
11.4: סינתזת חלבונים (תרגום)
- Page ID
- 208856

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מטרות למידה
- תאר את הקוד הגנטי והסבר מדוע הוא נחשב כמעט אוניברסלי
- הסבר את תהליך התרגום ואת הפונקציות של המנגנון המולקולרי של התרגום
- השווה תרגום באיקריוטים ופרוקריוטים
סינתזה של חלבונים צורכת יותר אנרגיה של התא מכל תהליך מטבולי אחר. בתורו, חלבונים מהווים יותר מסה מכל מקרומולקולה אחרת של אורגניזמים חיים. הם מבצעים כמעט כל פונקציה של תא, ומשמשים כאלמנטים פונקציונליים (למשל אנזימים) וגם כאלמנטים מבניים. תהליך התרגום, או סינתזת החלבון, החלק השני של ביטוי הגנים, כרוך בפענוח על ידי ריבוזום של הודעת mRNA למוצר פוליפפטיד.
הקוד הגנטי
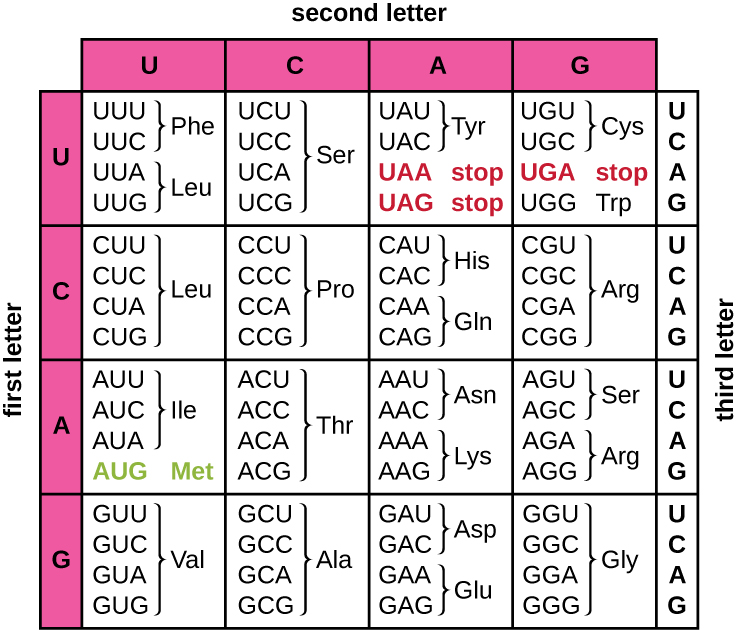
תרגום של תבנית ה-mRNA ממיר מידע גנטי מבוסס נוקלאוטידים ל"שפה" של חומצות אמינו ליצירת מוצר חלבון. רצף חלבונים מורכב מ-20 חומצות אמינו נפוצות. כל חומצת אמינו מוגדרת בתוך ה- mRNA על ידי שלישיית נוקלאוטידים הנקראת קודון. הקשר בין קודון mRNA לחומצת האמינו המתאימה לו נקרא הקוד הגנטי.
קוד שלושת הנוקלאוטידים אומר שיש בסך הכל 64 שילובים אפשריים (4 3, עם ארבעה נוקלאוטידים שונים אפשריים בכל אחד משלושת המיקומים השונים בתוך הקודון). מספר זה גדול ממספר חומצות האמינו וחומצת אמינו נתונה מקודדת על ידי יותר מקודון אחד (איור\(\PageIndex{1}\)). יתירות זו בקוד הגנטי נקראת ניוון. בדרך כלל, בעוד ששני המיקומים הראשונים בקודון חשובים לקביעת איזו חומצת אמינו תשולב בפוליפפטיד גדל, המיקום השלישי, הנקרא מיקום הנדנוד, הוא פחות קריטי. במקרים מסוימים, אם הנוקלאוטיד במיקום השלישי משתנה, אותה חומצת אמינו עדיין משולבת.
בעוד ש-61 מתוך 64 השלישיות האפשריות מקודדות לחומצות אמינו, שלושה מתוך 64 הקודונים אינם מקודדים לחומצת אמינו; הם מפסיקים את סינתזת החלבון ומשחררים את הפוליפפטיד ממכונת התרגום. אלה נקראים קודון עצור s או קודון שטויות s. לקודון נוסף, AUG, יש גם פונקציה מיוחדת. בנוסף לציון חומצת האמינו מתיונין, היא משמשת בדרך כלל גם כקודון ההתחלה ליזום תרגום. מסגרת הקריאה, האופן שבו נוקלאוטידים ב-mRNA מקובצים לקודונים, לתרגום נקבעת על ידי קודון ההתחלה של AUG ליד קצה 5' של ה-mRNA. כל קבוצה של שלושה נוקלאוטידים בעקבות קודון התחלה זה היא קודון בהודעת ה-mRNA.
