
8.2: קטבוליזם של פחמימות
- Page ID
- 209100

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מטרות למידה
- תאר מדוע הגליקוליזה אינה תלויה בחמצן
- הגדר ותאר את התשואה נטו של מולקולות שלוש פחמן, ATP ו- NADH מגליקוליזה
- הסבר כיצד מולקולות פירובט בעלות שלוש פחמן מומרות לקבוצות אצטיל דו-פחמניות שניתן להעביר למחזור קרבס.
- הגדר ותאר את התשואה נטו של CO 2, GTP/ATP, FADH 2 ו- NADH ממחזור קרבס
- הסבר כיצד ניתן להשתמש במולקולות פחמן ביניים של מחזור קרבס בתא
קיימים מסלולי אנזים נרחבים לפירוק פחמימות ללכידת אנרגיה בקשרי ATP. בנוסף, מסלולים קטבוליים רבים מייצרים מולקולות ביניים המשמשות גם כאבני בניין לאנבוליזם. הבנת תהליכים אלה חשובה מכמה סיבות. ראשית, מכיוון שהתהליכים המטבוליים העיקריים המעורבים משותפים למגוון רחב של אורגניזמים כימוהטרוטרופיים, אנו יכולים ללמוד רבות על חילוף החומרים האנושי על ידי לימוד חילוף החומרים בחיידקים הניתנים למניפולציה קלה יותר כמו אי - קולי. שנית, מכיוון שפתוגנים של בעלי חיים ובני אדם הם גם כימוהטרוטרופים, למידה על פרטי חילוף החומרים בחיידקים אלה, כולל הבדלים אפשריים בין מסלולים חיידקיים ואנושיים, שימושית לאבחון פתוגנים וכן לגילוי טיפולים אנטי-מיקרוביאליים המכוונים לפתוגנים ספציפיים. לבסוף, למידה ספציפית על המסלולים המעורבים בחילוף החומרים הכימוהטרוטרופי משמשת גם בסיס להשוואת אסטרטגיות מטבוליות חריגות אחרות המשמשות חיידקים. למרות שהמקור הכימי של אלקטרונים היוזמים העברת אלקטרונים שונה בין כימוהטרופים לכימואוטוטרופים, תהליכים דומים רבים משמשים בשני סוגי האורגניזמים.
הדוגמה האופיינית המשמשת להכנסת מושגים של חילוף חומרים לתלמידים היא קטבוליזם של פחמימות. עבור כימוהטרוטרופים, הדוגמאות שלנו לחילוף חומרים מתחילות בקטבוליזם של פוליסכרידים כגון גליקוגן, עמילן או תאית. אנזימים כגון עמילאז, המפרק גליקוגן או עמילן, וצלולאזות, המפרקות תאית, עלולים לגרום להידרוליזה של קשרים גליקוזידיים בין מונומרים הגלוקוז בפולימרים אלה, ולשחרר גלוקוז להמשך קטבוליזם.
גליקוליזה
עבור חיידקים, אוקריוטים ורוב הארכאים, הגליקוליזה היא המסלול הנפוץ ביותר לקטבוליזם של גלוקוז; הוא מייצר אנרגיה, נושאי אלקטרונים מופחתים ומולקולות מבשר למטבוליזם תאי. כל אורגניזם חי מבצע צורה כלשהי של גליקוליזה, דבר המצביע על כך שמנגנון זה הוא תהליך מטבולי אוניברסלי עתיק. התהליך עצמו אינו משתמש בחמצן; עם זאת, ניתן לשלב גליקוליזה עם תהליכים מטבוליים נוספים שהם אירוביים או אנאירוביים. הגליקוליזה מתרחשת בציטופלזמה של תאים פרוקריוטים ואיקריוטים. הוא מתחיל במולקולת גלוקוז אחת בעלת שישה פחמנים ומסתיים בשתי מולקולות של סוכר בעל שלושה פחמנים הנקרא פירובט. פירובט עשוי להתפרק עוד יותר לאחר הגליקוליזה כדי לרתום יותר אנרגיה באמצעות נשימה אירובית או אנאירובית, אך אורגניזמים רבים, כולל חיידקים רבים, עשויים שלא להיות מסוגלים לנשום; עבור אורגניזמים אלה, גליקוליזה עשויה להיות המקור היחיד שלהם ליצירת ATP.
