
11.2: שכפול DNA
- Page ID
- 208845

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מטרות למידה
- הסבר את המשמעות של שכפול DNA שמרני למחצה
- הסבר מדוע שכפול ה- DNA הוא דו כיווני וכולל גם גדיל מוביל וגם בפיגור
- הסבר מדוע נוצרים שברי אוקזאקי
- תאר את תהליך שכפול ה- DNA ואת תפקודי האנזימים המעורבים
- זהה את ההבדלים בין שכפול DNA בחיידקים ואיקריוטים
- הסבר את תהליך שכפול המעגל המתגלגל
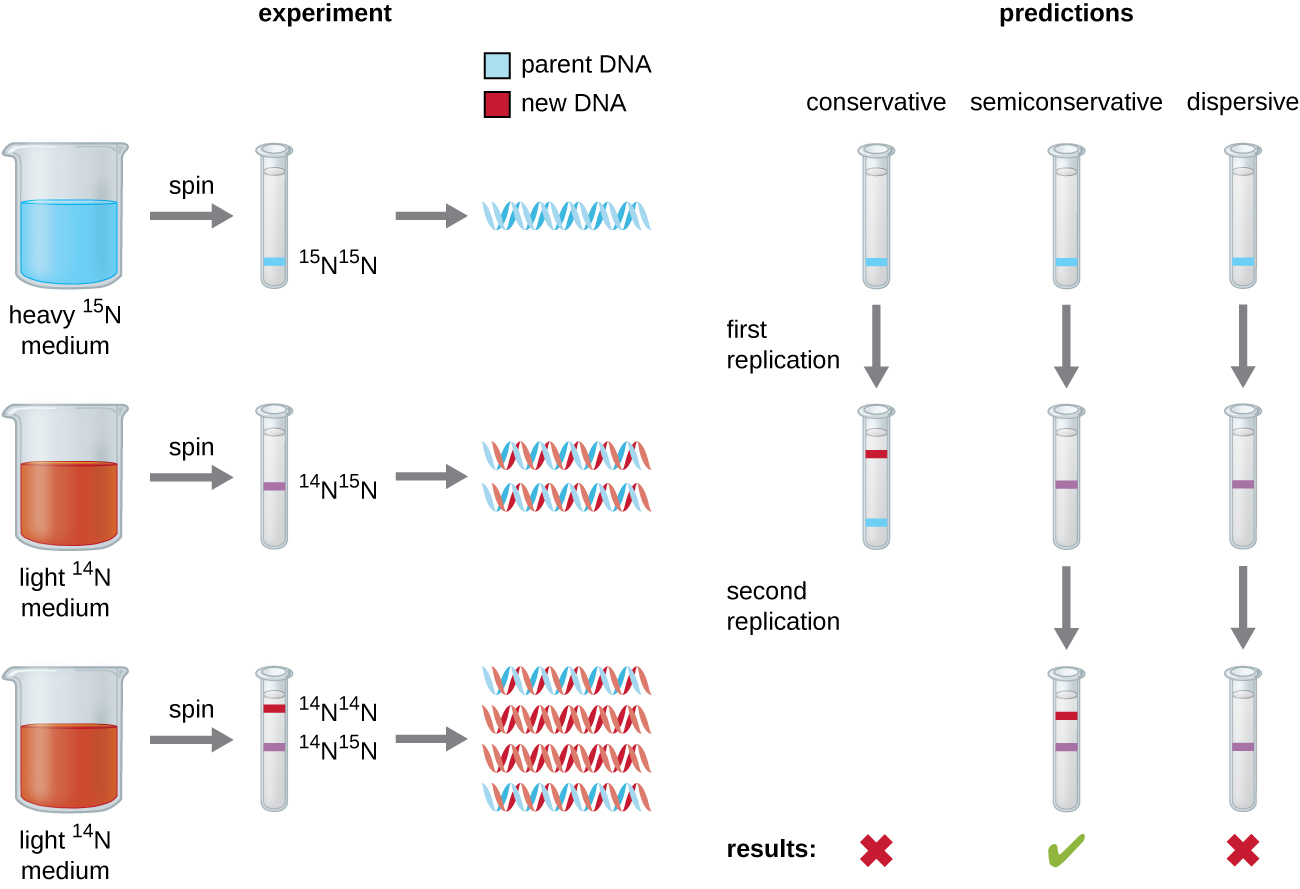
הבהרת מבנה הסליל הכפול על ידי ג'יימס ווטסון ופרנסיס קריק בשנת 1953 סיפקה רמז לאופן העתקת ה- DNA במהלך תהליך השכפול. הפרדת גדילי הסליל הכפול תספק שתי תבניות לסינתזה של גדילים משלימים חדשים, אך עדיין לא ברור כיצד בדיוק נבנו מולקולות DNA חדשות. במודל אחד, שכפול חצי שמרני, שני גדילי הסליל הכפול נפרדים במהלך שכפול ה- DNA, וכל גדיל משמש כתבנית שממנה מועתק הגדיל המשלים החדש; לאחר השכפול, כל DNA דו-גדילי כולל גדיל הורי או "ישן" אחד וגדיל "חדש" אחד. היו גם שני מודלים מתחרים שהוצעו: שמרני ופיזור, המוצגים באיור\(\PageIndex{1}\).