הקוד הגנטי הוא כמעט אוניברסלי. למעט כמה יוצאים מן הכלל, כמעט כל המינים משתמשים באותו קוד גנטי לסינתזת חלבונים, וזו עדות חזקה לכך שכל החיים הקיימים על פני כדור הארץ חולקים מקור משותף. עם זאת, חומצות אמינו יוצאות דופן כגון סלנוציסטאין ופירוליסין נצפו בארכאה ובחיידקים. במקרה של סלנוציסטאין, הקודון המשמש הוא UGA (בדרך כלל קודון עצירה). עם זאת, UGA יכול לקודד לסלנוציסטאין באמצעות מבנה לולאת גזע (המכונה רצף החדרת סלנוציסטאין, או אלמנט SECIS), שנמצא באזור 3' הלא מתורגם של ה- mRNA. פירוליסין משתמש בקודון עצירה אחר, UAG. שילוב פירוליסין דורש את הגן pyls ו- RNA העברה ייחודי (tRNA) עם אנטיקודון CUA.
תרגיל \(\PageIndex{1}\)
- כמה בסיסים יש בכל קודון?
- לאיזו חומצת אמינו מקודדת על ידי הקודון AAU?
- מה קורה כאשר מגיעים לקודון עצירה?
מכונות סינתזת החלבון
בנוסף לתבנית ה- mRNA, מולקולות ומקרומולקולות רבות תורמות לתהליך התרגום. ההרכב של כל רכיב משתנה בין הטקסים; לדוגמה, ריבוזומים עשויים להיות מורכבים ממספרים שונים של RNAs ריבוזומליים (rRNA) ופוליפפטידים בהתאם לאורגניזם. עם זאת, המבנים והתפקודים הכלליים של מכונות סינתזת החלבון דומים מחיידקים לתאים אנושיים. תרגום דורש קלט של תבנית mRNA, ריבוזומים, tRNAs וגורמים אנזימטיים שונים.
ריבוזומים
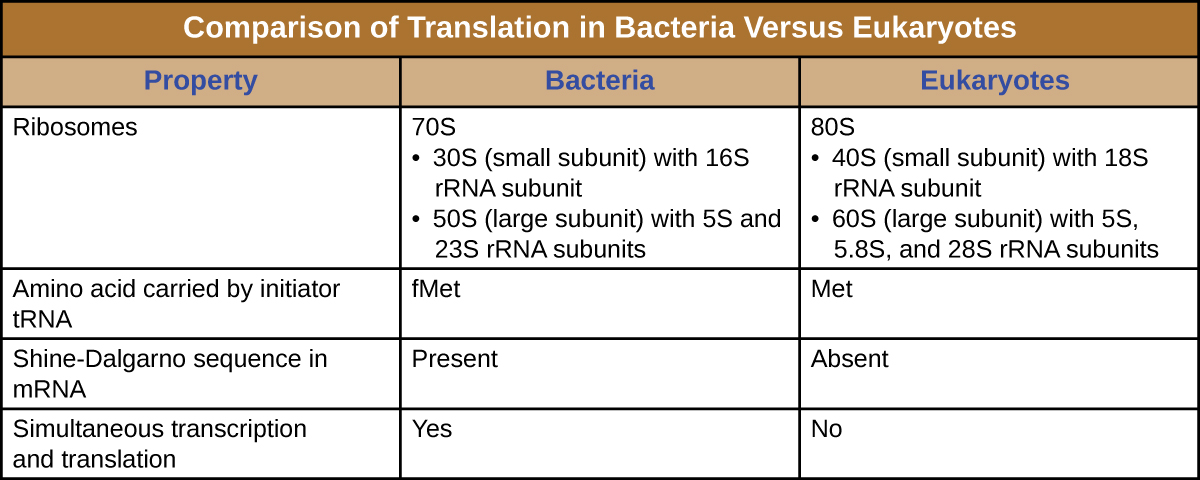
ריבוזום הוא מקרומולקולה מורכבת המורכבת מ- rRNA קטליטי (הנקרא ריבוזימים) ו- rRNA מבניים, כמו גם פוליפפטידים רבים ומובחנים. RRNAs בוגרים מהווים כ -50% מכל ריבוזום. לפרוקריוטים יש ריבוזומים 70S, בעוד שלאוקריוטים יש ריבוזומים 80S בציטופלזמה ורטיקולום אנדופלזמי מחוספס, וריבוזומים 70S במיטוכונדריה ובכלורופלסטים. ריבוזומים מתנתקים ליחידות משנה גדולות וקטנות כאשר הם אינם מסנתזים חלבונים ומתחברים מחדש במהלך תחילת התרגום. ב אי - קולי, יחידת המשנה הקטנה מתוארת כ- 30S (המכילה את יחידת המשנה 16S rRNA), ויחידת המשנה הגדולה היא 50S (המכילה את יחידות המשנה 5S ו- 23S rRNA), בסך הכל 70S (יחידות Svedberg אינן תוסף). לריבוזומים האוקריוטים יש תת-יחידה קטנה של 40S (המכילה את יחידת המשנה 18S rRNA) ויחידת משנה גדולה של 60S (המכילה את יחידות המשנה 5S, 5.8S ו- 28S rRNA), בסך הכל 80S. יחידת המשנה הקטנה אחראית לקשירת תבנית ה-mRNA, בעוד שתת-היחידה הגדולה קושרת tRNAs (נדון בסעיף המשנה הבא).
