
8.1: מבוא לתמלול
- Page ID
- 208468

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)כדי להשיג את המידע הגנטי בצורה הניתנת לקריאה בקלות ולאחר מכן משמשת לסינתזה של חלבונים מתפקדים, תחילה יש לתמלל את ה-DNA ל-RNA (חומצה ריבונוקלאית). כפי שראינו בפרק 1, RNA דומה מאוד ל- DNA, תוך שימוש בכמה מאותם בסיסים חנקניים (אדנין, גואנין, ציטוסין) וכן באחד ייחודי ל- RNA, אורציל. שימו לב שאורציל דומה מאוד לתימין (פרק 7, איור 1), במיוחד במיקום ובמרווח של האטומים הקשורים למימן. מכיוון שהאינטראקציה של קישור מימן של בסיסים אלה (כלומר זיווג בסיסים של גואנין לציטוסין, אדנין לתימין/אורציל) היא זו המהווה בסיס להעברת מידע מה- DNA המקורי ל- DNA של תאי בת, הגיוני לצפות שאותו סוג של מנגנון זיווג בסיסים משמש להעברת המידע ממצב אחסון בחומצת הגרעין הדו-גדילית (DNA) למצב שימושי/שמיש יותר בצורה של יחיד חומצת גרעין גדילית (RNA).
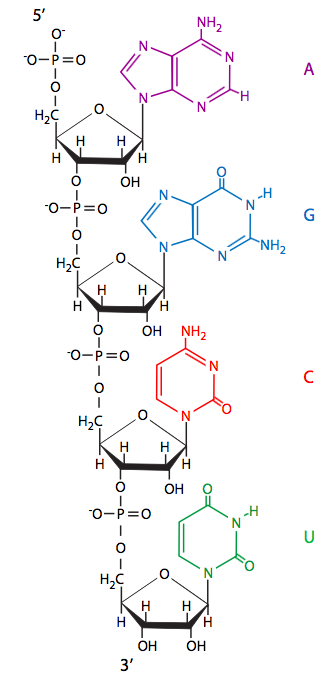
בניגוד לתפקידו הסלולרי כנשא חולף וחד פעמי של מידע גנטי, RNA נחשב כמולקולה העיקרית האחראית לאפשר חיים על פני כדור הארץ. זה זמן רב הונח כי הוא שימש תפקידים כפולים הן כמאגר של מידע גנטי והן כאנזים ראשוני לפעול על פי מידע זה. למרבה הצער, כימאים פרה-ביוטיים נחסמו במשך עשרות שנים בהמצאת מסלול סינתטי סביר שבאמצעותו RNA יכול לנבוע מהמולקולות הפשוטות של "המרק" הקדמוני של כדור הארץ. הבעיה המרכזית הייתה שניתן לסנתז ריבוז, אם כי לא ביעילות במיוחד, וניתן לסנתז בסיסים, אך לא הייתה דרך לחבר אותם זה לזה. הכימיה לא תאפשר תגובת עיבוי בין הבסיסים לסוכרים. בשנת 2009, על ידי השארת הרעיון המקובל כי ריבונוקלאוטידים בוודאי סונתזו מריבוז ופורינים/פירימידינים, פאונר, גרלנד וסאת'רלנד (טבע 459:239-242, 2009) הראו שלמעשה ניתן לסנתז ריבונוקלאוטידים מהתנאים הכימיים של כדור הארץ שזה עתה נוצר. במקום לנסות ליצור כל "חלק" ולחבר אותם יחד, Powner et al סינתזו מולקולה שהכילה חלקים ממה שבסופו של דבר יהיה גם הריבוז וגם פירימידין, 2-aminooxazole. באמצעות סדרה של תגובות שהשתמשו בפוספט כזרז וכנבלות, שכולן היו סבירות בבירור במודל הנוכחי של כדור הארץ הקדמוני, נוצרו הן ריבוציטידין והן ריבורידין. כמובן שזו רק ההתחלה, מכיוון שהדבר אינו משתרע ישירות להיווצרות נוקליאוטידים פורין, אך זהו צעד משמעותי מאוד בכימיה הפרה-ביוטית, ודוגמא מצוינת לסגולות של "לצאת מחוץ לקופסה" לפעמים.
תהליך העתקת ה- DNA ל- RNA נקרא שעתוק. הן בפרוקריוטים והן באיקריוטים, שעתוק דורש מרכיבי בקרה מסוימים (רצפים של נוקלאוטידים בתוך ה-DNA) כדי להתקדם כראוי. אלמנטים אלה הם מקדם, אתר התחלה ואתר עצירה. הצורך בנקודה מוכרת להתחיל ונקודה לסיום התהליך ברור למדי. היזם שונה במקצת. האמרגן שולט בתדירות התעתיק. אם אתה מדמיין את הצרכים של תא בכל זמן נתון, ברור שלא כל מוצרי הגנים נחוצים באותה כמות בו זמנית. חייבת להיות דרך לשלוט מתי או אם מתרחש שעתוק ובאיזו מהירות.
