
11.3: שעתוק RNA
- Page ID
- 208844

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מטרות למידה
- הסבר כיצד RNA מסונתז באמצעות DNA כתבנית
- להבחין בין שעתוק בפרוקריוטים לאיקריוטים
במהלך תהליך השעתוק, המידע המקודד בתוך רצף ה-DNA של גן אחד או יותר מועתק לגדיל של RNA, הנקרא גם תמליל RNA. מולקולת ה-RNA החד-גדילית המתקבלת, המורכבת מריבונוקלאוטידים המכילים את הבסיסים אדנין (A), ציטוסין (C), גואנין (G) ואורציל (U), פועלת כעותק מולקולרי נייד של רצף ה-DNA המקורי. שעתוק בפרוקריוטים ובאוקריוטים מחייב את הסליל הכפול של ה-DNA להירגע חלקית באזור סינתזת ה-RNA. האזור הלא מפותל נקרא בועת שעתוק. שעתוק של גן מסוים מתרחש תמיד מאחד משני גדילי ה-DNA הפועל כתבנית, מה שנקרא גדיל אנטי-סנס. תוצר ה- RNA משלים לגדיל התבנית של ה- DNA והוא כמעט זהה לגדיל ה- DNA הלא תבנית, או לגדיל החוש. ההבדל היחיד הוא שב- RNA, כל נוקלאוטידים T מוחלפים בנוקלאוטידים U; במהלך סינתזת RNA, U משולב כאשר יש A בגדיל האנטי-סנס המשלים.
תמלול בחיידקים
חיידקים משתמשים באותו RNA פולימראז כדי לתמלל את כל הגנים שלהם. בדומה לפולימראז DNA, RNA פולימראז מוסיף נוקלאוטידים בזה אחר זה לקבוצת 3'-OH של שרשרת הנוקלאוטידים הגדלה. הבדל קריטי אחד בפעילות בין DNA פולימראז ל-RNA פולימראז הוא הדרישה ל-3'-OH שעליו להוסיף נוקלאוטידים: פולימראז DNA דורש קבוצת 3'-OH כזו, ובכך מחייב פריימר, בעוד ש-RNA פולימראז לא. במהלך השעתוק, ריבונוקלאוטיד המשלים לגדיל תבנית ה-DNA מתווסף לגדיל ה-RNA הגדל וקשר פוספודיסטר קוולנטי נוצר על ידי סינתזת התייבשות בין הנוקלאוטיד החדש לזה האחרון שנוסף. ב אי - קולי, RNA פולימראז כולל שש יחידות משנה של פוליפפטיד, חמש מהן מרכיבות את אנזים הליבה של הפולימראז האחראי להוספת נוקלאוטידים של RNA לגדיל גדל. יחידת המשנה השישית ידועה בשם sigma (σ). גורם σ מאפשר ל-RNA פולימראז להיקשר למקדם ספציפי, ובכך מאפשר שעתוק של גנים שונים. ישנם גורמי σ שונים המאפשרים שעתוק של גנים שונים.
חניכה
התחלת השעתוק מתחילה בפרומוטור, רצף DNA שאליו נקשרת מנגנון השעתוק ומתחיל שעתוק. זוג הנוקלאוטידים בסליל הכפול של ה-DNA המתאים לאתר שממנו מתועתק נוקלאוטיד ה-RNA הראשון 5' הוא אתר ההתחלה. נוקלאוטידים שקדמו לאתר ההתחלה נקראים "במעלה הזרם", ואילו נוקלאוטידים העוקבים אחר אתר ההתחלה נקראים נוקלאוטידים "במורד הזרם". ברוב המקרים, היזמים ממוקמים ממש במעלה הזרם של הגנים שהם מווסתים. למרות שרצפי האמרגן משתנים בין הגנום החיידקי, כמה אלמנטים נשמרים. בעמדות -10 ו-35 בתוך ה-DNA לפני אתר ההתחלה (מסומן +1), ישנם שני רצפי קונצנזוס פרומטור, או אזורים דומים בכל המקדמים ועל פני מיני חיידקים שונים. רצף הקונצנזוס —10, הנקרא תיבת TATA, הוא TATAAT. רצף —35 מזוהה ומחויב על ידי σ.