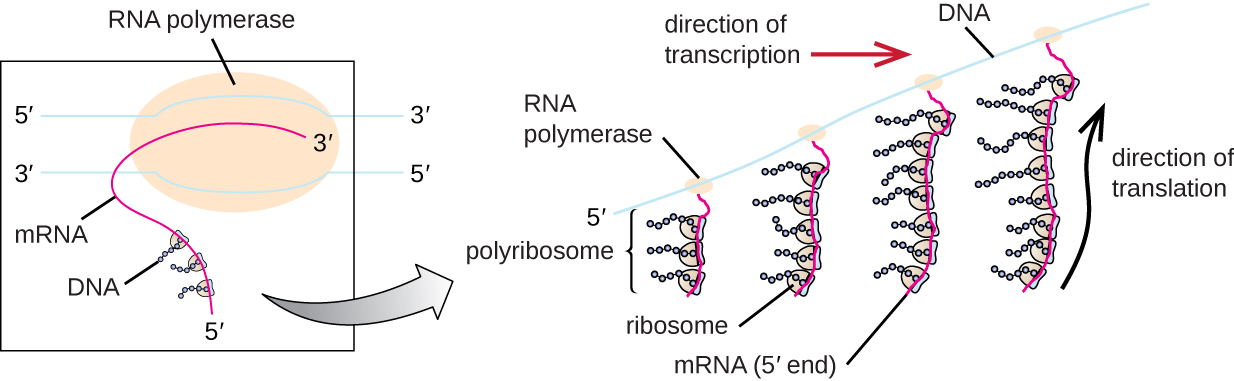
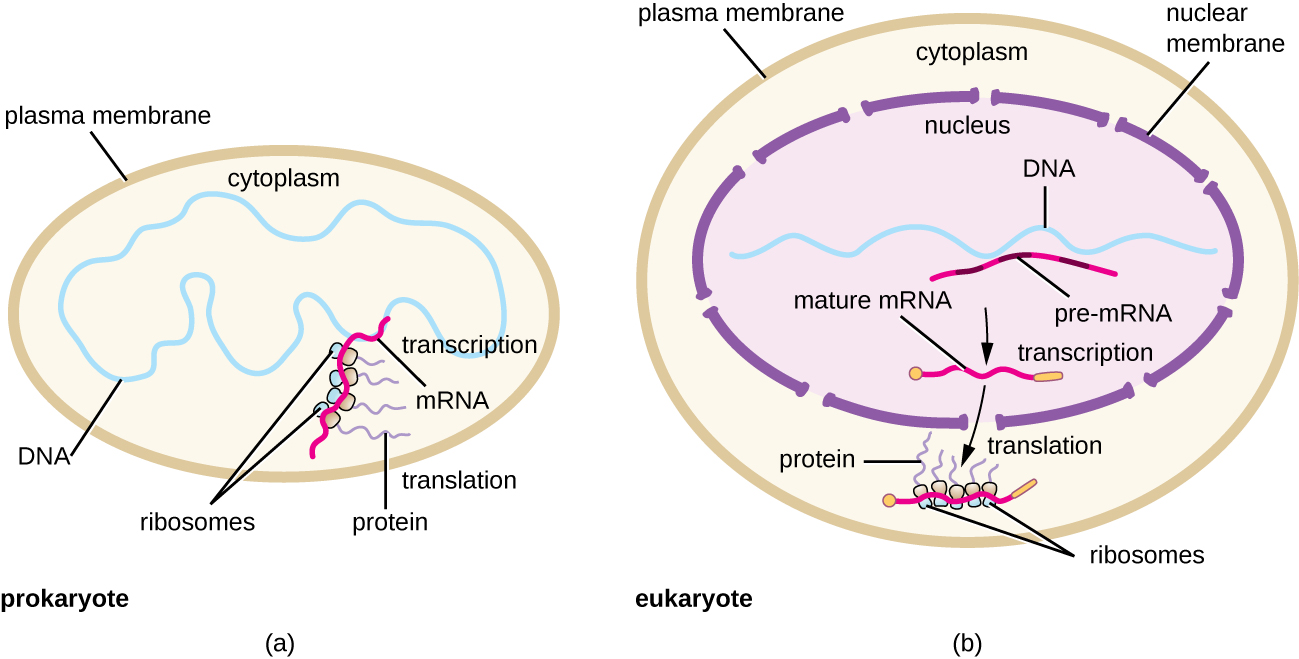
כל מולקולת mRNA מתורגמת בו זמנית על ידי ריבוזומים רבים, כולם מסנתזים חלבון באותו כיוון: קריאת ה-mRNA מ-5' עד 3' וסינתזה של הפוליפפטיד מקצה ה-N לקצה C. המבנה השלם המכיל mRNA עם ריבוזומים קשורים מרובים נקרא פוליריבוזום (או פוליזום). הן בחיידקים והן בארכיאה, לפני שמתרחשת הפסקת תעתיק, כל תמליל המקודד לחלבון כבר נמצא בשימוש כדי להתחיל בסינתזה של עותקים רבים של הפוליפפטידים המקודדים מכיוון שתהליכי השעתוק והתרגום יכולים להתרחש במקביל ויוצרים פוליריבוזומים (איור). \(\PageIndex{2}\) הסיבה לכך שתעתיק ותרגום יכולים להתרחש בו זמנית היא מכיוון ששני התהליכים הללו מתרחשים באותו כיוון 5' עד 3', שניהם מתרחשים בציטופלזמה של התא, ומכיוון שתעתיק ה-RNA אינו מעובד ברגע שהוא מתעתק. זה מאפשר לתא פרוקריוטי להגיב לאות סביבתי הדורש חלבונים חדשים במהירות רבה. לעומת זאת, בתאים אוקריוטיים, שעתוק ותרגום סימולטני אינם אפשריים. למרות שפוליריבוזומים נוצרים גם באיקריוטים, הם אינם יכולים לעשות זאת עד להשלמת סינתזת ה- RNA ומולקולת ה- RNA שונתה והועברה אל מחוץ לגרעין.
העברת RNAs
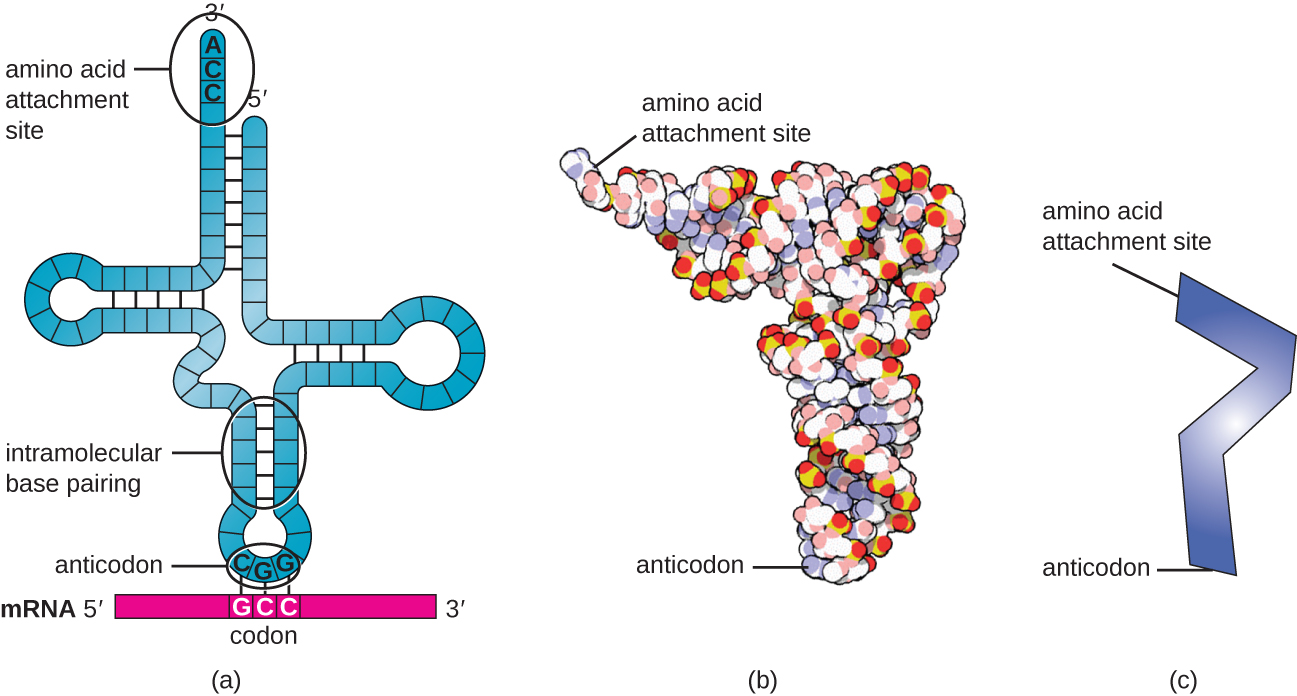
