
7.4: שכפול אוקריוטי
- Page ID
- 208746

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)בהתחשב באופי המכריע של שכפול כרומוזומלי לקיום החיים, אין זה מפתיע לגלות ששכפול ה- DNA האוקריוטי דומה מאוד לתהליך הפרוקריוטי. חלק זה ידגיש כמה מההבדלים, שהם בדרך כלל פירוט על הגרסה הפרוקריוטית.
שלא כמו פרוקריוטים, לכרומוזומים אוקריוטיים יש לעתים קרובות מקורות שכפול מרובים. בהתחשב בגודל הכרומוזומים האוקריוטיים, הדבר הכרחי כדי לסיים את השכפול המלא בזמן. כל אחד ממקורות אלה מגדיר העתק, או את קטע ה- DNA המשוכפל ממקור מסוים. ההעתקים אינם משתכפלים באותו זמן בדיוק (אם כי כולם באותו שלב של מחזור התא, ראה פרק 15), לכן חשוב לוודא כי נעשה שימוש בשכפולים פעם אחת בלבד במהלך מחזור התא.
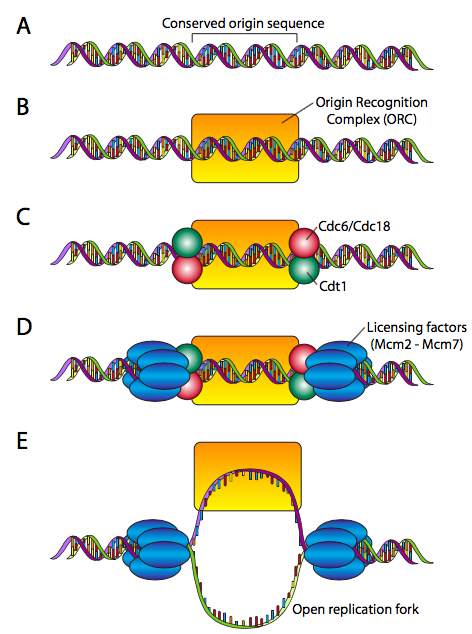
זה דורש מנגנון "רישוי". במהלך שלב מחזור התא לפני תחילת שכפול ה-DNA, מורכב קומפלקס טרום-רפליקטיבי בכל מקור (איור\(\PageIndex{13}\)). המקורות משתנים מאוד בהרכב ובאורך, נעים בין ~ 100 ליותר מ -10000 זוגות בסיס. החלבונים שלפני RC, לעומת זאת, שמורים מאוד. ה- Pre-RC מתחיל ביצירת ה- ORC (קומפלקס זיהוי מוצא, לא יצור הנלחם בפרודו ובאראגורן), המורכב משש יחידות משנה (Orc1-Orc6). למרות שאין הומולוגיה רצף משמעותית, ה- ORC מתקרב לתפקוד חלבון ה- DnaA ב אי - קולי. להשלמת ה- pre-RC, ה- ORC מגייס זוג חלבונים, Cdc6 ו- Cdt1 לכל צד, והם קושרים את קומפלקס MCM (הקסאמר של Mcm2-Mcm6 בעל פעילות הליקאז לא פעילה), מה שמוביל ל- Pre-RC המורשה במלואו. המקור מוכן כעת להפעלה.
הפעלת ה- pre-RC במקור שכפול דורשת תחילה Mcm10, מה שמקל על קינאזות חלבון המזרחנות את קומפלקס MCM, מפעילות את פעילות ההליקאז והפיכת מזלג השכפול מוכן לקבל את מכונת השכפול.
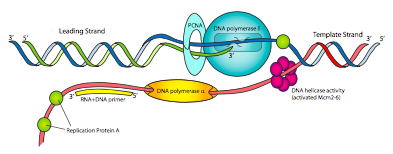
למרכיבי מכונת השכפול האוקריוטית עשויים להיות שמות שונים מהפרוקריוטי, אך הפונקציות צריכות להיות מוכרות מאוד (איור\(\PageIndex{14}\)). יש פרימאז לייצור פריימרים של RNA, וקשור הדוק לפרימאז הוא DNA פולימראז α. ל-Pol α אין אקסונוקלאז 5'-3' ולא אקסונוקלאז המגהה 3'-5', אבל הוא יכול לסנתז DNA. עם זאת, זה לא פולימראז ה- DNA העיקרי. קומפלקס הפרימאז/פול a הוא בעצם פרימאז מהודר שמתחיל בפריימר RNA קצר & lt10nt ולאחר מכן מוסיף עוד שבר DNA קצר ~ 15nt, מה שהופך פריימר היברידי.
גורם שכפול C (RFC), הפועל כמו קומפלקס מעמיס המהדק הפרוקריוטי, ואז מחליף את pol α ב- PCNA, הגרסה האוקריוטית של מהדק β. לאחר מכן זה מגייס DNA פולימראז δ, שהוא פולימראז ה- DNA המשכפל העיקרי, המקביל לתפקוד Pol III הפרוקריוטי, והכרחי לסינתזת גדילים מובילים ומפגרים כאחד. לבסוף, במקום SSB, תאים אוקריוטיים משתמשים בחלבון שכפול A (RPA) כדי לארגן ולשלוט ב- DNA חד-גדילי כפי שהוא נוצר במהלך תהליך השכפול.
אולי שמתם לב שלאף אחד מפולימראזות ה-DNA האוקריוטיות שנדונו עד כה אין פעילות אקסונוקלאז של 5' עד 3' כפי ששימשה את ה-DNA פולימראז I הפרוקריוטי להסרת פריימרים. במקום זאת, RNAseH1 ו- FEN-1 מסירים את פריימרים של RNA (כולם מלבד ריבונוקלאוטיד אחד, ואת הריבונוקלאוטיד האחרון, בהתאמה). מעניין לציין כי FEN-1 גם חותך נתחים של DNA גדיל בפיגור בתוך כ-15 זוגות בסיסים של ה-RNA אם הם מכילים טעויות. נראה שזה עוזר להקל על הבעיה של נאמנות נמוכה יותר של שכפול על ידי pol α, שאין לו יכולת הגהה. לאחר ש-RNAseH1 ו-FEN-1 הסירו פריימרים וטעויות כמעט פריימר, pol δ ממלא את הנוקלאוטידים החסרים, ואנזים ליגאז מצטרף לשברים. ל-Pol ε יש פעילות אקסונוקלאז 3'-5' הקוצצת DNA חד-גדילי לשברי אוליגונוקלאוטידים קטנים והיא קשורה גם למכונת השכפול. הפונקציה של Pol ε אינה מובנת בבירור.