
7.6: תיקון DNA
- Page ID
- 208736

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מוגדר בקפדנות, מנגנון התיקון הפשוט ביותר אינו משתמש באנזים. דיאלקילציה, או הסרה של קבוצות אלקיל (כמו -CH 3 או —C 2 H 5) כרוכה רק בהעברת קבוצת אלקיל מ-O6-מתילגואנין או O6-אתילגואנין על אלקילטרנספרז O6-אלקילגואניל-DNA. למרות השם, האלקילטרנספראז אינו באמת אנזים, מכיוון שהוא משתנה לצמיתות ומופעל על ידי התגובה ולכן אינו מתאים להגדרת זרז. שים לב שזה לא מתקן אלקילציה ב-N7 או באתרים אחרים, רק באתרים המקושרים ל-O6.
מנגנון התיקון הפשוט הבא הוא ניתוק דימרים של פירמידין ציקלובוטיל. ניתן להשיג זאת באמצעות פעילות של פוטוליאזות DNA, הידועות גם בשם אנזימים מפעילים פוטו. אלה נקראים לא רק בגלל שהיווצרות הדימרים הפירימידין ציקלובוטיל נובעת בדרך כלל מחשיפה לאור UV, אלא מכיוון שאנזימי התיקון עצמם דורשים חשיפה לאור (300-500 ננומטר, ליד UV לכחול גלוי) כדי לזרז את תגובת שבירת הדימר.
ליתר דיוק, פוטוליאזות DNA (חלבון ~ 60kD), קשורות באופן לא קוולנטי לכרומופור (N 5, N 10 -מתילנילטטרהידרופולאט או 5-דיאזפלבין) ו-FADH—. הפוטוליאז נקשר לדימר הפירימידין ציקלובוטיל של DNA חד-גדילי או דו-גדילי באופן בלתי תלוי באור ובלתי תלוי ברצף. עם זאת הוא אינו מזרז שום שינוי בקשר עד שהאור נספג בכרומופור, אשר לאחר מכן מעביר את האנרגיה ל-FADH—, גורם לו להיפלט ולאלקטרון לדימר, ובכך מפרק אותו.
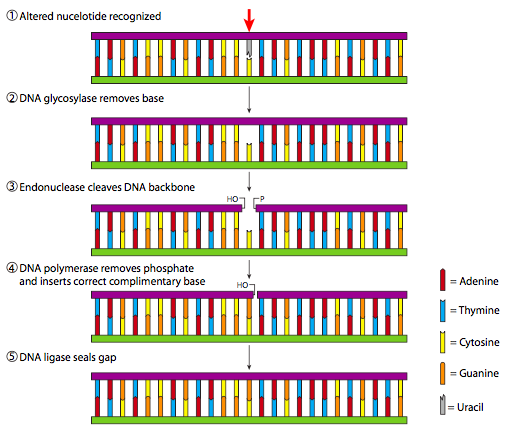
בעוד שדיאלקילציה ותמוגה דימר הם תהליכים פשוטים יחסית שעושים רק שינוי עדין ב-DNA, מנגנוני תיקון הכריתה מסובכים יותר ודורשים שלבים אנזימטיים מרובים כדי להשלים. כאשר נגע קטן (לא מגושם מבחינה סטרית) מוגבל לבסיס יחיד, בין אם חסר דפורינציה ובין אם נוצר בצורה לא נכונה עקב דמינציה או שילוב שגוי, התהליך המכונה תיקון כריתת בסיס (BER) מופעל. כפי שמודגם באיור\(\PageIndex{20}\), אם מזהים בסיס לא קונבנציונאלי, הוא מוסר על ידי גליקוזילאז DNA מתאים. נכון לעכשיו (חיפוש Genbank, יולי 2009), ישנם לפחות 8 גנים ספציפיים המקודדים לגליקוזילאזות DNA אנושיות, אם כי שלושה מקודדים גליקוזילאזים המזהים אורציל במצבים שונים. לאחר הסרת הבסיס על ידי הגליקוזילאז, מגויס אנדונוקלאז כדי לשבור את קשרי הפוספודיסטר מאשר להחזיק את הפוספודאוקסיריבוז הריק כעת. הפער שנוצר ב-DNA מתמלא על ידי פולימראז DNA ולבסוף הגדיל מחובר מחדש על ידי ליגאז DNA.