RNAs העברה (tRNAs) הם מולקולות RNA מבניות, ובהתאם למין, קיימים סוגים רבים ושונים של tRNA בציטופלזמה. למיני חיידקים יש בדרך כלל בין 60 ל-90 סוגים. משמש כמתאמים, כל סוג tRNA נקשר לקודון ספציפי בתבנית ה- mRNA ומוסיף את חומצת האמינו המתאימה לשרשרת הפוליפפטיד. לכן, tRNAs הן המולקולות שלמעשה "מתרגמות" את שפת ה- RNA לשפת החלבונים. כמולקולות המתאם של התרגום, מפתיע ש-tRNAs יכולים להתאים כל כך הרבה ספציפיות לחבילה כל כך קטנה. מולקולת ה- tRNA מקיימת אינטראקציה עם שלושה גורמים: סינתזות tRNA aminoacyl, ריבוזומים ו- mRNA.
tRNAs בוגרים מקבלים מבנה תלת מימדי כאשר בסיסים משלימים שנחשפים במולקולת ה- RNA החד-גדילית קשורים זה לזה (איור\(\PageIndex{3}\)). צורה זו ממקמת את אתר הקישור של חומצות אמינו, הנקרא קצה מחייב חומצת אמינו CCA, שהוא רצף ציטוזין-ציטוזין-אדנין בקצה 3' של ה- tRNA, והאנטי -קודונאט בקצה השני. האנטיקודון הוא רצף של שלושה נוקלאוטידים המתחבר לקודון mRNA באמצעות זיווג בסיסים משלים.
חומצת אמינו מתווספת לקצה מולקולת tRNA בתהליך של "טעינה" של tRNA, שבמהלכו כל מולקולת tRNA מקושרת לחומצת האמינו הנכונה או המוכרת שלה על ידי קבוצת אנזימים הנקראים aminoacyl tRNA synthetases. קיים לפחות סוג אחד של סינתטאז tRNA aminoacyl עבור כל אחת מ-20 חומצות האמינו. במהלך תהליך זה, חומצת האמינו מופעלת תחילה על ידי הוספת אדנוסין מונופוספט (AMP) ולאחר מכן מועברת ל- tRNA, מה שהופך אותה ל- tRNA טעון, ו- AMP משתחרר.
