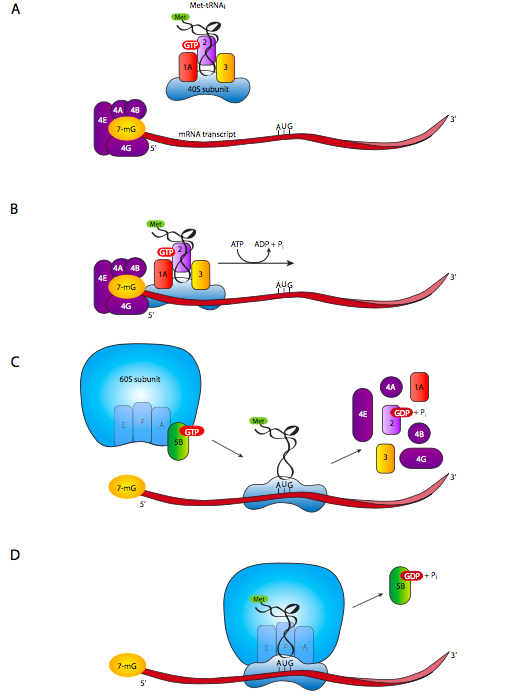
עבור אוקריוטים, כל mRNA מקודד גן אחד ויחיד (בניגוד לתמלילים מרובי גנים כגון אופרונים), כך שאין הרבה שאלה איזה AUG הוא קודון התחלה, ואילו הם רק מתיונינים רגילים. לכן, אין דרישה לרצף Shine-Dalgarno באיקריוטים. יחידת המשנה הריבוזומלית הקטנה, המלווה בגורמי התחלה אוקריוטיים eIF-3, eIF-2 ו- Met-tRNA i, המכונה יחד הקומפלקס הטרינרי, נקשרת ל- eIF-1a. בינתיים, eIF-4a, -4B, -4E ו--4G נקשרים למכסה 5' (7-מתיגואנוסין) של ה-mRNA (ז '7A). קומפלקס יחידת המשנה הקטנה וקומפלקס קושר הכובע eIF4/ mRNA מקיימים אינטראקציה ויוצרים את קומפלקס 43S, אשר לאחר מכן מתחיל לסרוק את ה-mRNA מ-5' עד 3' ומחפש את ה-AUG הראשון.
בדרך כלל, אך לא תמיד, ה- AUG הראשון הוא קודון ההתחלה לגנים אוקריוטיים. עם זאת, ההקשר של ה-AUG חשוב, והוא קודון התחלה חזק הרבה יותר (כלומר מוכר ומשומש בתדירות גבוהה יותר) אם יש שארית פורין (A או G) ב--3 ו-G ב-+4. ראו קוזאק, מ ', ביוכימיה 76: 815-821, 1994.
לאחר שמתחם הסריקה 43S מצא את קודון ההתחלה, גורמי ההתחלה יורדים, ותת-היחידה הריבוזומלית הגדולה מגיעה. יחידת המשנה הריבוזומלית הגדולה קשרה את eIF-6, מה שמונע ממנה להתחבר מחדש עם יחידות משנה קטנות, והסרתו נדרשת תחילה. גורם נוסף, eIF-5 נכנס למקום במהלך תהליך הצימוד בין יחידות המשנה הריבוזומליות הגדולות והקטנות, ונדרשת הידרוליזה של GTP מחובר eIF-5 להשלמת העגינה של יחידות המשנה ויצירת ריבוזום פונקציונלי שלם על ה- mRNA.
התארכות זהה מבחינה תפקודית כמו בפרוקריוטים פרט לכך שהפונקציות של EF-Tu מטופלות על ידי EF-1a, גם עם הידרוליזה של GTP. EF-2 הוא האנלוג האוקריוטי של EF-G, ומשתמש בהידרוליזה של GTP לצורך טרנסלוקציה של הריבוזום. סיום משתמש בהומולוגים אוקריוטיים של גורמי השחרור, אם כי eRF-1 תופס את מקומם של RF-1 הפרוקריוטי ו- RF-2.
למרות שפוליריבוזומים (aka polysomes) יכולים להיווצר הן על mRNA פרוקריוטיים והן על אוקריוטיים, לפוליזומים אוקריוטיים יש טוויסט נוסף. מבחינה טכנית, פוליזום הוא פשוט mRNA עם ריבוזומים מרובים המתרגמים אותו בו זמנית, אך באאוקריוטים, לפוליזום יש גם מורפולוגיה ייחודית מכיוון שהוא משתמש ב- PABPI, או חלבון מחייב פולי-A. חלבון זה לא רק נקשר לזנב 3' poly-A של mRNA, הוא גם מקיים אינטראקציה עם גורמי ההתחלה eIF-4, ובכך מעביר את ה- mRNA לצורה מעגלית. בדרך זו, ברגע שהריבוזום מגיע לקצה הגן ומשתחרר מה-mRNA, זה פיזית קרוב לתחילת ה-mRNA להתחיל לתרגם שוב.