גרסת העצמות החשופות של התהליך הולכת בערך כך: (1) חלבוני עגינה מיוחדים מזהים את רצף האמרגן ונקשרים אליו, פותחים חלק קטן סביב אתר "ההתחלה"; (2) פולימראז RNA נקשר לאותם חלבונים מיוחדים ולמעט ה-DNA החד-גדילי שנפתח זה עתה; (3) אנזים הליקאז (חלק מפולימראז או מחובר אליו) פותח את רוכסן ה-DNA; (4) ה-RNA פולימראז עוקב אחרי ההליקאז, "קורא" את רצף ה-DNA, לוקח ריבונוקלאוטידים מהסביבה, התאמתם לתבנית ה-DNA, ואם הם להתאים, להוסיף אותם לשרשרת הריבונוקלאוטיד או ה- RNA הקודמת. זה נמשך עד שהפולימראז מגיע לאתר העצירה, ובשלב זה הוא מתנתק מה-DNA של התבנית, ומשחרר גם את עותק ה-RNA החדש שנוצר של אותו DNA. כמובן שאם זה כל מה שיש בזה, לא היו כתבי עת שלמים המוקדשים לחקר RNA, התעתיק שלו והשליטה בתעתיק זה.
רצף המקדם קשור ישירות לתפקודו. יתכנו מקדמים לגנים של משק בית (יש צורך כל הזמן, אך במספר עותקים נמוך), גנים "נורמליים" (הדרושים כפי שמכתיב מצב התא, גם קצב השעתוק משתנה), גנים לתגובת לחץ (נדרשים לעיתים רחוקות) ומגוון קטגוריות אחרות. אפילו בתוך קטגוריה, רצף המקדם קובע את חוזקו. זה מבוסס על מה שמכונה "רצף הקונצנזוס". רצף הקונצנזוס הוא מקדם תיאורטי "הטוב ביותר" המבוסס על סקר של כל הגנים בקטגוריה מסוימת. האיור שלהלן מציג יישור של רצפי הפרומוטור של מגוון גנים שונים, שכולם מוסדרים על ידי אותו סוג של מקדם וחלבון קושר מקדם. התיבות המודגשות מציגות אזורים שבמרכזם -35 (35 נוקלאוטידים במעלה הזרם של אתר ההתחלה) ו -10.
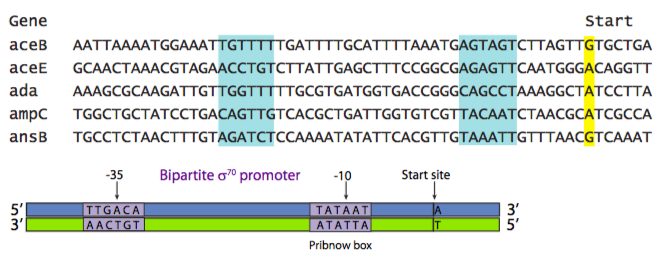
רצף הקונצנזוס באיור \(\PageIndex{2}\) מציג את הנוקלאוטיד הנפוץ ביותר שנמצא בכל מיקום בתוך אותם אזורי דמיון. בדוגמה זו מוצג האמרגן הפרוקריוטי הנפוץ ביותר: מקדם σ 70, הנקרא כך מכיוון שהוא מוכר ומחובר על ידי גורם השעתוק σ 70. [כאן, ועל פי מוסכמה מקובלת אוניברסלית, רצפי זיהוי של DNA נכתבים מכיוון שהנוקלאוטידים יתרחשו בין 5 'ל -3' בגדיל החוש, או שאינו תבנית.] זהו מקדם דו-חלקי, עם אזור שבמרכזו -35 (קונצנזוס TTGACA), ואזור (המכונה לפעמים תיבת Pribnow, קונצנזוס TATAAT) שבמרכזו -10. הסימן (-) מציין שהנוקלאוטיד נמצא "במעלה הזרם" של אתר ההתחלה. במעלה הזרם פירושו "משמאל" כאשר הנוקלאוטידים נכתבים כמחרוזת אותיות, ופירושו "בצד 5' של" ביחס לכיווניות 5'-3' של גדיל DNA. שימו לב לקשר בין המקדמים הבודדים השונים לבין רצף הקונצנזוס. באופן כללי, אותם מקדמים עם יותר התאמות לרצף הקונצנזוס הם מקדמים חזקים יותר.
לפני כמה פסקאות, המשימה של האמרגן הוגדרה כשליטה בתדירות התעתיק. איך זה עושה את זה? מה המשמעות של להיות מקדם חזק יותר (או חלש יותר)? ראשית, זכור כי הביטוי של כל גן נתון אינו אוטומטי, או 100%. בכל נקודת זמן, רבים מהגנים של התא יהיו קרובים ל -0%, או יכבו. עם זאת, אפילו גנים המופעלים מתועתקים בקצב שונה. אחד הגורמים השולטים הוא ההכרה באתר המקדם על ידי פולימראז ה- RNA. עבור מקדמים חזקים יותר, פולימראז ה- RNA נוטה יותר לזהות את האתר, לעגון כראוי, לפתוח את הסליל הכפול ולהתחיל לתמלל. מצד שני, פולימראז ה- RNA יכול לזהות מקדמים חלשים יותר, אך יש פחות סיכוי לעשות זאת, במקום להעביר אותו כסתם עוד קטע DNA לא חשוב. אמנם זה חלקית עניין של הכרה על ידי הפולימראז, אך זכור כי הוא נשלט למעשה על ידי הכרה ברצף הפרומוטור על ידי גורמי השעתוק הכלליים (שיידונו בקרוב) כגון גורמי סיגמא בפרוקריוטים המוכרים על ידי הפולימראז.
שימו לב שיש שיעור גבוה של (A) דנינים ו- (T) הימינים ברצפי האמרגן σ 70. זה נכון לגבי מקדמים רבים בגנים פרוקריוטים ואיקריוטים כאחד. כפי שבטח חשדתם, זה יתרון מכיוון שיש רק שני קשרי H בין זוגות A-T (בניגוד ל-3 קשרי H בין זוגות G-C), מה שאומר שקל יותר לפתוח 33%.