
7.3: שכפול פרוקריוטי
- Page ID
- 208737

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)שכפול ה- DNA מתחיל במקור שכפול. יש רק מקור אחד בפרוקריוטים (ב- E. coli, ORiC) והוא מאופיין במערכים של רצפים חוזרים ונשנים. רצפים אלה עוטפים חלבון קושר DNA, ובכך מפעילים לחץ על קשרי ה- H בין גדילי ה-DNA, והכרומוזום מתחיל להיפתח באזור עשיר ב-AT עטוף סביב חלבון זה. זכור כי זוגות A-T חלשים ב -33% מזוגות G-C בגלל פחות קשרי מימן. השימוש בקטעי DNA עשירים ב-AT כנקודות הפרדת גדילים הוא נושא שחוזר על עצמו באמצעות מגוון פעולות DNA. ההפרדה בין שני הגדילים היא דו כיוונית, ופולימראזות DNA יפעלו בשני הכיוונים על מנת לסיים את התהליך במהירות האפשרית. המהירות חשובה כאן מכיוון שבעוד שכפול מתרחש, ה- DNA חשוף לשבירה, ורוב התהליכים המטבוליים מושבתים כדי להקדיש את האנרגיה לשכפול. אפילו בפרוקריוטים, שבהם מולקולות ה-DNA קטנות בסדרי גודל מאשר באיקריוטים, גודל מולקולת ה-DNA כשהיא נפרמת מחלבוני אריזה מגנים הופך אותה לרגישה מאוד לנזק פיזי רק מתנועות התא.
החלבון המחייב oriC הראשון, DnaA, נקשר לקופסאות DnaA, שהן 9 מקטעי זוג בסיסים עם רצף קונצנזוס של TTATCCACA. ל-ORiC יש חמש מהחזרות הללו, וחלבון DnaA אחד נקשר לכל אחת מהן. HU ו-IHF הם חלבונים דמויי היסטון המתחברים ל-DnaA ויחד מכופפים את החלק הזה של ה-DNA ללולאה מעגלית, וממקמים אותו רק מעל התכונה העיקרית האחרת של oriC, החזרות העשירות ב-AT של 13 bp (GATCTNTNTNTTTTT). DnaA מבצע הידרוליזה של ATP ושובר את קשרי ה- H בין גדילים בחזרות 13mer, הידועות גם בשם המסת ה- DNA. זה מאפשר קומפלקסים של DnaB [ו- DNAc, שהוא חלבון טעינה המסייע בחיבור DnaB (6) לגדיל עם הידרוליזה נלווית של ATP. כמו כן, חמישה DnaA נוספים מגויסים לייצוב הלולאה.] להיקשר לכל אזור חד-גדילי של ה-DNA בצדדים מנוגדים של בועת השכפול החדשה שנפתחה.
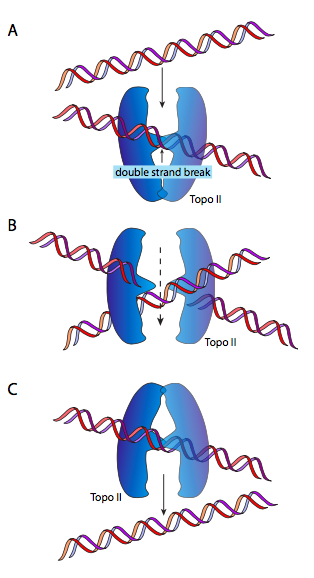
DnaB הוא הליקאז; הפעילות האנזימטית שלו היא לפתוח/לפרוק את ה- DNA לפני פולימראז ה- DNA, לתת לו DNA חד גדילי לקרוא ולהעתיק. הוא עושה זאת בשיתוף עם חלבונים קושרים ל-DNA חד-גדילי (SSB) ו-DNA gyrase. הפונקציה של SSB כמעט מובנת מאליה: DNA חד-גדילי הוא כמו RNA ביכולתו ליצור מבנים משניים מורכבים על ידי זיווג בסיסים פנימי, ולכן SSB מונע זאת. DNA gyrase הוא טופואיזומראז מסוג II, ומוטל עליו להכניס סליל-על שלילי ל-DNA. זה הכרחי מכיוון שפריקת ה- DNA על ידי הליקאז גם משחררת אותו (מכיוון שמדובר בסליל כפול) וגורמת להכנסת סליל-על חיובי. המשמעות היא שכל המולקולה המעגלית מתפתלת על עצמה: דמיין להחזיק גומייה בשתי ידיים ולסובב אותה. ככל שסליל העל מצטבר, ה-DNA הופך מפותל חזק יותר, עד כדי כך שאי אפשר יהיה להליקאז לפתוח אותו. DNAb/gyrase יכול להקל על הלחץ הזה על ידי חיתוך זמני של ה-DNA הדו-גדילי, העברת לולאה של המולקולה דרך הפער ואטימתו מחדש.