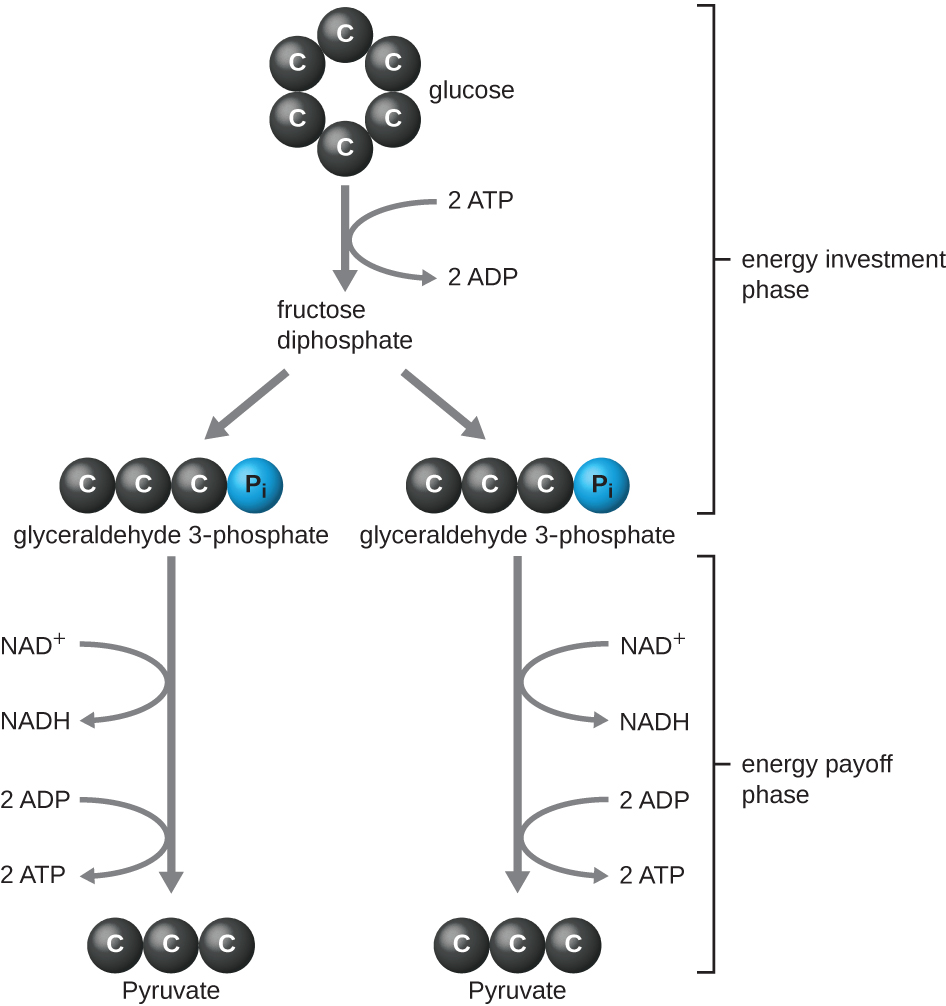
סוג הגליקוליזה המצוי בבעלי חיים וזה הנפוץ ביותר בחיידקים הוא מסלול אמבדן-מאיירהופ-פרנס (EMP), הקרוי על שם גוסטב אמבדן (1874—1933), אוטו מאיירהוף (1884—1951) ויעקב פרנס (1884—1949). גליקוליזה באמצעות מסלול EMP מורכבת משני שלבים נפרדים (איור). \(\PageIndex{1}\) החלק הראשון של המסלול, הנקרא שלב השקעת האנרגיה, משתמש באנרגיה משתי מולקולות ATP כדי לשנות מולקולת גלוקוז כך שניתן יהיה לפצל את מולקולת הסוכר בעלת שישה פחמנים באופן שווה לשתי מולקולות שלוש פחמן זרחניות הנקראות גליצראלדהיד 3-פוספט (G3P). החלק השני של המסלול, הנקרא שלב התמורה האנרגטית, מחלץ אנרגיה על ידי חמצון G3P לפירובט, ייצור ארבע מולקולות ATP והפחתת שתי מולקולות של NAD + לשתי מולקולות של NADH, באמצעות אלקטרונים שמקורם בגלוקוז. (דיון והמחשה של מסלול ה- EMP המלא עם מבנים כימיים ושמות אנזימים מופיעים בנספח ג '.)