מתיו מסלסון (1930—) ופרנקלין סטאל (1929—) המציאו ניסוי בשנת 1958 כדי לבדוק איזה מהמודלים הללו מייצג נכון את שכפול ה-DNA (איור). \(\PageIndex{2}\) הם גידלו אי - קולי במשך כמה דורות במדיום המכיל איזוטופ "כבד" של חנקן (15 N) ששולב בבסיסים חנקניים ובסופו של דבר ב- DNA. זה תייג את ה- DNA ההורי. תרבות E. coli הועברה לאחר מכן למדיום המכיל 14 N והורשה לגדול במשך דור אחד. התאים נקצרו וה- DNA היה מבודד. ה- DNA הופרד על ידי אולטרה-צנטריפוגה, שבמהלכה יצר ה- DNA להקות בהתאם לצפיפותו. DNA שגדל ב-15 N צפוי ליצור רצועה במיקום צפיפות גבוה יותר מזה שגדל ב-14 נ 'Meselson ו-Stahl ציינו כי לאחר דור אחד של צמיחה ב-14 N, הלהקה הבודדת שנצפתה הייתה בינונית במיקום בין ה-DNA של תאים שגדלו אך ורק ב-15 N או 14 N. זה הציע אופן שכפול חצי שמרני או מפוזר. חלק מהתאים הורשו לגדול עוד דור אחד ב-14 N והסתובבו שוב. ה- DNA שנקטף מתאים שגדלו במשך שני דורות ב- 14 N יצר שתי להקות: רצועת DNA אחת הייתה במיקום הביניים בין 15 N ל- 14 N, והשנייה התאימה לרצועה של 14 N DNA. ניתן להסביר תוצאות אלו רק אם ה- DNA משתכפל בצורה שמרנית למחצה. לכן, שני הדגמים האחרים נשללו. כתוצאה מניסוי זה, אנו יודעים כעת כי במהלך שכפול ה- DNA, כל אחד משני הגדילים המרכיבים את הסליל הכפול משמש כתבנית שממנה מועתקים גדילים חדשים. הגדיל החדש יהיה משלים לחוט ההורי או "הישן". למולקולות ה-DNA המתקבלות יש אותו רצף והן מחולקות באופן שווה לשני תאי הבת.
תרגיל \(\PageIndex{1}\)
מה הייתה המסקנה של הניסוי של מסלסון וסטאל אם, אחרי הדור הראשון, הם היו מוצאים שתי להקות DNA?
שכפול DNA בחיידקים
שכפול ה- DNA נחקר היטב בחיידקים בעיקר בגלל גודלו הקטן של הגנום והמוטנטים הזמינים. אי - קולי יש 4.6 מיליון זוגות בסיסים (Mbp) בכרומוזום מעגלי יחיד וכולו משוכפל תוך כ-42 דקות, החל ממקור שכפול יחיד וממשיך סביב המעגל דו כיווני (כלומר, בשני הכיוונים). משמעות הדבר היא כי כ 1000 נוקלאוטידים מתווספים לשנייה. התהליך מהיר למדי ומתרחש עם מעט שגיאות.
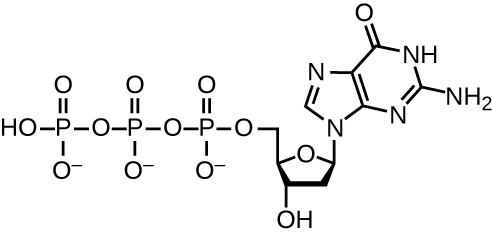
שכפול DNA משתמש במספר רב של חלבונים ואנזימים (טבלה\(\PageIndex{1}\)). אחד השחקנים המרכזיים הוא האנזים DNA פולימראז, המכונה גם DNA pol. בחיידקים ידועים שלושה סוגים עיקריים של פולימראזות DNA: DNA pol I, DNA pol II ו- DNA pol III. כיום ידוע כי DNA pol III הוא האנזים הנדרש לסינתזת DNA; DNA pol I ו- DNA pol II נדרשים בעיקר לתיקון. DNA pol III מוסיף דאוקסיריבונוקלאוטידים שכל אחד מהם משלים לנוקלאוטיד על גדיל התבנית, אחד אחד לקבוצת 3'-OH של שרשרת ה-DNA הגדלה. תוספת של נוקליאוטידים אלה דורשת אנרגיה. אנרגיה זו קיימת בקשרים של שלוש קבוצות פוספט המחוברות לכל נוקלאוטיד (נוקלאוטיד טריפוספט), בדומה לאופן שבו האנרגיה מאוחסנת בקשרי הפוספט של אדנוסין טריפוספט (ATP) (איור). \(\PageIndex{3}\) כאשר הקשר בין הפוספטים נשבר ודיפוספט משתחרר, האנרגיה המשתחררת מאפשרת יצירת קשר פוספודיסטר קוולנטי על ידי סינתזת התייבשות בין הנוקלאוטיד הנכנס לקבוצת 3'-OH החופשית על גדיל ה-DNA הגדל.