התארכות
ההתארכות בשלב השעתוק מתחילה כאשר תת-היחידה σ מתנתקת מהפולימראז, ומאפשרת לאנזים הליבה לסנתז RNA המשלים לתבנית ה-DNA בכיוון 5' עד 3' בקצב של כ-40 נוקלאוטידים בשנייה. ככל שההתארכות מתקדמת, ה-DNA מתפרק ברציפות לפני אנזים הליבה ומתפתל מאחוריו (איור). \(\PageIndex{1}\)
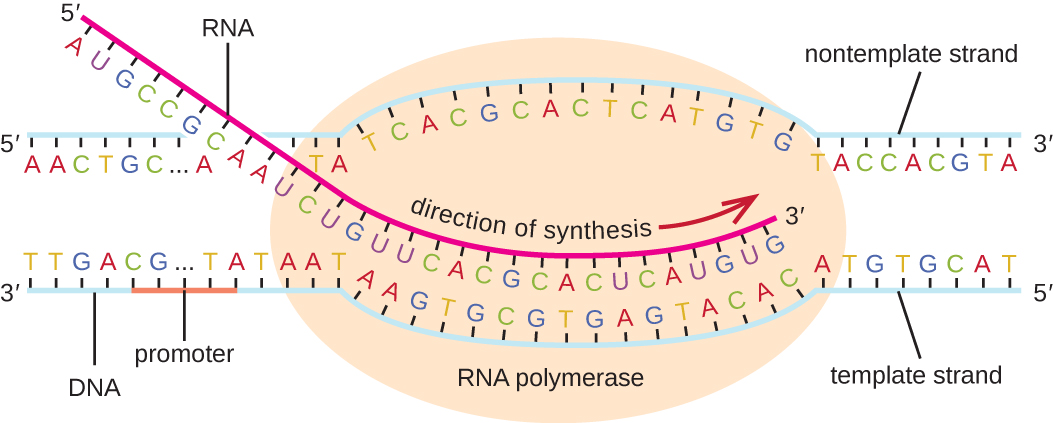
סיום
לאחר שעתוק גן, הפולימראז החיידקי חייב להתנתק מתבנית ה-DNA ולשחרר את ה-RNA החדש שנוצר. זה מכונה סיום התעתיק. תבנית ה-DNA כוללת רצפי נוקלאוטידים חוזרים ונשנים הפועלים כאותות סיום, וגורמים ל-RNA פולימראז להיעצר ולהשתחרר מתבנית ה-DNA, ומשחררים את תמליל ה-RNA.
תרגיל \(\PageIndex{1}\)
- היכן גורם σ של RNA פולימראז קושר DNA כדי להתחיל בשעתוק?
- מה קורה כדי ליזום את פעילות הפילמור של RNA פולימראז?
- מאיפה מגיע האות לסיום התעתיק?
תמלול באוקריוטים
פרוקריוטים ואיקריוטים מבצעים ביסודו את אותו תהליך שעתוק, עם כמה הבדלים משמעותיים (ראה טבלה\(\PageIndex{1}\)). אוקריוטים משתמשים בשלושה פולימראזות שונות, פולימראזות RNA I, II ו- III, כולם נבדלים מבחינה מבנית מפולימראז ה- RNA החיידקי. כל אחד מתמלל תת-קבוצה אחרת של גנים. מעניין לציין כי ארכאים מכילים פולימראז RNA יחיד הקשור יותר ל-RNA פולימראז II אוקריוטי מאשר למקבילו החיידקי. mRNA אוקריוטי הם בדרך כלל גם מונוציסטרוניים, כלומר כל אחד מהם מקודד רק לפוליפפטיד בודד, בעוד ש-mRNA פרוקריוטיים של חיידקים וארכאים הם בדרך כלל פוליציסטרוניים, כלומר הם מקודדים לפוליפפטידים מרובים.
ההבדל החשוב ביותר בין פרוקריוטים לאיקריוטים הוא הגרעין הקשור לממברנה של האחרון, המשפיע על קלות השימוש במולקולות RNA לסינתזת חלבונים. כאשר הגנים קשורים בגרעין, התא האוקריוטי חייב להעביר מולקולות RNA המקודדות חלבון לציטופלזמה לתרגום. תמלילים ראשוניים המקודדים לחלבון, מולקולות ה-RNA המסונתזות ישירות על ידי RNA פולימראז, חייבות לעבור מספר שלבי עיבוד כדי להגן על מולקולות ה-RNA הללו מפני פירוק במהלך הזמן שהן מועברות מהגרעין לציטופלזמה ומתורגמות לחלבון. לדוגמה, mRNA אוקריוטי עשוי להימשך מספר שעות, בעוד שה-mRNA הפרוקריוטי הטיפוסי נמשך לא יותר מ-5 שניות.
