
7.5: נגעי DNA
- Page ID
- 208713

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)האופי החזק של ה- DNA בשל הגדילים הכפולים המשלימים שלו צוין כבר מספר פעמים. כעת אנו שוקלים ביתר פירוט את תהליכי התיקון המצילים DNA פגום. ה- DNA אינו חזק כמעט כמו שהתקשורת הפופולרית גורמת לו להיות. למעשה, כדי לקחת את הספר והסרט שוברי הקופות, פארק היורה, כדוגמה, למרות שללא ספק ניתן למצוא DNA כלשהו מוטבע בטפילים הקשורים לענבר, או אולי ברקמה רכה משומרת (שנמצאת עמוק בתוך עצם הירך המאובנת, Schweitzer et al, 2007). סביר להניח שהוא מושפל מאוד, ורבייה מדויקת בלתי אפשרית ללא דגימות רבות לעבוד מהן.
העלבון השכיח ביותר ל- DNA של אורגניזמים חיים הוא דפורינציה, שבה הקשר β-N-glycosidic בין אדנין או גואנין לדאוקסיריבוז עובר הידרוליזה. בתאי יונקים, הוא מוערך בכמעט 10000 פורינים לדור תא, ובאופן כללי, קצב האובדן הממוצע ב-pH פיזיולוגי ובחוזק יוני, וב-37 מעלות צלזיוס, הוא כ-3 x 10 -11/sec. דפירימידינציה של שאריות ציטוסין ותימין יכולה להתרחש גם היא, אך עשו זאת בקצב איטי בהרבה מאשר דפורינציה. למרות שיעור האובדן הגבוה של בסיסים אלה, הם בדרך כלל מתקנים בקלות על ידי תיקון כריתת בסיס (BER), עליו נדון בהמשך פרק זה. לכן נדיר שדפורינציה או דפירימידינציה מובילים למוטציה.
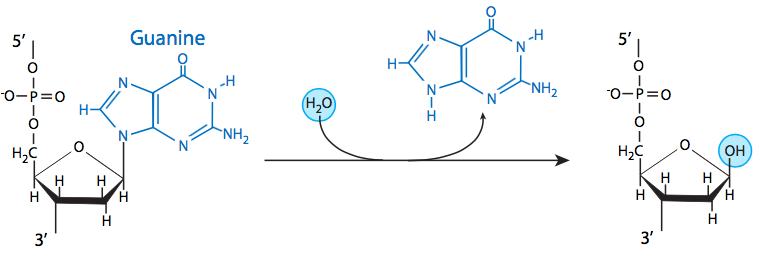
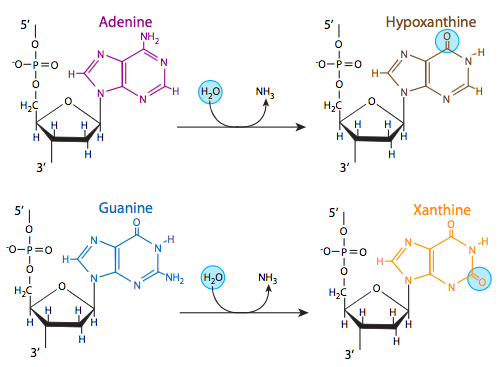
שלושה מתוך ארבעת בסיסי ה-DNA, אדנין, גואנין וציטוסין, מכילים קבוצות אמינים שיכולות ללכת לאיבוד במגוון תגובות pH ותלויות טמפרטורה הממירות את הבסיסים להיפוקסנטין, קסנטין ואורציל, בהתאמה. לפעמים זה יכול להוביל למוטציות קבועות שכן במהלך השכפול הם משמשים תבנית לסינתזה של גדיל משלים, ולאן צריך ללכת גואנין, למשל (משלים לציטוסין), ניתן להכניס אדנין (מכיוון שהוא משלים את אורציל, תוצר הדימינציה של ציטוסין).
דמינציה נוספת, של הבסיס המתוקן מתילציטוסין, יכולה להוביל גם למוטציה עם שכפול. חלק מהציטוזינים עשויים להיות מתילטים כחלק מתהליך רגולטורי להשבתת גנים מסוימים באיקריוטים, או בפרוקריוטים כהגנה מפני אנדונוקליזות הגבלה. כאשר הציטוסין המתילט מנוטרל, הוא מייצר תימין, שמשנה את הנוקלאוטיד המשלים (עם שכפול) מגואנין לאדנין. דמינציה של ציטוזינים מתרחשת כמעט באותו קצב כמו דפורינציה, אך דימינציה של בסיסים אחרים אינה נפוצה כל כך: דמינציה של אדנינים, למשל, סבירה פי 50 מדמינציה של ציטוסין.