במקרה של נגעים מגושמים המשנים באופן משמעותי את ההצגה הפיזית של ה- DNA לפולימראזות ולאנזימים אחרים המעבדים DNA, מעורב סוג אחר של תהליך תיקון. תיקון כריתת נוקלאוטידים (NER), אולי בשם טוב יותר תיקון כריתת פולי נוקלאוטיד, כרוך בהסרת הנגע כמו גם בחלק מהנוקלאוטידים בסביבה הקרובה. ישנם שני יוזמים עיקריים של NER: או שחלק לא פעיל מבחינה תעתיק של ה-DNA נסרק על ידי XPC (איור \(\PageIndex{21}\) A), שמזהה נגע מגושם ומגייס את קומפלקס התיקון, או כאשר גן מתועתק, RNA פולימראז נתקל בנגע, ולאחר מכן מגייס את מתחם התיקון באמצעות CSA ו-CSB (איור F ו-G). \(\PageIndex{21}\) אם הזיהוי הוא באמצעות XPC, אחד מגורמי התיקון המוקדמים שגויסו לאתר הוא גורם שעתוק IIH/XPB/XPD, שהוא הליקאז DNA (איור B). \(\PageIndex{21}\) סוג זה של זיהוי גנום עולמי אינו יעיל ואיטי יחסית, אך מספק רמה בסיסית של בדיקת שגיאות עבור כל ה- DNA. במקרה של תמלול DNA, קומפלקס ה- RNA פולימראז כבר כולל TFIIH, מתוכם XPB ו- XPD הם חלק. זיהוי מכוון תעתיק זה יעיל יותר ומכוון לאותם חלקים של ה- DNA בשימוש הגדול ביותר בתא נתון. בשלב הבא (איור \(\PageIndex{21}\) C), XPG, המשויך ל- BRCA1/2 ו- XPF, הקשורים ל- ERCC1, מוציאים חלק מהגדיל המושפע, כולל אך לא מוגבל לנגע עצמו. DNA פולימראז δ או ε יכול לאחר מכן להוסיף ל-3'OH החופשי ל-ll בפער המבוסס על רצף הגדיל המשלים (איור \(\PageIndex{21}\) D). לבסוף, התיקון מחובר בקצה 3' שלו לשאר הגדיל על ידי ליגאז DNA (איור \(\PageIndex{21}\) E).
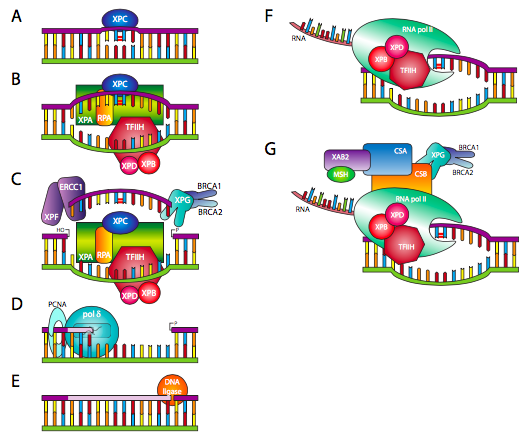
ה-"XP" ב-XPC, XPB, XPD והאחרים באיור \(\PageIndex{21}\) מתייחס ל-xeroderma pigmentosa, מחלה אוטוזומלית רצסיבית נוספת, שהמאפיין העיקרי שלה הוא היווצרות קרצינומות בעור בגיל צעיר. מכיוון ש- NER היא צורה עיקרית של תיקון דימר פירמידין (בנוסף לפוטוליאזות), שיבושו על ידי מוטציות לאחד או יותר מהגנים XP מוביל לרגישות קיצונית לנגעים הנגרמים על ידי UV. אנשים מושפעים חייבים למזער את החשיפה לשמש. שם המחלה נובע מהנגעים הפיגמנטיים האופייניים (קרטוזות) הנוצרים לעיתים קרובות על העור כאשר הם נחשפים לשמש.
CSA ו- CSB נקראים על שם תסמונת קוקיין, הפרעת הזדקנות אוטוזומלית רצסיבית. מוטציות בכל אחד מהגנים עלולות לגרום להפרעה, המאופיינת בהזדקנות מוקדמת, צמיחה מעוכבת, רגישות לאור ופגמים התפתחותיים של מערכת העצבים. יש להניח כי ביטול יכולת תיקון ה-DNA של CSA או CSB מוביל להצטברות מהירה של נזק, חוסר יכולת לתמלל גנים נחוצים ובסופו של דבר למוות של תאים.