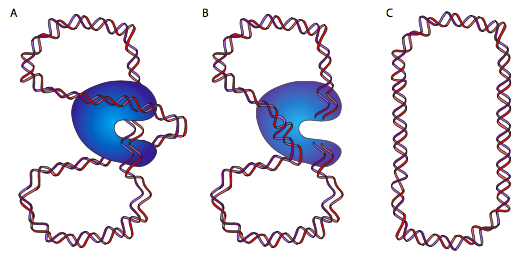
זה (אני מקווה) הגיוני הרבה יותר להסתכל על התרשים. לחלופין, כשחוזרים לגומייה שלנו, תנו לגומייה טוויסט או שניים ואז הדביקו את שני הקצוות. אם אתה חותך את הגומייה ומעביר חלק סמוך של הגומייה דרך החיתוך הזה, ואז חבר מחדש את הקצוות החתוכים, תגלה שיש טוויסט אחד פחות. נחמד, אה? בשלב זה, חלקכם יגידו, אבל אם תסובבו רצועת גומי צפה חופשית, כפי שאפשר לדמיין כרומוזום DNA מעגלי צף חופשי ב אי - קולי, הייתם מצפים שהוא יתנתק באופן טבעי. מבחינה טכנית, כן, אבל בגלל המסה הגדולה של הכרומוזום, הקשר שלו עם חלבונים שונים וקרום התא, וצמיגות סביבתו, הוא אינו מתנהג כאילו הוא חופשי לחלוטין.
לאחר פתיחת oriC וההליקסות התחברו לשני הצדדים של מזלג השכפול, מכונת השכפול, המכונה הרפליזום יכולה להתחיל להיווצר. עם זאת, לפני שפולימראזות ה-DNA תופסות עמדות, הן צריכות להיות דרוכות. פולימראזות DNA אינן מסוגלות לחבר שני נוקלאוטידים חופשיים בודדים יחד כדי להתחיל ליצור חומצת גרעין; הם יכולים להוסיף רק על גדיל קיים של לפחות שני נוקלאוטידים. לכן, פולימראז RNA מיוחד (ל- RNAP אין מגבלה זו) המכונה פרימאז הוא חלק מהרפליזום, וקריאות יוצרות גדיל RNA קצר המכונה פריימר עבור פולימראז ה- DNA להוסיף עליו. למרות שיש צורך רק בכמה נוקלאוטידים, הפריימרים הפרוקריוטים עשויים להיות ארוכים עד 60 nt בהתאם למין.
לפחות חמישה פולימראזות DNA פרוקריוטיות התגלו עד כה. פולימראז ה- DNA העיקרי לשכפול ב אי - קולי הוא DNA פולימראז III (Pol III). פול I מעורב גם במנגנון הבסיסי של שכפול ה-DNA, בעיקר כדי לאתר פערים שנוצרו במהלך סינתזת גדילים בפיגור (מוגדר 3 עמודים קדימה) או באמצעות מנגנוני תיקון שגיאות. DNA פולימראז II וה-Pol IV ו-Pol V שהתגלו לאחרונה אינם משתתפים בשכפול כרומוזומלי, אלא משמשים לסינתזת DNA כאשר יש צורך בסוגים מסוימים של תיקון בזמנים אחרים במחזור החיים התאי.