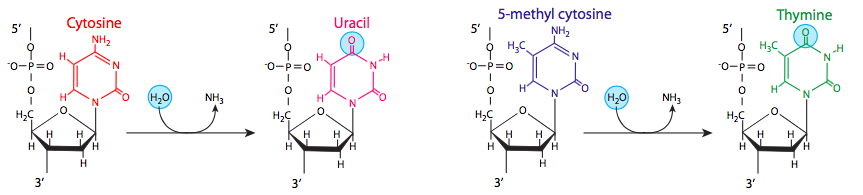
תימין טוב, אורציל רע. מדוע תימין נמצא ב- DNA ולא באורציל? מתברר שתדירות דימינציה של ציטוסין עשויה להניב רמז מדוע תאים עברו את הצעד הנוסף (תרתי משמע, מכיוון שאורציל הוא מבשר בביוסינתזה של תימין) ליצירת נוקלאוטיד "סטנדרטי" חדש ל- DNA כאשר אורציל עבד מצוין עבור RNA, ככל הנראה המולקולה הגנטית הישנה יותר. שקול זאת: אם אורציל היה סטנדרטי ל-DNA, אז המרות הדימינציה התכופות מאוד של C ל-U לא היו נתפסות על ידי בדיקת שגיאות עבור בסיסים שאינם DNA, ושיעור המוטציות היה מרקיע שחקים. למרבה המזל, מכיוון ש-T התפתח להיות השותף הסטנדרטי לזיווג בסיסים של אדנין ב-DNA, אורציל מזוהה ומוסר במהירות על ידי מספר גליקוזילאזות DNA של אורציל (עוד על כך בהמשך פרק זה), והשלמות של רצפי ה-DNA שלנו הרבה יותר בטוחה.
כל בסיסי ה-DNA יכולים לעבור באופן ספונטני לאיזומר טאוטומרי (אמינו לאימינו, קטו לאנול וכו'), אם כי שיווי המשקל נוטה בכבדות לכיוון האחד מהשני. כאשר מתרחש טאוטומר נדיר, הוא מתמזג בצורה שונה מצורתו המבנית הנפוצה יותר: גואנינים עם תימין ואדנינים עם ציטוזינים. גם כאן, ניתן להפיץ מוטציה במהלך שכפול ה- DNA.
DNA בתוך תא חייב להתמודד גם עם מינים חמצוניים תגובתיים (ROS) הנוצרים על ידי התהליכים המטבוליים של התא. אלה כוללים רדיקלים של חמצן בודד, חמצן וחמצן, כמו גם רדיקלים הידרוקסיל. אם כי נהוג לחשוב שרדיקלי מי החמצן והחמצן אינם תוקפים ישירות את ה- DNA אלא מייצרים רדיקלים הידרוקסיליים שכן. רוב ה-ROS הללו נוצרים במיטוכונדריה במהלך זרחון חמצוני ודולפים החוצה, אם כי חלקם עשויים להיווצר בפרוקסיזומים, או בתגובות ציטוזוליות מסוימות. תלוי לאיזה חלק של ה-DNA ממוקד, ROS יכול לגרום למגוון נגעים כולל שבירות גדילים והסרת בסיסים.