חניכה
תחילת השכפול מתרחשת ברצף נוקלאוטידים ספציפי הנקרא מקור השכפול, כאשר חלבונים שונים נקשרים כדי להתחיל בתהליך השכפול. אי - קולי יש מקור שכפול יחיד (כמו רוב הפרוקריוטים), הנקרא oriC, על הכרומוזום האחד שלו. מקור השכפול הוא באורך של כ-245 זוגות בסיסים והוא עשיר ברצפי אדנין-תימין (AT).
חלק מהחלבונים הנקשרים למקור השכפול חשובים בהנגשת אזורים חד-גדיליים של DNA לשכפול. DNA כרומוזומלי עטוף בדרך כלל סביב היסטונים (באאוקריוטים ובארכיאה) או חלבונים דמויי היסטון (בחיידקים), והוא מפותל, או עטוף ומפותל בהרחבה על עצמו. אריזה זו הופכת את המידע במולקולת ה- DNA לבלתי נגיש. עם זאת, אנזימים הנקראים טופואיזומראזות משנים את הצורה ואת סליל העל של הכרומוזום. כדי להתחיל שכפול DNA חיידקי, הכרומוזום המפותל נרגע על ידי טופואיזומראז II, הנקרא גם DNA gyrase. אנזים הנקרא helicase מפריד אז את גדילי ה-DNA על ידי שבירת קשרי המימן בין זוגות הבסיס החנקניים. נזכיר שלרצפי AT יש פחות קשרי מימן, ומכאן שיש להם אינטראקציות חלשות יותר מרצפי גואנין-ציטוסין (GC). אנזימים אלה דורשים הידרוליזה של ATP. כאשר ה- DNA נפתח נוצרים מבנים בצורת Y הנקראים מזלגות שכפול. שני מזלגות שכפול נוצרים במקור השכפול, המאפשרים שכפול דו כיווני ויצירת מבנה שנראה כמו בועה במבט במיקרוסקופ אלקטרוני שידור; כתוצאה מכך, מבנה זה נקרא בועת שכפול. ה-DNA ליד כל מזלג שכפול מצופה בחלבונים קושרים חד-גדיליים כדי למנוע מה-DNA החד-גדילי להתפתל מחדש לסליל כפול.
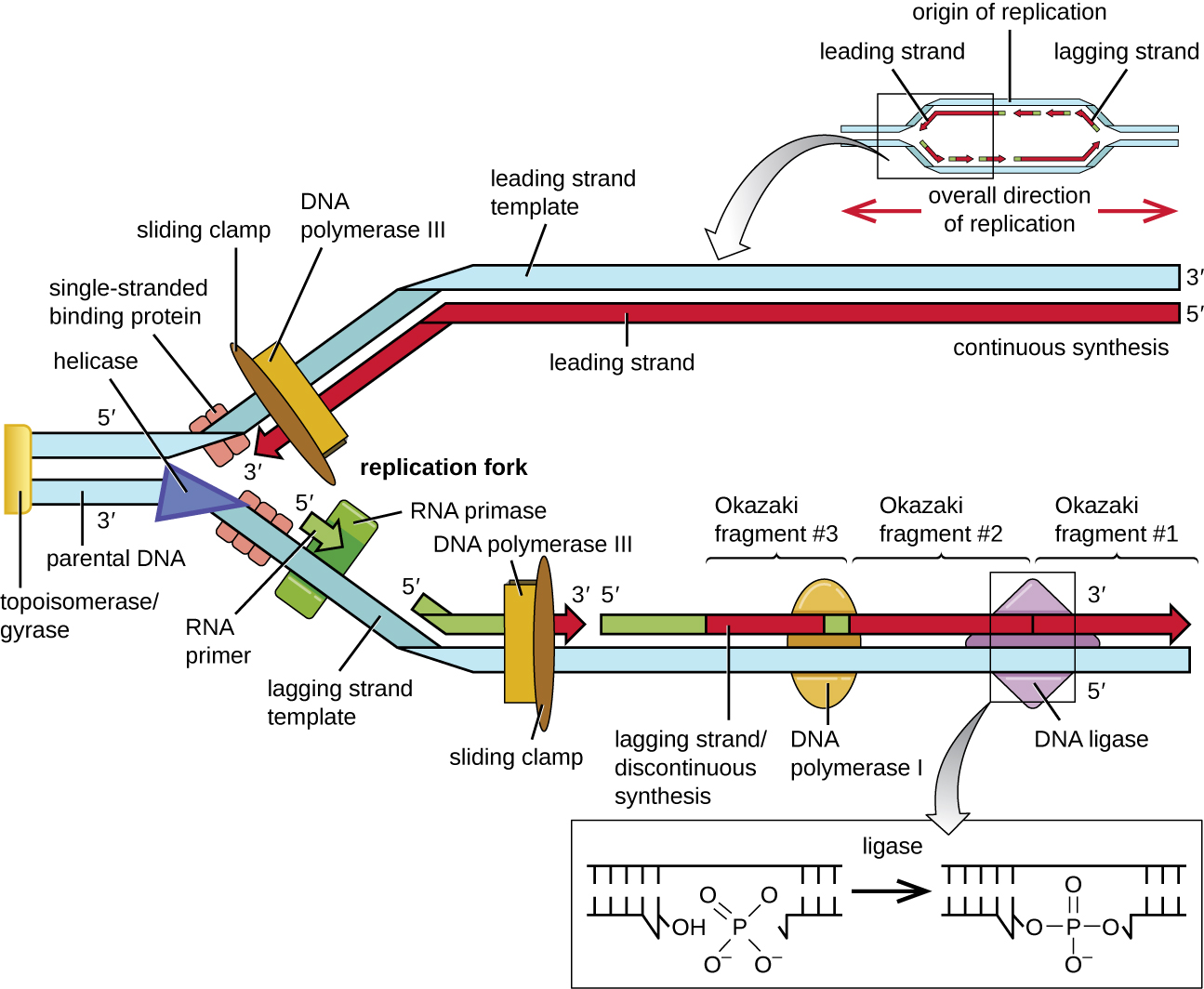
ברגע ש- DNA חד גדילי נגיש במקור השכפול, שכפול ה- DNA יכול להתחיל. עם זאת, DNA pol III מסוגל להוסיף נוקלאוטידים רק בכיוון 5' עד 3' (ניתן להרחיב גדיל DNA חדש רק בכיוון זה). הסיבה לכך היא ש-DNA פולימראז דורש קבוצת 3'-OH חופשית שאליה הוא יכול להוסיף נוקלאוטידים על ידי יצירת קשר פוספודיסטר קוולנטי בין קצה 3'-OH לפוספט 5' של הנוקלאוטיד הבא. זה