DNA פולימראז III הוא הולואנזים מרובה תת-יחידות, עם יחידות משנה α, ε ו- θ הכוללות את פולימראז הליבה, ו τ, γ, δ, δ ', χ, Ψ ו- β מתאחדים ליצירת ההולואנזים השלם. לפולימראז הליבה שתי פעילויות: יחידת המשנה α היא פונקציית הפולימראז, קוראת קווצת DNA ומסנתזת גדיל משלים במהירות רבה, בסביבות 150 nt/sec; יחידת המשנה ε היא אקסונוקלאז "הגהה" 3'-5' ומשמשת כמגיה מיידית, ומסירה את הנוקלאוטיד האחרון אם הוא לא נכון. קריאת הוכחה זו אינה מגיעה לאחור יותר: היא פועלת רק על הנוקלאוטיד שנוסף לאחרונה כדי לתקן שילוב שגוי. מנגנונים ואנזימים אחרים משמשים לתיקון נגעי DNA המתעוררים בזמנים אחרים. [כעניין של מינוח, נוקלאזות אקסו מנתקות רק נוקלאוטידים מ-DNA או RNA משני קצותיו, אך לא באמצע. נוקלאזות אנדו מבקעות קשרי פוספודיסטר הממוקמים עמוק יותר בתוך גדיל חומצת גרעין.] ליחידת המשנה θ אין פעילות אנזימטית והיא מווסתת את תפקוד האקסונוקלאז. למרות שיש לו פעילות פולימראז, לפולימראז הליבה של Pol III יש תהליכים גרועים - כלומר, הוא יכול להוסיף רק עד 15 נוקלאוטידים לפני שהוא מתנתק מה- DNA של התבנית. מכיוון שגנום של זני E. coli ממוצעים קרוב ל-5 מיליון זוגות בסיסים, שכפול במקטעים קטנים של 15 nt יהיה לא יעיל במיוחד.
קומפלקס מעמיס המהדק הוא מכלול ATPase הנקשר ליחידת β-clamp עם קישור ATP (אך פעילות ATPase אינה מופעלת). כאשר הקומפלקס נקשר לאחר מכן ל-DNA, הוא מפעיל את ה-ATPase, וההידרוליזה המתקבלת של ATP מובילה לשינויים קונפורמטיביים הפותחים את המהדק באופן זמני (כדי להקיף או להתרחק מגדיל ה-DNA), ולאחר מכן לניתוק של מעמיס המהדק ממכלול המהדק.
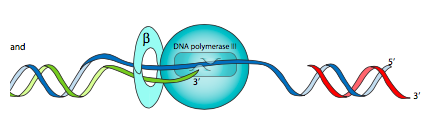
כאן יש צורך ביחידת המשנה β. ידוע גם בשם מהדק β, זהו דימר של יחידות משנה חצי עגולות שיש להן חור מרכזי דרכו מושחל ה- DNA. פולימראז הליבה, באמצעות אינטראקציה α-β, מחובר למהדק β זה כך שהוא יישאר על ה-DNA זמן רב יותר, מה שמגדיל את התהליכות של Pol III ליותר מ-5000 nt. מהדק β נטען על ה- DNA (ונפרק ממנו) על ידי קומפלקס מעמיס מהדק (הנקרא גם קומפלקס γ) המורכב מיחידות משנה γ (איקס3), δ, δ', χ ו- Ψ.
לבועת השכפול יש שני מזלגות שכפול - ברגע שה-DNA נפתח (לא רוכסן) במקור, יכולה להיווצר מכונת שכפול בכל קצה, כאשר המסוקים פונים לכיוונים מנוגדים. לשם הפישוט, נשקול רק מזלג אחד - פתיחה משמאל לימין - בדיון זה מתוך הבנה שאותו דבר קורה עם המזלג השני, אך בכיוון ההפוך.