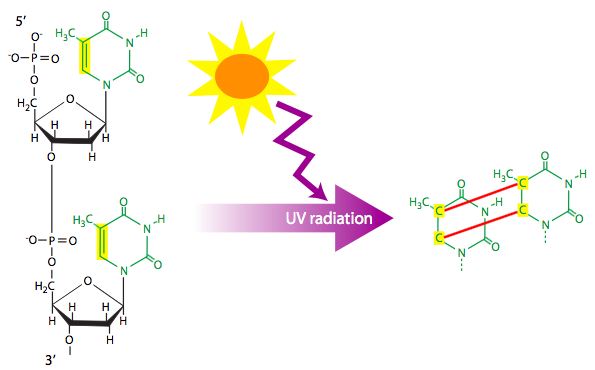
קרינה מייננת (למשל צילומי רנטגן) וקרינה אולטרה סגולה יכולים לגרום כל אחד לנגעי DNA. קרינה מייננת היא לעתים קרובות סיבה להפסקות דו-גדיליות של ה-DNA. כפי שתואר בהמשך הפרק, תהליך התיקון להפסקות דו-גדיליות מוביל בהכרח לאובדן מידע מסוים, ועלול לדפוק גן. קרינה אולטרה סגולה הפוגעת בתימינים סמוכים עלולה לגרום להם להגיב וליצור דימר תימין ציקלובוטיל (ארבעה פחמנים הקשורים בלולאה סגורה). הדימר מושך כל תימין לכיוון השני, מתוך היישור הרגיל. בהתאם לצורה המבנית של הדימר, זה מספיק כדי לעצור את מכונת השכפול ולעצור את השכפול. עם זאת, נתונים מסוימים מצביעים על כך שזיווג בסיס תקין לאדנין עשוי להיות אפשרי בתנאים מסוימים, אם כי סביר להניח שרק זוג בסיסים אחד ייווצר, והבסיס החסר עלול להוביל להחלפה אקראית או למחיקה בגדיל החדש שסונתז.
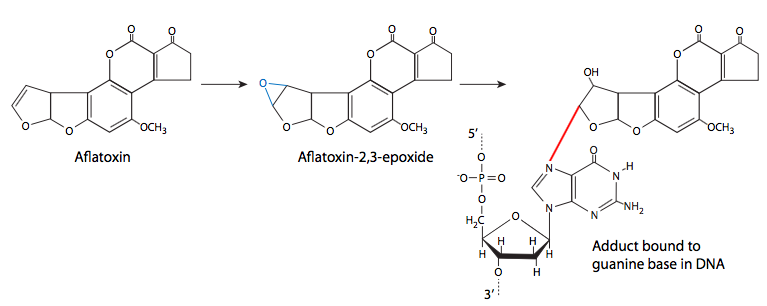
לבסוף, אנו שוקלים היווצרות של תוספים כימיים (קבוצות מחוברות קוולנטית) על DNA. הם עשויים להגיע ממגוון מקורות, כולל חמצון שומנים, עשן סיגריות ורעלים פטרייתיים. תוספות אלו נצמדות ל-DNA בדרכים שונות, כך שיש מגוון השפעות שונות גם מהתוספים. חלקם עשויים להיות תוספים קטנים מאוד - חומרים מסרטנים סביבתיים רבים הם חומרי אלקילציה, המעבירים קבוצות מתיל או קבוצות אלקיל קטנות אחרות ל- DNA. תוספות אחרות גדולות יותר, אך גם נצמדות באופן קוולנטי לבסיס חנקני של DNA. דוגמאות נפוצות הן בנזו (א) פירן, מרכיב מוטגני עיקרי של עשן סיגריות, ואפלטוקסין B1, המיוצר על ידי מגוון פטריות ממשפחת אספרגילוס. בנזו (א) פירן מומר לבנזו (א) פירון דיול אפוקסיד, שיכול לאחר מכן לתקוף את ה- DNA. כאשר זה קורה, טבעת ה-at pyrene משתלבת בין הבסיסים, וגורמת לשינויים סטריים המובילים לעיוות מקומי של ה-DNA ולהפרעה בשכפול ה-DNA הרגיל.
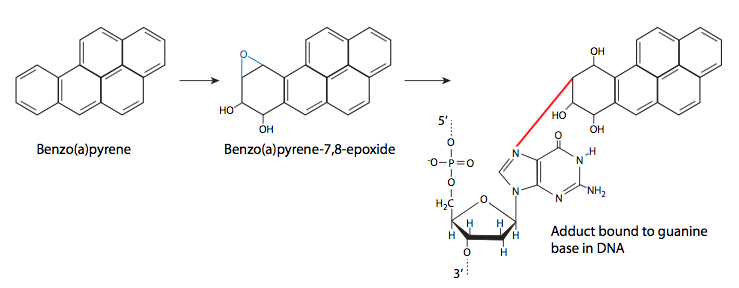
אפלטוקסין B1 הוא האפלטוקסין העיקרי המיוצר על ידי מינים מסוימים (במיוחד. פלבוס, פרזיטיקוס) של אספרגילוס, עובש נפוץ מאוד הגדל על תבואה מאוחסנת (כמו גם דטריטוס וחומר צמחי מת או גוסס אחר). בנוסף להדבקת תבואה, זוהי בעיה שכיחה בבוטנים מאוחסנים. ברמות גבוהות, אפלטוקסין רעיל באופן חריף, אך ברמות נמוכות יותר, יש לו את המאפיין החתרני להיות רעיל באופן בלתי מורגש אך מוטגני. כמו בנזו (א) פירן, הוא עובר חילוף חומרים לאפוקסיד ולאחר מכן יגיב עם DNA ליצירת תוסף שעלול לשבש את השכפול.