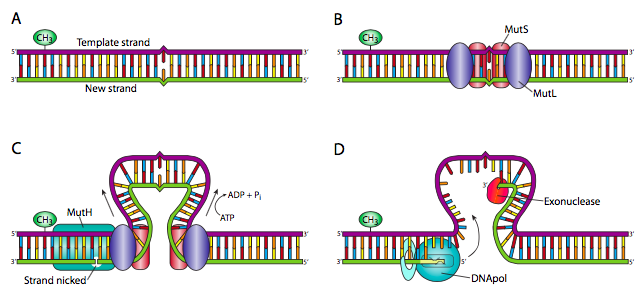
מעין וריאציה ב- NER היא מערכת תיקון אי התאמה (MMR). זה מובן בצורה הטובה ביותר בפרוקריוטים: ב אי - קולי, MUTs הוא חלבון קטן היוצר הומודימרים באתרי חוסר התאמה. הדימרים של MutS מגייסים שני חלבוני MutL, שכל אחד מהם מקיים אינטראקציה עם אחת מיחידות ה- MutS. כל קומפלקס Muts/Mutl דוחף את ה-DNA פנימה, ויוצר לולאה עם חוסר ההתאמה במרכז הלולאה. זה נמשך עד שאחד ממתחמי Muts/Mutl נתקל ברצף GATC המימתיל. זה גורם לגיוס של MutH, אנדונוקלאז מיוחד מאוד שיוצר ניק חד גדילי בעמוד השדרה של הגדיל הלא מתיל. זה מספק פתח לאקסונוקלאז I 3'-5' או לאקסונוקלאז VII 5'-3' (או RecJ) כדי לפרק את הגדיל מהניק עד כדי חוסר התאמה. זה אז, כפי שאולי ניחשתם, מתמלא על ידי DNA פולימראז ועמוד השדרה המחובר על ידי ליגאז. באאוקריוטים התגלו מספר הומולוגים לחלבוני MutS ו- MutL והתהליך דומה, אך עדיין לא מובן בבירור, מכיוון שטרם התגלה הומולוג ל- MutH.
נזכיר כי ב אי - קולי, Dam methyltransferases בסופו של דבר מתילטים את ה- DNA כשיטה להגנה על הגנום שלו, אך DNA שסונתז לאחרונה אינו מתילט. לפיכך, ההנחה היא שהגדיל המתילט מכיל את הבסיס המקורי והנכון, בעוד שחוסר ההתאמה נובע משילוב שגוי בגדיל החדש יותר.
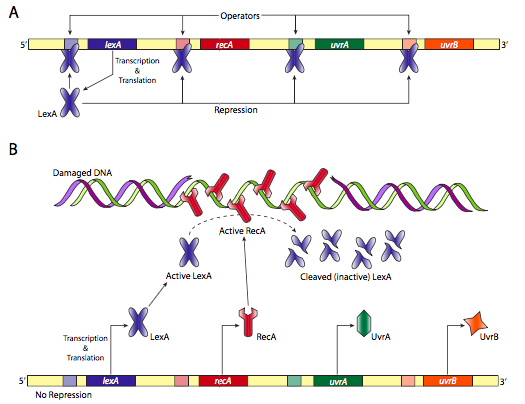
מערכת תיקון DNA פרוקריוטית נוספת היא תגובת SOS. כפי שמתואר ב דמות \(\PageIndex{23}\) להלן, אם אין נזק, RecA אינו פעיל, כך שחלבון LexA יכול להדחיק את הייצור של חלבוני תיקון SOS נוספים. עם זאת, אם יש נזק, חלבוני RecA נקשרים ל-DNA החד-גדילי ומופעלים. הם בתורם מבקעים את מדכא LexA המאפשר ייצור ממספר גנים לתיקון DNA.
עד כה, התיקונים התבססו על ההנחה שנגע משפיע רק על גדיל אחד, והגדיל השני יכול לספק תבנית אמינה לביצוע תיקונים ללא אובדן מידע. למרבה הצער, זה לא תמיד המקרה, וכמה נגעים ותהליכי תיקון מובילים בהכרח לאובדן רצף. כאשר מתרחשת הפסקה כפולה, אולי כתוצר של קרינה מייננת, מנגנון התיקון הנפוץ ביותר ידוע בשם חיבור קצה לא הומולוגי (NHEJ). הקצוות הדו-גדיליים מזוהים לראשונה על ידי Ku, חלבון מעגלי הטרודימרי הקושר את קצוות ה-DNA. לאחר מכן Ku מגייס את קינאז DNA-PK CS. ה- DNA-PK CS פועל כגשר לקירוב שני הקצוות, ולאחר מכן ליגאז DNA יכול לחבר את הקצוות יחד. אם הגדילים נשברו במקומות שונים, וכתוצאה מכך התלויות חד-גדיליות משלימות בכל קצה (כמו אלה שנוצרו על ידי אנדונוקלאזות הגבלה מסוימות) אז התיקון לרוב מושלם, מכיוון שהרצפים המשלימים מיישרים את שני הקצוות בצורה נכונה במיקומם המקורי. עם זאת, אם קצוות הגדיל כבר הופעלו על ידי נוקלאזות ואינם משלימים עוד, סביר להניח שהחיבור מחדש של הקצוות יוביל לאובדן מידע. בחלקים מסוימים של ה- DNA, תהיה לכך השפעה מועטה, אך אם זה קרה בתוך גן, לתוצר הגן שעבר מוטציה יכול להיות תפקוד לא תקין או נפגע.