גם אומר שהוא לא יכול להוסיף נוקלאוטידים אם קבוצת 3'-OH חופשית אינה זמינה, וזה המקרה של גדיל בודד של DNA. הבעיה נפתרת בעזרת רצף RNA המספק את קצה 3'-OH החופשי. מכיוון שרצף זה מאפשר את תחילת סינתזת ה- DNA, הוא נקרא כראוי פריימר. אורכו של הפריימר הוא חמישה עד 10 נוקלאוטידים ומשלים ל-DNA ההורי או התבנית. הוא מסונתז על ידי RNA פרימאז, שהוא פולימראז RNA. שלא כמו פולימראזות DNA, פולימראזות RNA אינן זקוקות לקבוצת 3'-OH חופשית כדי לסנתז מולקולת RNA. כעת, כשהפריימר מספק את קבוצת 3'-OH החופשית, DNA פולימראז III יכול כעת להרחיב את פריימר ה- RNA הזה, ולהוסיף נוקלאוטידים של DNA בזה אחר זה המשלימים לגדיל התבנית (איור). \(\PageIndex{1}\)
התארכות
במהלך התארכות בשכפול ה- DNA, הוספת נוקלאוטידים מתרחשת בקצב המרבי שלה של כ -1000 נוקלאוטידים בשנייה. DNA פולימראז III יכול להתרחב רק בכיוון 5' עד 3', מה שמציב בעיה במזלג השכפול. הסליל הכפול של ה- DNA הוא אנטי מקביל; כלומר, גדיל אחד מכוון בכיוון 5 'עד 3' והשני מכוון לכיוון 3' עד 5' (ראה מבנה ותפקוד ה- DNA). במהלך השכפול, גדיל אחד, המשלים את גדיל ה-DNA ההורי 3' עד 5', מסונתז ברציפות לכיוון מזלג השכפול מכיוון שפולימראז יכול להוסיף נוקלאוטידים בכיוון זה. גדיל מסונתז ברציפות זה ידוע כגדיל המוביל. הגדיל השני, המשלים ל-DNA ההורי 5' עד 3', צומח הרחק ממזלג השכפול, ולכן הפולימראז חייב לנוע חזרה לכיוון מזלג השכפול כדי להתחיל להוסיף בסיסים לפריימר חדש, שוב בכיוון הרחק ממזלג השכפול. הוא עושה זאת עד שהוא נתקל בגדיל שסונתז בעבר ואז הוא נע שוב אחורה (איור\(\PageIndex{4}\)). שלבים אלה מייצרים שברי רצף DNA קטנים המכונים שברי Okazaki, כל אחד מופרד על ידי פריימר RNA. שברי אוקזאקי נקראים על שם צוות המחקר היפני והזוג הנשוי רייג'י וצונקו אוקזאקי, שגילו אותם לראשונה בשנת 1966. הגדיל עם שברי אוקאזאקי ידוע כגדיל הפיגור, ואומרים שהסינתזה שלו אינה רציפה.
