7.7: טלומרים
- Page ID
- 208724
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)אם יש מנגנון לזיהוי קצוות רופפים של DNA, מה לגבי הקצוות של כל כרומוזום אוקריוטי? הם כרומוזומים ליניאריים, אז יש להם קצוות, נכון? מה מונע ממערכות התיקון של שבירת גדילים כפולים לזהות לא נכון את כולן כדנ"א שבור ולשרשר את כל הכרומוזומים יחד? מעניין שהתשובה לשאלה זו קשורה קשר הדוק לתשובה לבעיית השכפול הסופי, שנרמז בקצרה בתיאור השכפול שלנו.
בעיית השכפול הסופית היא כזו שמשפיעה על כל הכרומוזומים הליניאריים. זה מסתכם בעובדה אחת פשוטה: יש צורך בפריימר RNA כדי להתחיל כל שכפול DNA. אז בקצה 5' של כל גדיל נמצא פריימר RNA (איור \(\PageIndex{24}\) בצהוב) שמוסר על ידי תהליך תיקון השגיאות. כך עם כל סבב שכפול, מידע הולך לאיבוד מקצה 5' של כל גדיל של כל כרומוזום.
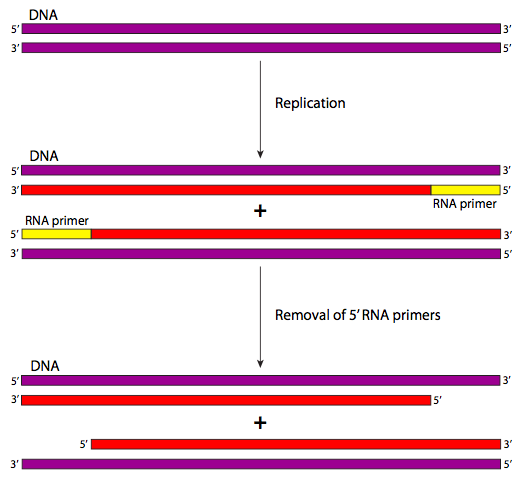
בסופו של דבר, גנים מכריעים הולכים לאיבוד והתא ימות; סביר להניח שתפקודים סלולריים רבים ייפגעו הרבה לפני שזה יקרה. הפתרון לבעיית השכפול הסופי עשוי להיחשב יותר כטיפול בסימפטומים מאשר תרופה, כדי להשתמש באנלוגיה לרפואה. בקיצור, בשלבים המוקדמים מאוד של חיי האורגניזם, הרבה DNA שאינו מקודד מתווסף לקצות ה-DNA כך שככל שהתא וצאצאיו ממשיכים להתרבות, הנוקלאוטידים אינם משפיעים על גנים פונקציונליים כלשהם. תהליך זה מזורז על ידי האנזים טלומראז.
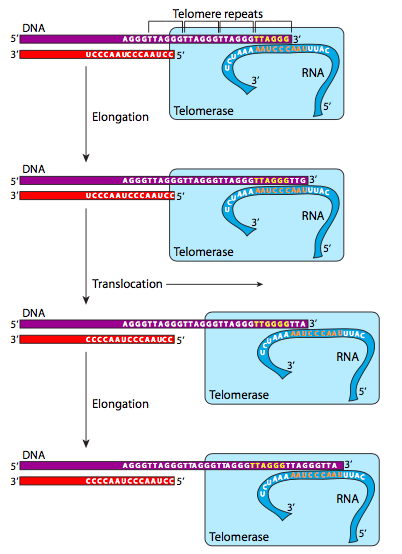
טלומראז הוא הולואנזים גדול הפועל כטרנסקריפטאז הפוך, וקורא תבנית RNA עצמאית כדי להוסיף על רצף הטלומרים לקצוות 3' של כרומוזומים ליניאריים. שים לב שזה לא מוסיף לקצוות 5' - כאמור לעיל, אין מה לעשות בקשר לקצוות 5' ישירות. עם זאת, בעוד הטלומראזות פעילות, ניתן להרחיב את קצוות ה-3', וכך כאשר מסירים את פריימר הקצה 5', הרצף שאבד (לנצח) מאותו גדיל ה-DNA הוא רק חזרה טלומרית ולא משהו שימושי יותר. החזרות נשמרות היטב על פני אוקריוטים, ונשמרות כמעט לחלוטין על פני מיני יונקים (מוצג באיור\(\PageIndex{25}\)).