התמליל העיקרי (נקרא גם pre-mRNA) מצופה תחילה בחלבונים מייצבי RNA כדי להגן עליו מפני פירוק בזמן שהוא מעובד ומיוצא מהגרעין. סוג העיבוד הראשון מתחיל בזמן שהתמליל הראשוני עדיין מסונתז; נוקלאוטיד מיוחד של 7-מתילגואנוסין, הנקרא מכסה 5', מתווסף לקצה 5' של התמליל הגדל. בנוסף למניעת פירוק, גורמים המעורבים בסינתזת החלבון שלאחר מכן מזהים את המכסה, המסייע ליזום תרגום על ידי ריבוזומים. לאחר השלמת ההתארכות, אנזים עיבוד אחר מוסיף מחרוזת של כ-200 נוקלאוטידים אדנין לקצה 3', הנקרא זנב פולי-A. שינוי זה מגן עוד יותר על הפרה-mRNA מפני השפלה ומאותת לגורמים סלולריים שיש לייצא את התמליל לציטופלזמה.
גנים אוקריוטיים המקודדים לפוליפפטידים מורכבים מרצפי קידוד הנקראים אקסונים (ex-on מסמל שהם לחוצים לשעבר) ורצפים מתערבים הנקראים אינטרונים (int -ron מציין את תפקידם המתמשך). רצפי RNA מתועתקים המתאימים לאינטרונים אינם מקודדים לאזורים של הפוליפפטיד הפונקציונלי ומוסרים מהפרה-mRNA במהלך העיבוד. חיוני שכל רצפי ה-RNA המקודדים לאינטרון יוסרו לחלוטין ומדויק מ-pre-mRNA לפני סינתזת החלבון, כך שרצפי ה-RNA המקודדים לאקסון מחוברים כראוי כדי לקודד פוליפפטיד פונקציונלי. אם התהליך טועה אפילו על ידי נוקלאוטיד בודד, הרצפים של האקסונים המחוברים מחדש יוסטו, והפוליפפטיד המתקבל לא יהיה מתפקד. תהליך הסרת רצפי RNA המקודדים לאינטרון וחיבור מחדש של אלה המקודדים על ידי אקסונים נקרא שחבור RNA והוא מתאפשר על ידי פעולה של ספליזוזום המכיל חלבוני ריבונוקליאו גרעיניים קטנים (snRNPs). רצפי RNA המקודדים לאינטרון מוסרים מה- pre-mRNA בזמן שהוא עדיין בגרעין. למרות שהם אינם מתורגמים, נראה כי לאינטרונים יש פונקציות שונות, כולל ויסות גנים והובלת mRNA. עם השלמת השינויים הללו, התמליל הבוגר, ה-mRNA המקודד לפוליפפטיד, מועבר אל מחוץ לגרעין, המיועד לציטופלזמה לתרגום. ניתן לחבר אינטרונים בצורה שונה, וכתוצאה מכך אקסונים שונים נכללים או לא נכללים במוצר ה-mRNA הסופי. תהליך זה ידוע בשם שחבור חלופי. היתרון של שחבור חלופי הוא שניתן ליצור סוגים שונים של תמלילי mRNA, כולם נגזרים מאותו רצף DNA. בשנים האחרונות הוכח שלחלק מהארכאים יש גם את היכולת לחבר את הפרה-mRNA שלהם.
| רכוש | חיידקים | אוקריוטים |
|---|---|---|
| מספר הפוליפפטידים המקודדים לכל mRNA | מונוציסטרוני או פוליציסטרוני | מונוציסטרוני בלעדי |
| התארכות סטרנד | ליבה+ σ = הולואנזים | פולימראזות RNA I, II או III |
| תוספת של כובע 5 ' | לא | הן |
| תוספת של זנב פולי-A בגודל 3 אינץ' | לא | הן |
| שחבור של פרה-mRNA | לא | הן |
דמיינו כיצד שחבור mRNA מתרחש על ידי צפייה בתהליך בפעולה בסרטון זה. ראה כיצד אינטרונים מוסרים במהלך שחבור RNA כאן.