ניתן להרחיב את הגדיל המוביל מפריימר אחד בלבד, ואילו הגדיל המפגר זקוק לפריימר חדש לכל אחד משברי האוקזאקי הקצרים. הכיוון הכללי של הגדיל המפגר יהיה 3' עד 5', וזה של הגדיל המוביל 5' עד 3'. חלבון הנקרא מהדק הזזה מחזיק את פולימראז ה- DNA במקומו כשהוא ממשיך להוסיף נוקלאוטידים. מהדק הזזה הוא חלבון בצורת טבעת הנקשר ל-DNA ומחזיק את הפולימראז במקומו. מעבר לתפקידו בהתחלה, טופואיזומראז מונע גם את סלילה יתר של הסליל הכפול של ה- DNA לפני מזלג השכפול כאשר ה- DNA נפתח; הוא עושה זאת על ידי גרימת חתכים זמניים בסליל ה- DNA ולאחר מכן אטימתו מחדש. ככל שהסינתזה מתקדמת, הפריימרים של ה- RNA מוחלפים ב- DNA. הפריימרים מוסרים על ידי פעילות האקסונוקלאז של DNA פולימראז I, והפערים מתמלאים. החתכים שנותרו בין ה-DNA החדש שסונתז (שהחליף את פריימר ה-RNA) לבין ה-DNA שסונתז בעבר נאטמים על ידי האנזים DNA ligase המזרז את היווצרות קישור פוספודיסטר קוולנטי בין קצה 3'-OH של שבר DNA אחד לקצה הפוספט 5' של השבר השני, מייצב את עמוד השדרה של הסוכר-פוספט של מולקולת ה-DNA.
סיום
לאחר שכפול הכרומוזום השלם, הפסקת שכפול ה- DNA חייבת להתרחש. למרות שידוע הרבה על התחלת שכפול, פחות ידוע על תהליך הסיום. לאחר השכפול, הגנום המעגלי השלם המתקבל של פרוקריוטים משורשר, כלומר כרומוזומי ה-DNA המעגליים משולבים וחייבים להיות מופרדים זה מזה. זה מושג באמצעות פעילות של טופואיזומראז IV חיידקי, המציג הפסקות דו-גדיליות למולקולות DNA, ומאפשר להן להיפרד זו מזו; לאחר מכן האנזים אוטם מחדש את הכרומוזומים המעגליים. הרזולוציה של קונקטמרים היא סוגיה ייחודית לשכפול ה- DNA הפרוקריוטי בגלל הכרומוזומים המעגליים שלהם. מכיוון שגם DNA חיידקי gyrase וגם topoisomerase IV נבדלים מעמיתיהם האוקריוטיים, אנזימים אלה משמשים כמטרות לסוג של תרופות אנטי-מיקרוביאליות הנקראות קווינולונים.
| אנזים או גורם | פונקציה |
|---|---|
| DNA פול I | פעילות אקסונוקלאז מסירה פריימר RNA ומחליפה אותו ב- DNA שסונתז לאחרונה |
| פול DNA III | אנזים ראשי שמוסיף נוקלאוטידים בכיוון 5' עד 3' |
| הליקאז | פותח את סליל ה- DNA על ידי שבירת קשרי מימן בין הבסיסים החנקניים |
| ליגאז | אוטם את הפערים בין שברי Okazaki על הגדיל בפיגור כדי ליצור גדיל DNA רציף אחד |
| פרימייס | מסנתז פריימרים של RNA הדרושים כדי להתחיל בשכפול |
| חלבונים מחייבים חד-גדיליים | נקשר ל-DNA חד-גדילי כדי למנוע קשר מימן בין גדילי DNA, רפורמה ב-DNA דו-גדילי |
| מהדק הזזה | עוזר להחזיק את ה- DNA pol III במקומו כאשר מוסיפים נוקלאוטידים |
| טופואיזומראז II (DNA gyrase) | מרגיע כרומוזום מפותל כדי להפוך את ה- DNA לנגיש יותר לתחילת השכפול; עוזר להקל על הלחץ על ה- DNA בעת התנתקות, על ידי גרימת הפסקות ואז איטום מחדש של ה- DNA |
| טופואיזומראז IV | מציג פריצה חד-גדילית לכרומוזומים משורשרים כדי לשחרר אותם זה מזה, ולאחר מכן אוטם מחדש את ה-DNA |
תרגיל \(\PageIndex{2}\)
- איזה אנזים שובר את קשרי המימן המחזיקים את שני גדילי ה-DNA יחד כך שיכול להתרחש שכפול?
- האם הגדיל המפגר או הגדיל המוביל מסונתז בכיוון לכיוון פתיחת מזלג השכפול?
- איזה אנזים אחראי להסרת הפריימרים של ה-RNA ב-DNA חיידקי ששוכפל לאחרונה?
שכפול DNA באיקריוטים
גנומים אוקריוטיים הם הרבה יותר מורכבים וגדולים מגנום פרוקריוטי והם מורכבים בדרך כלל מכרומוזומים ליניאריים מרובים (טבלה\(\PageIndex{2}\)). לגנום האנושי, למשל, יש 3 מיליארד זוגות בסיסים לכל קבוצה הפלואידית של כרומוזומים, ו-6 מיליארד זוגות בסיסים מוכנסים במהלך השכפול. ישנם מקורות שכפול מרובים בכל כרומוזום אוקריוטי (איור\(\PageIndex{5}\)); לגנום האנושי יש 30,000 עד 50,000 מקורות שכפול. קצב השכפול הוא כ-100 נוקלאוטידים בשנייה - פי 10 איטי יותר משכפול פרוקריוטי.