במטאזואנים, פעילות הטלומראז גבוהה בשלבי החיים העובריים, אך כמעט ואינה קיימת במבוגרים למעט בסוגי תאים שחייבים להתרבות כל הזמן (למשל דם ותאי אפיתל). פעילות הטלומראז מווסתת בעיקר על ידי ביטוי של הגן TERT (telomerase reverse transcriptase) אם כי בניית הטלומראז המלא דורשת גם ביטוי של הגן TERC (RNA הטלומראז, גם הוא מקוצר TR) ודיסקרין. באופן גס, מספר החזרות הטלומריות המונחות על כרומוזום בהתפתחות מוקדמת קובע את מספר שכפול ה-DNA וחלוקות התאים שהתא יכול לעבור לפני שהוא נכנע לאפופטוזיס (מוות תאי מתוכנת). ניסויים על תאים בתרבית מדגימים מתאם חזק בין אורך הטלומרים לאריכות ימים, וידוע שלתאים שנלקחו מאנשים עם מחלת ההזדקנות המוקדמת, פרוגריה, יש טלומרים קצרים יחסית.
לעומת זאת, לתאים סרטניים יש כמעט באופן אוניברסלי ביטוי לא מוסדר של טלומראז. בהתחשב בכך שמאפיין מגדיר של תאים סרטניים הוא היכולת להתרבות במהירות וללא הגבלת זמן, הפעלת הטלומראז היא, באופן לא מפתיע, היבט חשוב של קרצינוגנזה. לכן זהו יעד לטיפולים נגד סרטן; עם זאת, עד כה אף טיפולים המכוונים לטלומראז לא הוכחו כיעילים.
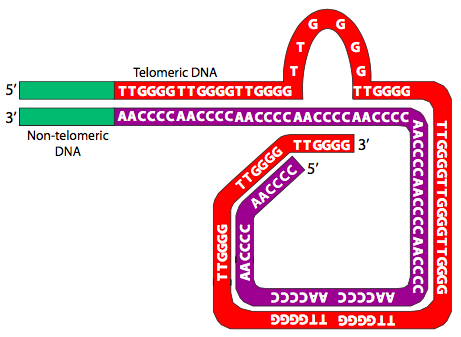
כעת, כשאנחנו יודעים על טלומרים, השאלה שהתחילה את החלק הזה הופכת לבעייתית עוד יותר: עם תלייות הרצף החוזרות ונשנות הללו, כיצד מונעים מכרומוזומים להתחבר מקצה לקצה באמצעות תהליך דמוי תיקון כפול גדילים? בין השאר בשל הרצפים החוזרים ונשנים שלהם, הטלומרים מסוגלים ליצור כובעי קצה ולהגן על קצוות כרומוזומליים. הטלומרים מגנים על הקצוות של כל כרומוזום על ידי קשירה לחלבוני מגן ועל ידי יצירת מבנים מורכבים. חלבונים קושרים לקצה הטלומרים (TEBP) נקשרים לקצה התלוי 3 'של הטלומרים. חלבוני מכסה אחרים, כגון TRF1 היונקים ו- TRF2 (גורמי קישור חוזרים של טלומרים) לא רק קושרים את הטלומרים, אלא עוזרים לארגן אותו למבנים גדולים עם לולאה המכונים לולאות T (איור). \(\PageIndex{26}\)
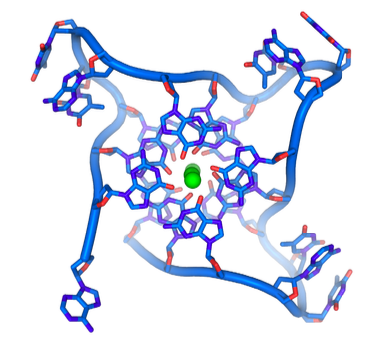
לבסוף, קצוות לולאת T מיוצבים עוד יותר על ידי היווצרות של G-quartets (איור). \(\PageIndex{27}\) רביעיות G הן טטרמרים מחזוריים שיכולים להיווצר ברצפים עם ארבעה שאריות גואנין רצופות, אשר נקשרות מימן זו לזו כדי ליצור צורה מרובעת מקושרת המייצבת על ידי יון מתכת במרכז. יתר על כן במקרים כמו הטלומרים, שבהם רצפים כאלה חוזרים על עצמם, רביעיות ה- G יכולות לערום ולהתחבר בתלת מימד, ולהגדיל את יציבותן.