תרגיל \(\PageIndex{2}\)
- תאר את המבנה וההרכב של הריבוזום הפרוקריוטי.
- באיזה כיוון נקראת תבנית ה-mRNA?
- תאר את המבנה והתפקוד של tRNA.
מנגנון סינתזת החלבון
התרגום דומה בפרוקריוטים ובאוקריוטים. כאן נחקור כיצד מתרחש תרגום ב אי - קולי, פרוקריוט מייצג, ונציין כל הבדל בין תרגום חיידקי לאיקריוטי.
חניכה
התחלת סינתזת החלבון מתחילה ביצירת קומפלקס חניכה. ב אי - קולי, קומפלקס זה כולל את הריבוזום הקטן 30S, תבנית ה- mRNA, שלושה גורמי התחלה המסייעים לריבוזום להתאסף בצורה נכונה, גואנוסין טריפוספט (GTP) הפועל כמקור אנרגיה, ו- tRNA יוזם מיוחד הנושא N -formyl-methionine (fMet-tRNA fMet) (איור). \(\PageIndex{4}\) ה-tRNA היוזם מקיים אינטראקציה עם קודון ההתחלה AUG של ה-mRNA ונושא מתיונין פורמלי (fMet). בגלל מעורבותו בהתחלה, fMet מוכנס בתחילת (מסוף N) של כל שרשרת פוליפפטיד המסונתזת על ידי אי - קולי. ב אי - קולי mRNA, רצף מוביל במעלה הזרם של קודון AUG הראשון, הנקרא רצף Shine-Dalgarno (הידוע גם כאתר הקישור הריבוזומלי AGGAGG), מקיים אינטראקציה באמצעות זיווג בסיסים משלים עם מולקולות ה- rRNA המרכיבות את הריבוזום. אינטראקציה זו מעגנת את יחידת המשנה הריבוזומלית 30S במיקום הנכון בתבנית ה- mRNA. בשלב זה, תת-היחידה הריבוזומלית 50S נקשרת לאחר מכן לקומפלקס ההתחלה, ויוצרת ריבוזום שלם.