הדבר הראשון שיש לשים לב אליו כאשר מסתכלים על תרשים של מזלג שכפול (איור\(\PageIndex{11}\)) הוא ששני החלקים החד-גדילים של ה- DNA של התבנית הם אנטי מקבילים. זה לא אמור להפתיע בשלב זה של הקורס, אבל זה כן מציג בעיה מכנית מעניינת. Helicase פותח את ה- DNA הכפול ומוביל את שאר מכונת השכפול. לכן, באזור החד-גדילי הנגרר אחר ההליקאז, אם נסתכל משמאל לימין, גדיל תבנית אחד הוא 3 'עד 5' (בכחול), ואילו השני הוא 5' עד 3' (באדום). מכיוון שאנו יודעים שחומצות גרעין מפולמרות על ידי הוספת פוספט 5' של נוקלאוטיד חדש להידרוקסיל 3' של הנוקלאוטיד הקודם (5' עד 3', בירוק), פירוש הדבר שאחד הגדילים, הנקרא הגדיל המוביל, מסונתז באותו כיוון שמכונת השכפול נעה. אין בעיה שם.
הגדיל השני הוא בעייתי: במבט ליניארי, הגדיל החדש שסונתז יעבור 3 'עד 5' משמאל לימין, אך פולימראזות DNA אינן יכולות להוסיף נוקלאוטידים כך. כיצד תאים פותרים בעיה זו? הוצעו מספר אפשרויות, אך המודל הנוכחי מתואר כאן. מכונת השכפול מורכבת מההליקאז, פרימאזות ושני הולואנזימים של DNA פולימראז III הנעים באותו כיוון פיזי (בעקבות ההליקאז). למעשה, מתחמי pol III מקושרים פיזית דרך יחידות משנה τ.
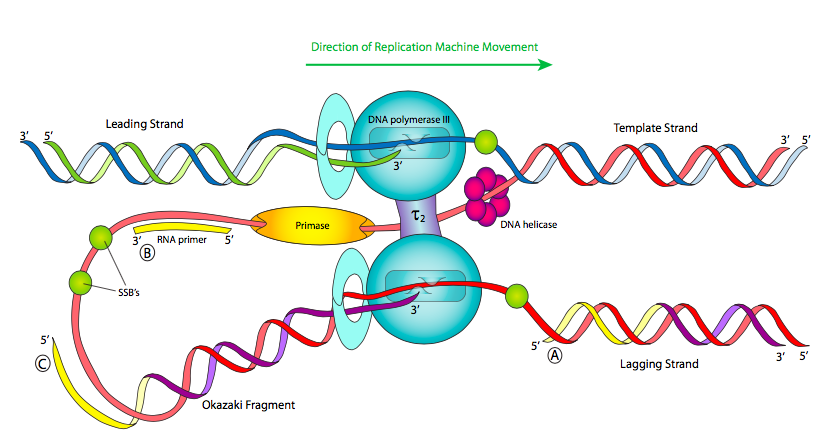
על מנת שכפל גדיל התבנית שהוא 5 'עד 3' משמאל לימין, יש להזין את הגדיל לפולימראז לאחור. ניתן להשיג זאת על ידי סיבוב הפולימראז או על ידי לולאה של ה- DNA מסביב. כפי שמראה האיור, המודל הנוכחי הוא שהפרימאז נע גם משמאל לימין, כך שיש לו רק זמן קצר לסנתז במהירות פריימר קצר לפני שיצטרך להתקדם עם הרפליזום ולהתחיל שוב, ולהשאיר פריימרים לסירוגין בעקבותיו. בגלל זה, Pol III נאלץ לסנתז רק שברים קצרים של הכרומוזום בכל פעם, הנקראים שברי Okazaki על שם מגלה שלהם. Pol III מתחיל לסנתז על ידי הוספת נוקלאוטידים לקצה 3' של פריימר וממשיך עד שהוא מגיע לקצה 5' של הפריימר הבא. זה לא (ולא יכול) לחבר את הגדיל שהוא מסנתז עם קצה הפריימר 5'.
שכפול DNA נקרא תהליך חצי לא רציף מכיוון שבעוד שהגדיל המוביל מסונתז ברציפות, הגדיל המפגר מסונתז בשברים. זה מוביל לשתי בעיות עיקריות: ראשית, נותרו פיסות קטנות של RNA בגדילים החדשים שנוצרו (רק בקצה 5' עבור הגדיל המוביל, במקומות רבים לפיגור); ושנית, Pol III יכול להוסיף רק נוקלאוטידים חופשיים לשבר של DNA חד גדילי; זה לא יכול לחבר שבר אחר. לכן, ה"גדיל "החדש אינו שלם, אלא רצוף בקשרי פוספודיסטר חסרים.