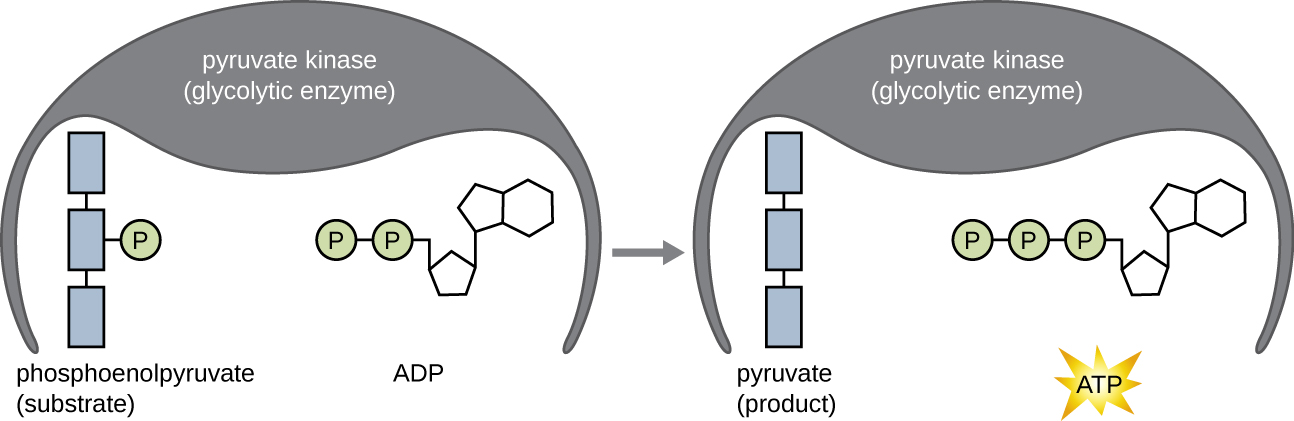
מולקולות ה- ATP המיוצרות בשלב התמורה האנרגטית של הגליקוליזה נוצרות על ידי זרחון ברמת המצע (איור\(\PageIndex{1}\)), אחד משני מנגנונים לייצור ATP. בזרחון ברמת המצע, קבוצת פוספט מוסרת ממולקולה אורגנית ומועברת ישירות למולקולת ADP זמינה, המייצרת ATP. במהלך הגליקוליזה, קבוצות פוספט בעלות אנרגיה גבוהה ממולקולות הביניים מתווספות ל- ADP ליצירת ATP.
בסך הכל, בתהליך זה של גליקוליזה, הרווח הנקי מפירוק מולקולת גלוקוז אחת הוא:
- שתי מולקולות ATP
- שתי מולקולות NADH, ו
- שתי מולקולות פירובט.