באיקריוטים, היווצרות קומפלקס חניכה דומה, עם ההבדלים הבאים:
- ה-tRNA היוזם הוא tRNA מיוחד אחר הנושא מתיונין, הנקרא Met-tRNAi
- במקום להיקשר ל-mRNA ברצף Shine-Dalgarno, קומפלקס ההתחלה האוקריוטי מזהה את מכסה ה-5' של ה-mRNA האיקריוטי, ואז עוקב לאורך ה-mRNA בכיוון 5' עד 3' עד לזיהוי קודון ההתחלה של AUG. בשלב זה, יחידת המשנה 60S נקשרת לקומפלקס של Met-tRNAi, mRNA ויחידת המשנה 40S.
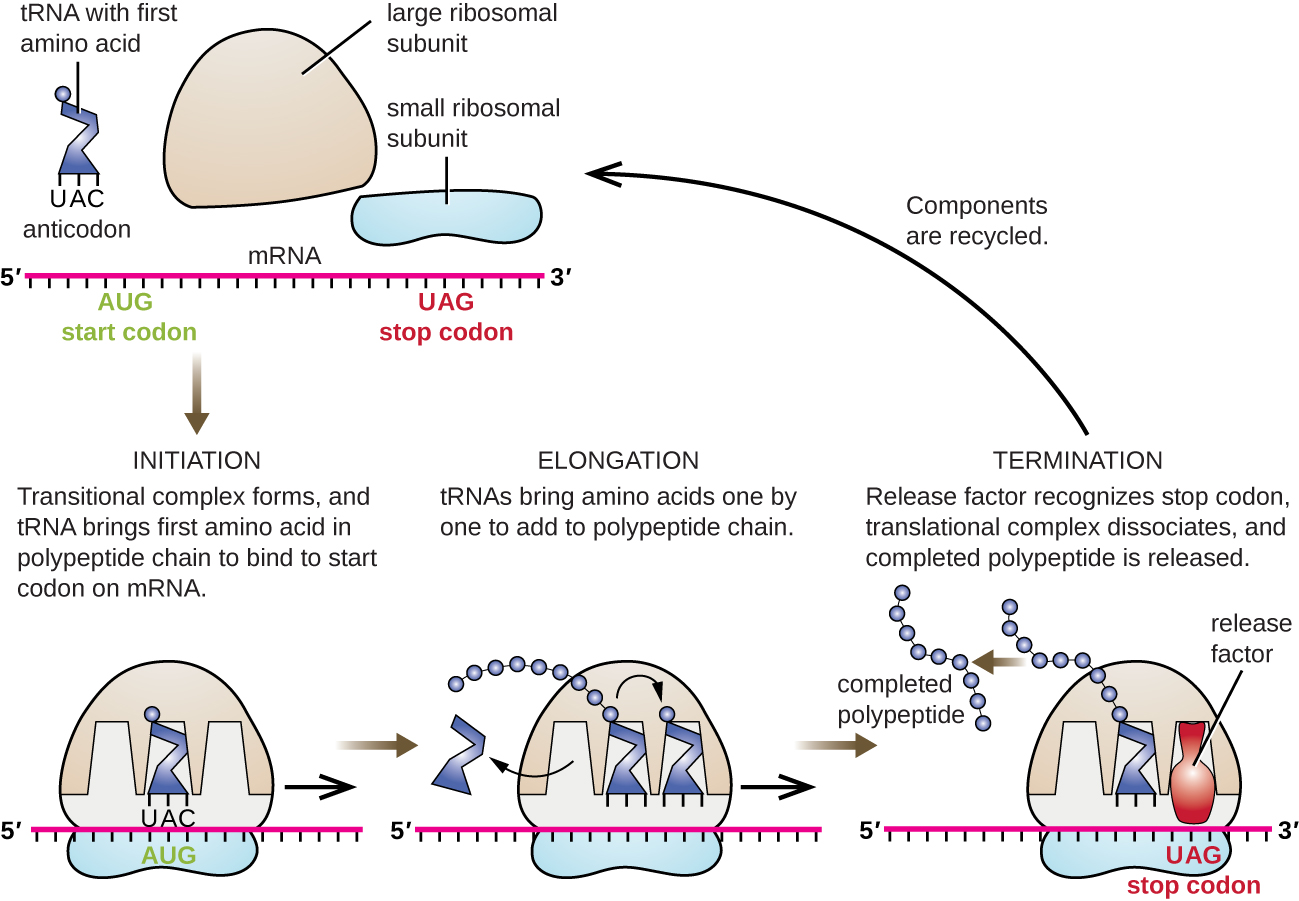
התארכות
בפרוקריוטים ובאוקריוטים, יסודות התארכות התרגום זהים. ב אי - קולי, הקישור של יחידת המשנה הריבוזומלית 50S לייצור הריבוזום השלם יוצר שלושה אתרים ריבוזומליים חשובים מבחינה תפקודית: אתר A (aminoacyl) קושר tRNAs aminoacyl טעונים נכנסים. אתר P (פפטידיל) קושר tRNAs טעונים הנושאים חומצות אמינו שיצרו קשרי פפטיד עם שרשרת הפוליפפטיד הגדלה אך עדיין לא התנתקו מה-tRNA המקביל שלהם. אתר E (יציאה) משחרר tRNAs מנותקים כך שניתן יהיה לטעון אותם מחדש עם חומצות אמינו חופשיות. יש חריג אחד בולט לקו הייצור הזה של tRNAs: במהלך היווצרות קומפלקס התחלה, fMET-tRNA fMet חיידקי או Met-tRNAi אוקריוטי נכנסים לאתר P ישירות מבלי להיכנס תחילה לאתר A, ומספקים אתר A בחינם מוכן לקבל את ה-tRNA המתאים לקודון הראשון לאחר ה-AUG.
ההתארכות ממשיכה בתנועות קודון יחיד של הריבוזום הנקראות כל אחת אירוע טרנסלוקציה. במהלך כל אירוע טרנסלוקציה, ה- tRNAs הטעונים נכנסים לאתר A, ואז עוברים לאתר P, ולאחר מכן לבסוף לאתר E להסרה. תנועות ריבוזומליות, או