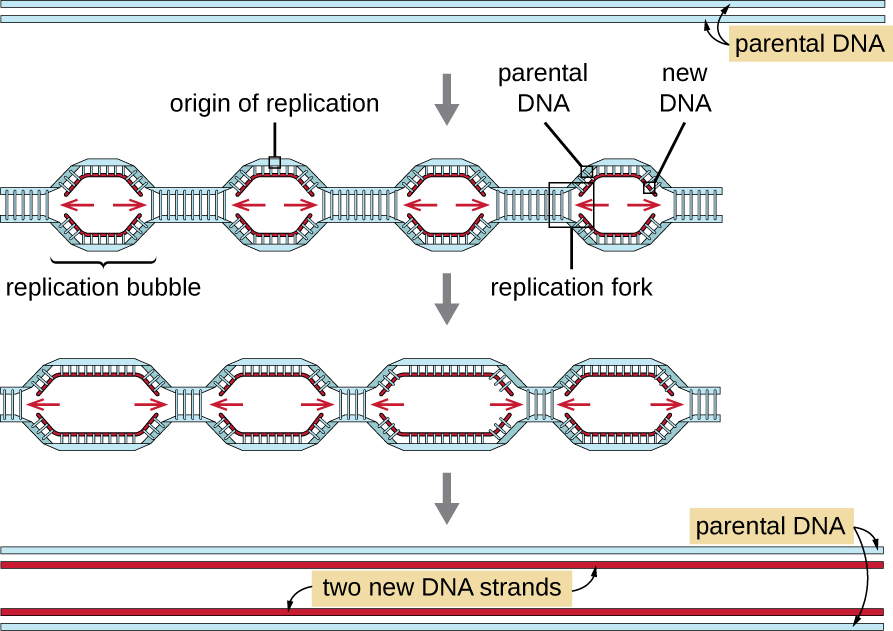
השלבים החיוניים של שכפול באאוקריוטים זהים לפרוקריוטים. לפני שהשכפול יכול להתחיל, ה-DNA צריך להיות זמין כתבנית. ה- DNA האוקריוטי מפותל וארוז מאוד, מה שמקל על ידי חלבונים רבים, כולל היסטונים (ראה מבנה ותפקוד הגנום הסלולרי). במקור השכפול, קומפלקס טרום-שכפול המורכב ממספר חלבונים, כולל הליקאז, יוצר ומגייס אנזימים אחרים המעורבים בהתחלת השכפול, כולל טופואיזומראז להרפיית סליל-על, חלבון מחייב חד-גדילי, RNA פרימאז ופולימראז DNA. לאחר תחילת השכפול, בתהליך דומה לזה שנמצא בפרוקריוטים, התארכות מתאפשרת על ידי פולימראזות DNA אוקריוטיות. הגדיל המוביל מסונתז ברציפות על ידי האנזים הפולימראז האוקריוטי pol δ, בעוד שהגדיל המפגר מסונתז על ידי pol ε. חלבון מהדק מחליק מחזיק את פולימראז ה- DNA במקומו כך שהוא לא ייפול מה- DNA. האנזים ריבונוקלאז H (RNase H), במקום פולימראז DNA כמו בחיידקים, מסיר את פריימר ה- RNA, אשר מוחלף לאחר מכן בנוקלאוטידים של DNA. הפערים שנותרו אטומים על ידי ליגאז DNA.
מכיוון שכרומוזומים אוקריוטיים הם ליניאריים, אפשר לצפות שהשכפול שלהם יהיה פשוט יותר. כמו בפרוקריוטים, פולימראז ה- DNA האוקריוטי יכול להוסיף נוקלאוטידים רק בכיוון 5' עד 3'. בגדיל המוביל הסינתזה נמשכת עד שהיא מגיעה לקצה הכרומוזום או למזלג שכפול אחר המתקדם בכיוון ההפוך. על הגדיל הפיגור, DNA מסונתז במתיחות קצרות, שכל אחת מהן יוזמת על ידי פריימר נפרד. כאשר מזלג השכפול מגיע לקצה הכרומוזום הליניארי, אין מקום ליצור פריימר לשבר ה-DNA שיועתק בסוף הכרומוזום. קצוות אלה נותרים אפוא ללא זיווג ועם הזמן הם עשויים להתקצר בהדרגה ככל שהתאים ממשיכים להתחלק.