הבעיה הראשונה נפתרת על ידי DNA פולימראז I. שלא כמו Pol III, Pol I הוא חלבון מונומרי ופועל לבד, ללא חלבונים נוספים. יש גם פי 10-20 מולקולות Pol I מאשר מולקולות Pol III, מכיוון שהן נחוצות לכל כך הרבה שברי אוקזאקי. ל-DNA Polymerase I יש שלוש פעילויות: (1) כמו Pol III, הוא יכול לסנתז גדיל DNA המבוסס על תבנית DNA, (2) כמו Pol III, זהו אקסונוקלאז הגהה 3'-5', אך בניגוד ל-Pol III, (3) זהו גם אקסונוקלאז 5'-3'. פעילות האקסונוקלאז 5'-3' חיונית בהסרת פריימר ה-RNA (איור). \(\PageIndex{12}\) האקסונוקלאז 5'-3' נקשר ל-DNA דו-גדילי שיש לו שבירה חד-גדילית בעמוד השדרה הפוספודיאסטר כמו מה שקורה לאחר סינתזה של שברי אוקזאקי מפריימר אחד למשנהו, אך לא ניתן לחבר אותם. אקסונוקלאז זה 5'-3' מסיר לאחר מכן את פריימר ה- RNA. לאחר מכן, פעילות הפולימראז מוסיפה נוקלאוטידים DNA חדשים לשבר אוקאזאקי במעלה הזרם, וממלאת את הפער שנוצר על ידי הסרת פריימר ה- RNA. האקסונוקלאז ההגהה פועל בדיוק כמו שהוא עושה עבור Pol III, ומסיר מיד נוקלאוטיד שגוי ששולב לאחרונה. לאחר הגהה, שיעור השגיאות הכולל של שילוב נוקלאוטידים הוא בערך 1 מתוך 107.
מבחינה טכנית, האקסונוקלאז 5'-3' מבקע את ה-DNA באזור דו-גדילי במורד הזרם של הניק, ולאחר מכן עשוי להסיר מכל מקום בין 1-10nt בכל פעם. בניסוי, ניתן לפצל את פעילות האקסונוקלאז 5'-3' משאר Pol I על ידי טריפסין הפרוטאז. זה יוצר את "שבר קלנוב" המכיל את הפולימראז ו-3'-5' הגהה אקסונוקלאז.
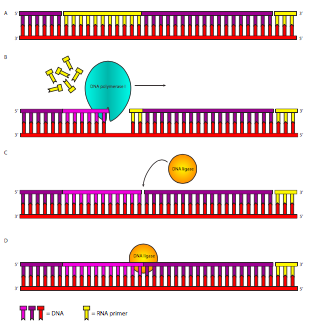
איור\(\PageIndex{12}\). סינתזת סטרנד בפיגור. לאחר ש- DNA פולימראז III האריך את הפריימרים (צהוב), DNA פולימראז I מסיר את הפריימר ומחליף אותו על ידי הוספה למקטע הקודם. כאשר הוא מסיים להסיר RNA ולהחליף אותו ב-DNA, הוא משאיר את ה-DNA עם קשר פוספודיסטר חסר בין ה-DNA המסונתז pol III במורד הזרם לבין ה-DNA המסונתז pol I במעלה הזרם. שבר זה בעמוד השדרה של סוכר-פוספט מתוקן על ידי ליגאז DNA.
למרות שה- RNA הוחלף ב- DNA, זה עדיין משאיר גדיל מקוטע. השחקן המרכזי האחרון בסיפור שכפול ה-DNA מופיע לבסוף: ליגאז DNA. לאנזים זה יש משימה אחת פשוטה אך מכריעה: הוא מזרז את ההתקפה של ה-3'-OH מקטע אחד על הפוספט 5' של השבר הבא, ויוצר קשר פוספודיסטר. תגובה זו דורשת אנרגיה בצורה של הידרוליזה של ATP או NAD + בהתאם למין (אי - קולי משתמש ב- NAD +) ויוצר AMP או PP i או NMN +.