צעדים, נגרמות על ידי שינויים קונפורמטיביים המקדמים את הריבוזום בשלושה בסיסים בכיוון 3'. קשרי פפטיד נוצרים בין קבוצת האמינו של חומצת האמינו המחוברת ל- tRNA באתר A לבין קבוצת הקרבוקסיל של חומצת האמינו המחוברת ל- tRNA של אתר P. היווצרות כל קשר פפטיד מזורזת על ידי פפטידיל טרנספראז, ריבוזים מבוסס RNA המשולב ביחידת המשנה הריבוזומלית 50S. חומצת האמינו הקשורה ל- tRNA של אתר P קשורה גם לשרשרת הפוליפפטיד הגדלה. כאשר הריבוזום עובר על פני ה-mRNA, ה-tRNA לשעבר של אתר P נכנס לאתר E, מתנתק מחומצת האמינו וגורש. כמה מהשלבים במהלך התארכות, כולל קישור של tRNA aminoacyl טעון לאתר A וטרנסלוקציה, דורשים אנרגיה הנגזרת מהידרוליזה של GTP, המזרזת על ידי גורמי התארכות ספציפיים. למרבה הפלא, מנגנון התרגום E. coli לוקח רק 0.05 שניות להוסיף כל חומצת אמינו, כלומר ניתן לתרגם חלבון של 200 חומצות אמינו תוך 10 שניות בלבד.
סיום
סיום התרגום מתרחש כאשר נתקל בקודון שטויות (UAA, UAG או UGA) שאין לו tRNA משלים. ביישור עם אתר A, קודונים שטויות אלה מזוהים על ידי גורמי שחרור בפרוקריוטים ובאוקריוטים שגורמים לכך שחומצת האמינו באתר P מתנתקת מה-tRNA שלה, ומשחררת את הפוליפפטיד החדש שנוצר. יחידות המשנה הריבוזומליות הקטנות והגדולות מתנתקות מה- mRNA ומזו מזו; הם מגויסים כמעט מיד למתחם תרגום אחר.
לסיכום, ישנם מספר מאפיינים מרכזיים המבדילים בין ביטוי גנים פרוקריוטיים לזה שנראה באוקריוטים. אלה מומחשים באיור \(\PageIndex{5}\) ומופיעים באיור\(\PageIndex{6}\).