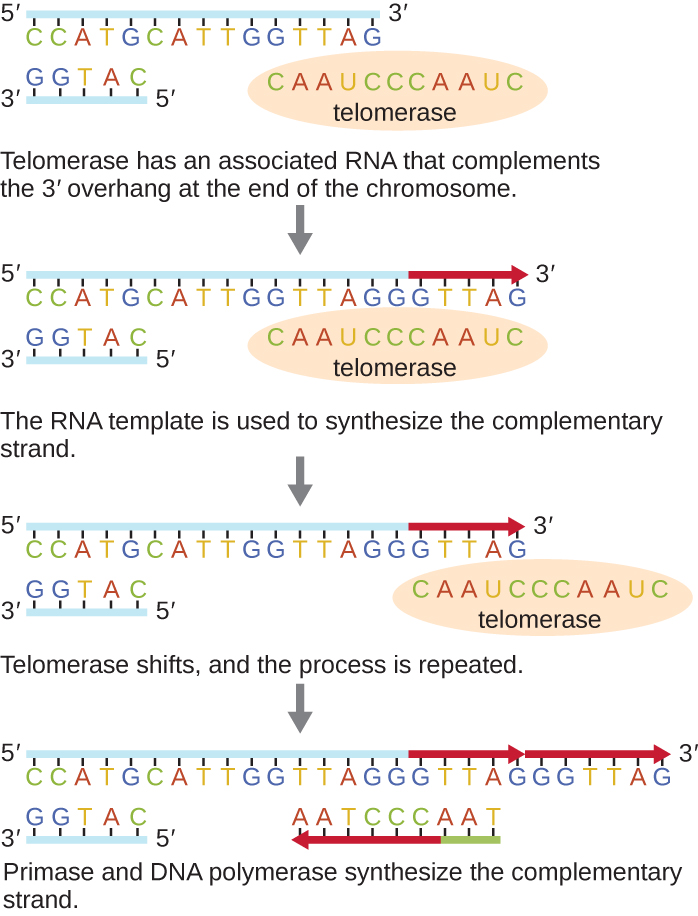
קצות הכרומוזומים הליניאריים ידועים בשם טלומרים ומורכבים מרצפים חוזרים שאינם מקודדים. הטלומרים מגנים על רצפי קידוד מפני אובדן כאשר התאים ממשיכים להתחלק. בבני אדם, רצף של שישה זוגות בסיסים, TTAGGG, חוזר על עצמו 100 עד 1000 פעמים ליצירת הטלומרים. גילוי האנזים טלומראז (איור\(\PageIndex{6}\)) הבהיר את הבנתנו כיצד נשמרים קצוות הכרומוזומים. טלומראז מכיל חלק קטליטי ותבנית RNA מובנית. הוא מתחבר לקצה הכרומוזום, ובסיסים משלימים לתבנית ה-RNA מתווספים בקצה 3' של גדיל ה-DNA. ברגע שקצה 3' של תבנית הגדיל המפגר מוארך מספיק, פולימראז DNA יכול להוסיף את הנוקלאוטידים המשלימים לקצוות הכרומוזומים. באופן זה משוכפלים קצות הכרומוזומים. בבני אדם, טלומראז פעיל בדרך כלל בתאי נבט ובתאי גזע בוגרים; הוא אינו פעיל בתאים סומטיים בוגרים ועשוי להיות קשור להזדקנות של תאים אלה. חיידקים אוקריוטיים כולל פטריות ופרוטוזואנים מייצרים גם טלומראז לשמירה על שלמות כרומוזומלית. על גילוי הטלומראז ופעולתו, אליזבת בלקברן (1948—) קיבלה את פרס נובל לרפואה או פיזיולוגיה בשנת 2009.
| רכוש | חיידקים | אוקריוטים |
|---|---|---|
| מבנה הגנום | כרומוזום מעגלי יחיד | כרומוזומים ליניאריים מרובים |
| מספר המקורות לכרומוזום | רווק | מרובה |
| קצב השכפול | 1000 נוקלאוטידים בשנייה | 100 נוקלאוטידים בשנייה |
| טלומראז | לא נוכח | הווה |
| הסרת פריימר RNA | DNA פול I | RNase H |
| התארכות סטרנד | פול DNA III | פול δ, פול ε |
תרגיל \(\PageIndex{3}\)
- במה שונה מקור השכפול בין אוקריוטים לפרוקריוטים?
- אילו אנזימי פולימראז אחראים לסינתזת ה- DNA במהלך שכפול אוקריוטי?
- מה נמצא בקצות הכרומוזומים באיקריוטים ומדוע?
שכפול DNA של אלמנטים אקסטרכרומוזומליים: פלסמידים ווירוסים
כדי להעתיק את חומצות הגרעין שלהם, פלסמידים ווירוסים משתמשים לעתים קרובות בווריאציות על דפוס שכפול ה-DNA המתואר עבור גנומים פרוקריוטים. למידע נוסף על המגוון הרחב של אסטרטגיות שכפול ויראלי, ראה מחזור החיים הנגיפי.