מסלולים גליקוליטיים אחרים
כאשר אנו מתייחסים לגליקוליזה, אלא אם כן צוין אחרת, אנו מתייחסים למסלול ה- EMP המשמש בעלי חיים וחיידקים רבים. עם זאת, חלק מהפרוקריוטים משתמשים במסלולים גליקוליטיים חלופיים. חלופה חשובה אחת היא מסלול אנטנר-דודורוף (ED), הקרוי על שם מגליו נתן אנטנר ומייקל דודורוף (1911—1975). למרות שחלק מהחיידקים, כולל הפתוגן גרם שלילי אופורטוניסטי Pseudomonas aeruginosa, מכילים רק את מסלול ה- ED לגליקוליזה, לחיידקים אחרים, כמו אי - קולי, יש את היכולת להשתמש במסלול ED או במסלול ה- EMP.
סוג שלישי של מסלול גליקוליטי המתרחש בכל התאים, השונה בתכלית משני המסלולים הקודמים, הוא מסלול הפנטוז פוספט (PPP) הנקרא גם מסלול הפוספוגלוקונאט או שאנט המונופוספט הקסוז. עדויות מצביעות על כך שה- PPP עשוי להיות המסלול הגליקוליטי האוניברסלי העתיק ביותר. תוצרי הביניים מה- PPP משמשים לביוסינתזה של נוקלאוטידים וחומצות אמינו. לכן, מסלול גליקוליטי זה עשוי להיות מועדף כאשר לתא יש צורך בסינתזת חומצת גרעין ו/או חלבון, בהתאמה. דיון והמחשה של מסלול ה- ED המלא ו- PPP עם מבנים כימיים ושמות אנזימים מופיעים בנספח ג '.