כמה חומרי אלקילציה, במיוחד תרכובות N-nitroso, נוצרים בתנאים החומציים של הקיבה מניטרוזציה של ניטריטים טבעיים המיוצרים ממזון (הפחתת חנקות), או ניטריטים סביבתיים במי שתייה. למרבה האירוניה, בעוד שחלק מחומרי האלקילציה עלולים לגרום לסרטן, אחרים משמשים טיפולית כטיפולים נגד סרטן, למשל מיטומיצין, מלפלן. הרעיון, כמו בטיפולים רבים בסרטן, הוא שלמרות שתרופות כאלה גורמות נזק ל- DNA לתאים לא סרטניים כמו גם לתאים סרטניים, השיעור הגבוה של התפשטות תאים סרטניים נותן להם פחות סיכויים לתיקון DNA פגום, ובכך סבירות גבוהה יותר שהנזק עלול לעצור את השכפול ולהוביל למוות של תאים.
ברוח דומה, חומרים כימותרפיים צולבים כגון ציספלטין (אטום פלטינה הקשור לשתי קבוצות כלוריד ושתי קבוצות אמינו) נקשרים גם הם ל- DNA. קבוצות הכלוריד נעקרות תחילה על ידי מים ולאחר מכן על ידי קבוצות אחרות כולל אתרים ב- DNA. למרות שלפעמים מסווג כסוכן אלקילציה, ברור שהוא לא, אבל הוא פועל באופן דומה. עם זאת, ציספלטין הולך צעד אחד קדימה מחומר אלקילציה פשוט, מכיוון שיש לו אתר תגובתי אחר ובכך יכול להצליב (לקשר קוולנטית) נוקלאוטיד אחר, אולי על גדיל אחר של DNA, מה שיוצר חסימה חזקה לשכפול ה-DNA. ציספלטין יכול גם לחבר חלבונים ל- DNA.
בנזו (א) פירן ואפלטוקסין B1 אינם בעצמם מוטגנים. ברגע שהם נמצאים בתא, חילוף החומרים הרגיל של תרכובות אלה מוביל להיווצרות דיול אפוקסיד, שיכול לתקוף את ה- DNA. למרות ש-7-חנקן (N7) של גואנין הוא נוקלאופילי יותר, ומהווה יעד לאפלטוקסין, רוב תוספי הבנזו (א) פירון דיול אפוקסיד מתחברים ל-2-חנקן של שאריות גואנין.
ישנם תקנים פדרליים (20-300 חלקים למיליארד תלוי בשימוש) לאפלטוקסין בצורות שונות של מזון לבעלי חיים על בסיס דגנים, במיוחד הזנות מבוססות תירס, מכיוון שהרעלן יכול לעבור דרך החיה לחלב, כמו גם להתעכב בבשר. בנוסף להאכלה, ישנם מקסימום פדרלי לבוטנים ומוצרי בוטנים, אגוזי ברזיל, פיסטוקים ומוצרי מזון אחרים (ניתנים לפעולה ב -20 ppb).
ובכן, מה תא עני לעשות כאשר ה- DNA שלו נהרס כל הזמן? כפי שמתברר, ישנם כמה תהליכי תיקון טובים מאוד שעובדים כל הזמן על ה- DNA, סורקים אותו לאיתור פגמים, ובמידת האפשר מבצעים תיקונים. לעתים קרובות התיקונים מושלמים, אם הגדיל המשלים שלם, לפעמים יש להכניס מוטציות, ולבסוף יש מקרים שבהם תיקון בלתי אפשרי, ואפופטוזיס מופעלת כדי להרוג את התא ולמנוע התפשטות של DNA פגום.