שכפול מעגל מתגלגל
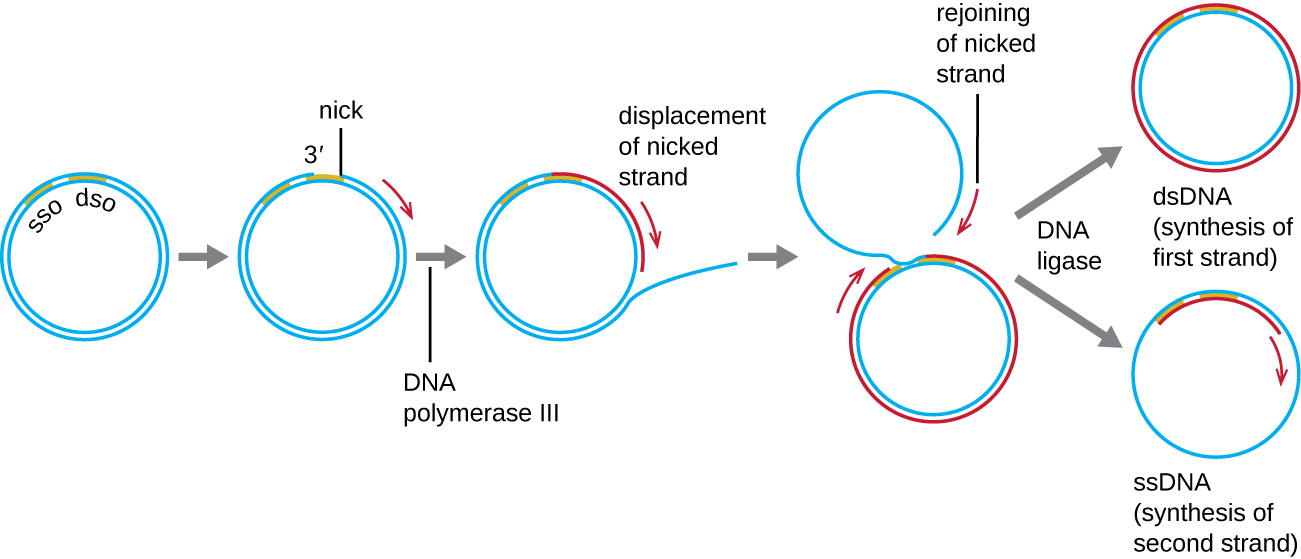
בעוד שפלסמידים חיידקיים רבים (ראה מאפיינים ייחודיים של תאים פרוקריוטים) משתכפלים בתהליך דומה לזה המשמש להעתקת הכרומוזום החיידקי, פלסמידים אחרים, מספר בקטריופאגים וכמה וירוסים של אוקריוטים משתמשים בשכפול מעגל מתגלגל (איור). \(\PageIndex{7}\) האופי המעגלי של פלסמידים והמעגל של כמה גנומים ויראליים בזיהום מאפשרים זאת. שכפול מעגל מתגלגל מתחיל בחיתוך אנזימטי של גדיל אחד של המולקולה המעגלית הדו-גדילית באתר המקור הדו-גדילי (dso). בחיידקים, DNA פולימראז III נקשר לקבוצת 3'-OH של הגדיל המחורץ ומתחיל לשכפל את ה-DNA באופן חד כיווני באמצעות הגדיל הלא מחורץ כתבנית, תוך עקירת הגדיל החתוך תוך כדי כך. השלמת שכפול ה- DNA באתר של הניק המקורי גורמת לעקירה מלאה של הגדיל המחורץ, אשר לאחר מכן עשוי לחזור למולקולת DNA חד-גדילית. לאחר מכן RNA פרימאז מסנתז פריימר ליזום שכפול DNA באתר המוצא החד-גדילי (sso) של מולקולת ה- DNA החד-גדילית (ssDNA), וכתוצאה מכך מולקולת DNA דו-גדילית (dsDNA) זהה למולקולת ה- DNA המעגלית האחרת.
תרגיל \(\PageIndex{4}\)
האם יש גדיל בפיגור בשכפול המעגל המתגלגל? למה או למה לא?
מושגי מפתח וסיכום
- תהליך שכפול ה-DNA הוא שמרני למחצה, מה שמביא לשתי מולקולות DNA, שלכל אחת מהן גדיל DNA הורי אחד וגדיל אחד מסונתז חדש.
- בחיידקים, התחלת השכפול מתרחשת במקור השכפול, כאשר ה-DNA המפותל נפרק על ידי DNA gyrase, מיוצר חד-גדילי על ידי הליקאז, ונקשר על ידי חלבון מחייב חד-גדילי כדי לשמור על מצבו החד-גדילי. Primase מסנתז פריימר RNA קצר, המספק קבוצת 3'-OH חופשית שאליה יכול DNA פולימראז III להוסיף נוקלאוטידים של DNA.
- במהלך התארכות, גדיל ה-DNA המוביל מסונתז ברציפות מפריימר בודד. הגדיל המפגר מסונתז באופן לא רציף בשברי אוקזאקי קצרים, שכל אחד מהם דורש פריימר משלו. הפריימרים של ה-RNA מוסרים ומוחלפים בנוקלאוטידים של DNA על ידי DNA פולימראז I חיידקי, ו-DNA ligase אוטם את הפערים בין השברים הללו.
- הפסקת השכפול בחיידקים כרוכה ברזולוציה של קונקטמרים של DNA מעגליים על ידי טופואיזומראז IV לשחרור שני העותקים של הכרומוזום המעגלי.
- לאיקריוטים יש בדרך כלל כרומוזומים ליניאריים מרובים, שלכל אחד מהם מקורות שכפול מרובים. בסך הכל, שכפול באאוקריוטים דומה לזה שבפרוקריוטים.
- האופי הליניארי של הכרומוזומים האוקריוטיים מחייב טלומרים כדי להגן על גנים בסמוך לקצה הכרומוזומים. טלומראז מרחיב את הטלומרים, ומונע את התדרדרותם, בכמה סוגי תאים.
- שכפול מעגל מתגלגל הוא סוג של סינתזת DNA חד כיוונית מהירה של מולקולת DNA מעגלית המשמשת לשכפול של כמה פלסמידים.