תרגיל \(\PageIndex{2}\)
- בתאים אוקריוטיים, כיצד משתנה תמליל ה- RNA מגן לחלבון לאחר תעתוקו?
- האם אקסונים או אינטרונים מכילים מידע לרצפי חלבון?
מיקוד קליני: חלק 2
במחלקת החירום סיפרה אחות למארק כי קיבל החלטה טובה להגיע לבית החולים מכיוון שהתסמינים שלו הצביעו על זיהום שיצא משליטה. הסימפטומים של מארק התקדמו, כאשר אזור העור מושפע וכמות הנפיחות גדלה. בתוך האזור הפגוע החלה פריחה, שלפוחיות וכיסי גז קטנים מתחת לשכבת העור החיצונית ביותר נוצרו, וחלק מהעור הפך לאפור. בהתבסס על הריח המחריד של מוגלה המתנקזת מאחת השלפוחיות, ההתקדמות המהירה של הזיהום והמראה החזותי של העור הפגוע, החל הרופא מיד בטיפול בפאסיטיס נמק. הרופא של מארק הורה על תרבית של הנוזל המתנקז מהשלפוחית וגם הורה על בדיקת דם, כולל ספירת תאי דם לבנים.
מארק אושפז ביחידה לטיפול נמרץ והחל במתן תוך ורידי של אנטיביוטיקה רחבת טווח כדי לנסות למזער התפשטות נוספת של הזיהום. למרות הטיפול האנטיביוטי, מצבו של מארק הידרדר במהירות. מארק נעשה מבולבל ומסוחרר. תוך שעות ספורות מאשפוזו בבית החולים, לחץ הדם שלו ירד משמעותית ונשימתו נעשתה רדודה ומהירה יותר. בנוסף, שלפוחיות גברו, כאשר השלפוחיות התעצמו בצבע לשחור סגול, ונראה שהפצע עצמו מתקדם במהירות במעלה רגלו של מארק.
תרגיל \(\PageIndex{3}\)
- מהם הגורמים הסיבתיים האפשריים לדלקת המעי הגס של מארק?
- מהם כמה הסברים אפשריים מדוע נראה כי הטיפול האנטיביוטי אינו פועל?
מושגי מפתח וסיכום
- במהלך התעתיק, המידע המקודד ב- DNA משמש לייצור RNA.
- RNA פולימראז מסנתז RNA, תוך שימוש בגדיל האנטי-סנס של ה-DNA כתבנית על ידי הוספת נוקלאוטידים משלימים של RNA לקצה 3' של הגדיל הגדל.
- RNA פולימראז נקשר ל- DNA ברצף הנקרא מקדם במהלך תחילת השעתוק.
- גנים המקודדים לחלבונים בעלי פונקציות קשורות מתועתקים לעתים קרובות בשליטת מקדם יחיד בפרוקריוטים, וכתוצאה מכך נוצרת מולקולת mRNA פוליציסטרונית המקודדת לפוליפפטידים מרובים.
- שלא כמו פולימראז DNA, RNA פולימראז אינו דורש קבוצת 3'-OH כדי להוסיף נוקלאוטידים, ולכן אין צורך בפריימר במהלך ההתחלה.
- הפסקת השעתוק בחיידקים מתרחשת כאשר פולימראז ה- RNA נתקל ברצפי DNA ספציפיים המובילים לעצירת הפולימראז. התוצאה היא שחרור של RNA פולימראז מחוט תבנית ה-DNA, ומשחרר את תמליל ה-RNA.
- לאוקריוטים יש שלוש פולימראזות RNA שונות. לאיקריוטים יש גם mRNA מונוציסטרוני, שכל אחד מהם מקודד לפוליפפטיד בודד בלבד.
- תמלילים ראשוניים אוקריוטיים מעובדים במספר דרכים, כולל תוספת של מכסה 5' וזנב 3′- פולי-A, כמו גם שחבור, ליצירת מולקולת mRNA בוגרת הניתנת להובלה אל מחוץ לגרעין ומוגנת מפני השפלה.