מיקוד, קיפול ושינוי חלבונים
במהלך התרגום ואחריו, ייתכן שיהיה צורך לשנות פוליפפטידים לפני שהם פעילים ביולוגית. שינויים שלאחר התרגום כוללים:
- הסרת רצפי אותות מתורגמים - זנבות קצרים של חומצות אמינו המסייעים בהכוונת חלבון לתא סלולרי ספציפי
- "קיפול" נכון של הפוליפפטיד והקשר של יחידות משנה מרובות של פוליפפטידים, לעתים קרובות מקלים על ידי חלבוני מלווה, למבנה תלת מימדי מובהק
- עיבוד פרוטאוליטי של פוליפפטיד לא פעיל לשחרור מרכיב חלבון פעיל, וכן
- שינויים כימיים שונים (למשל זרחון, מתילציה או גליקוזילציה) של חומצות אמינו בודדות.
תרגיל \(\PageIndex{3}\)
- מהם המרכיבים של מתחם החניכה לתרגום בפרוקריוטים?
- מהם שני הבדלים בין התחלת תרגום פרוקריוטי לאוקריוטי?
- מה קורה בכל אחד משלושת האתרים הפעילים של הריבוזום?
- מה גורם לסיום התרגום?
מושגי מפתח וסיכום
- בתרגום, פוליפפטידים מסונתזים באמצעות רצפי mRNA ומכונות סלולריות, כולל tRNAs התואמים קודונים mRNA לחומצות אמינו ספציפיות וריבוזומים המורכבים מ- RNA וחלבונים המזרזים את התגובה.
- הקוד הגנטי מנוון בכך שמספר קודוני mRNA מקודדים לאותן חומצות אמינו. הקוד הגנטי הוא כמעט אוניברסלי בקרב אורגניזמים חיים.
- ריבוזומים פרוקריוטיים (70S) וציטופלסמיים אוקריוטיים (80S) מורכבים כל אחד מיחידת משנה גדולה ויחידת משנה קטנה בגדלים שונים בין שתי הקבוצות. כל יחידת משנה מורכבת מ- rRNA וחלבון. ריבוזומים של אברון בתאים אוקריוטיים דומים לריבוזומים פרוקריוטיים.
- כ-60 עד 90 מינים של tRNA קיימים בחיידקים. לכל tRNA יש אנטיקודון בעל שלושה נוקלאוטידים וכן אתר קישור לחומצת אמינו קשורה. כל tRNAs עם אנטיקודון ספציפי ישאו את אותה חומצת אמינו.
- התחלת התרגום מתרחשת כאשר תת-היחידה הריבוזומלית הקטנה נקשרת עם גורמי התחלה ו-tRNA יוזם בקודון ההתחלה של mRNA, ואחריו הקישור למכלול ההתחלה של תת-היחידה הריבוזומלית הגדולה.
- בתאים פרוקריוטים, קודון ההתחלה מקודד ל- N-formyl-methionine הנישא על ידי tRNA יוזם מיוחד. בתאים אוקריוטיים, קודון ההתחלה מקודד למתיונין הנישא על ידי tRNA יוזם מיוחד. בנוסף, בעוד שהקישור הריבוזומלי של ה-mRNA בפרוקריוטים מתאפשר על ידי רצף Shine-Dalgarno בתוך ה-mRNA, ריבוזומים אוקריוטיים נקשרים למכסה 5' של ה-mRNA.
- במהלך שלב ההתארכות של התרגום, tRNA טעון נקשר ל-mRNA באתר A של הריבוזום; קשר פפטיד מזורז בין שתי חומצות האמינו הסמוכות, ושובר את הקשר בין חומצת האמינו הראשונה ל-tRNA שלה; הריבוזום מזיז קודון אחד לאורך ה-mRNA; וה-tRNA הראשון מועבר מאתר P של הריבוזום לאתר E ועוזב את הקומפלקס הריבוזומלי.
- סיום התרגום מתרחש כאשר הריבוזום נתקל בקודון עצירה, שאינו מקודד ל- tRNA. גורמי שחרור גורמים לשחרור הפוליפפטיד, והמתחם הריבוזומלי מתנתק.
- בפרוקריוטים, שעתוק ותרגום עשויים להיות משולבים, עם תרגום של מולקולת mRNA שמתחיל ברגע שהשעתוק מאפשר מספיק חשיפה ל-mRNA לקשירה של ריבוזום, לפני סיום השעתוק. שעתוק ותרגום אינם מצמידים באאוקריוטים מכיוון שתעתיק מתרחש בגרעין, ואילו תרגום מתרחש בציטופלזמה או בשיתוף עם הרשת האנדופלזמית המחוספסת.
- פוליפפטידים דורשים לעתים קרובות שינוי אחד או יותר לאחר התרגום כדי להפוך לפעילים ביולוגית.