תרגיל \(\PageIndex{1}\)
מתי אורגניזם יכול להשתמש במסלול ED או ב- PPP לגליקוליזה?
תגובת מעבר, קואנזים A ומחזור קרבס
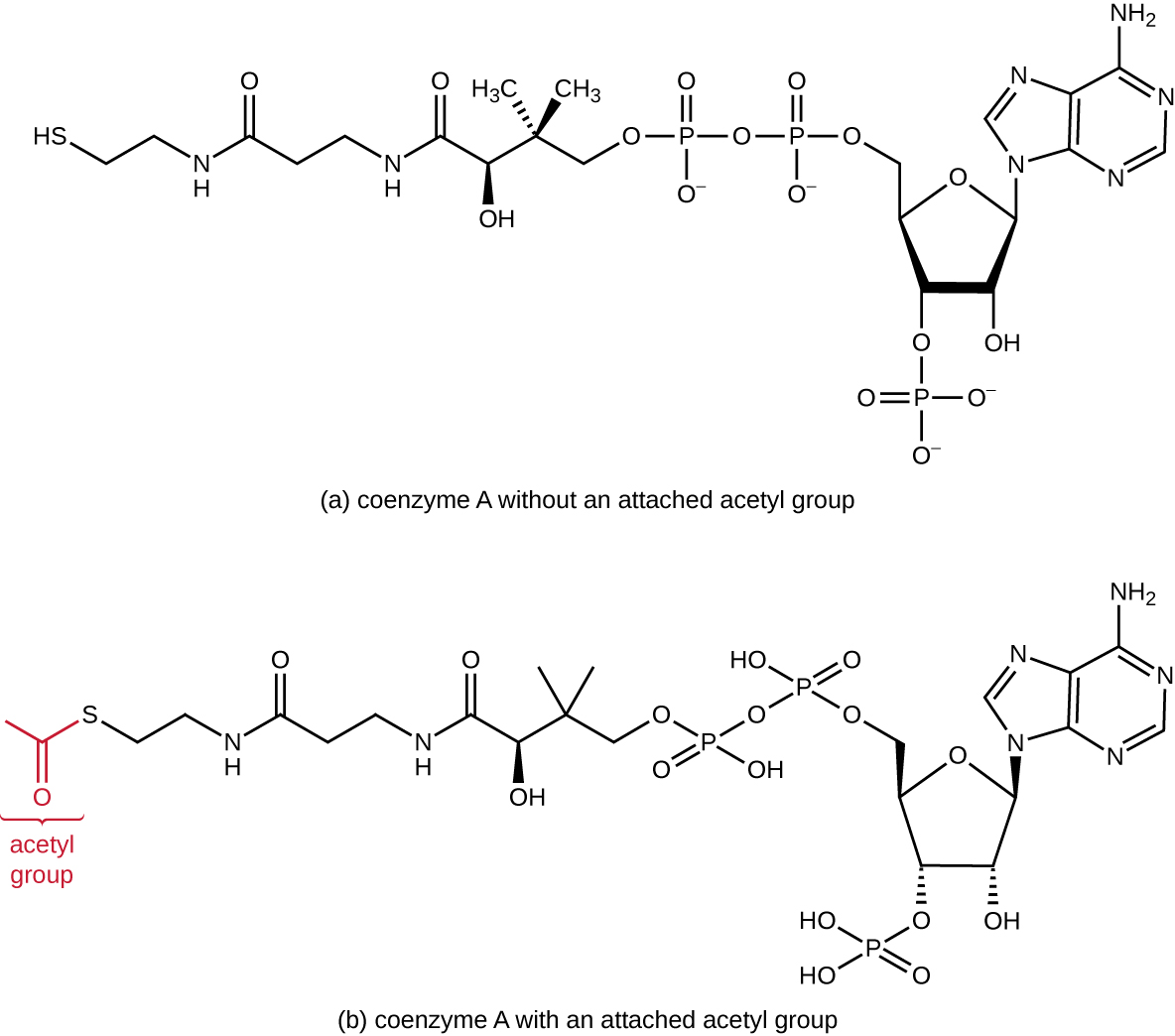
הגליקוליזה מייצרת פירובט, שניתן לחמצן עוד יותר כדי ללכוד יותר אנרגיה. כדי שפירובאט ייכנס למסלול החמצוני הבא, תחילה יש לדה-קרבוקסילציה על ידי קומפלקס האנזים פירובאט דהידרוגנאז לקבוצת אצטיל דו-פחמנית בתגובת המעבר, הנקראת גם תגובת הגשר (ראה נספח ג' ואיור). \(\PageIndex{3}\) בתגובת המעבר, אלקטרונים מועברים גם ל- NAD + ליצירת NADH. כדי להמשיך לשלב הבא של תהליך מטבולי זה, יש לחבר את האצטיל הזעיר יחסית של שני פחמנים לתרכובת נשא גדולה מאוד הנקראת קואנזים A (CoA). תגובת המעבר מתרחשת במטריקס המיטוכונדריאלי של האאוקריוטים; בפרוקריוטים זה מתרחש בציטופלזמה מכיוון שלפרוקריוטים חסרים אברונים סגורים בקרום.
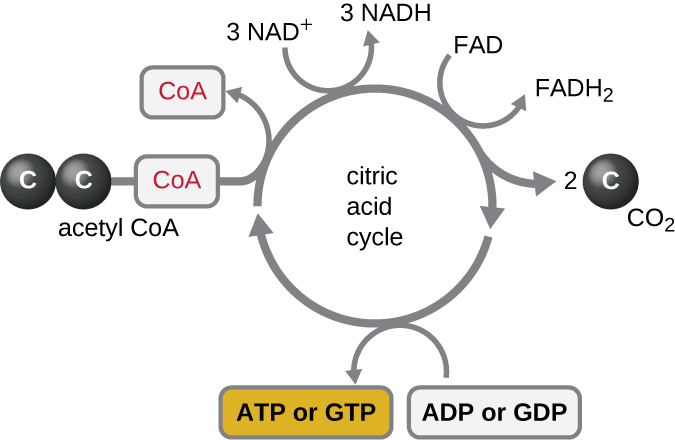
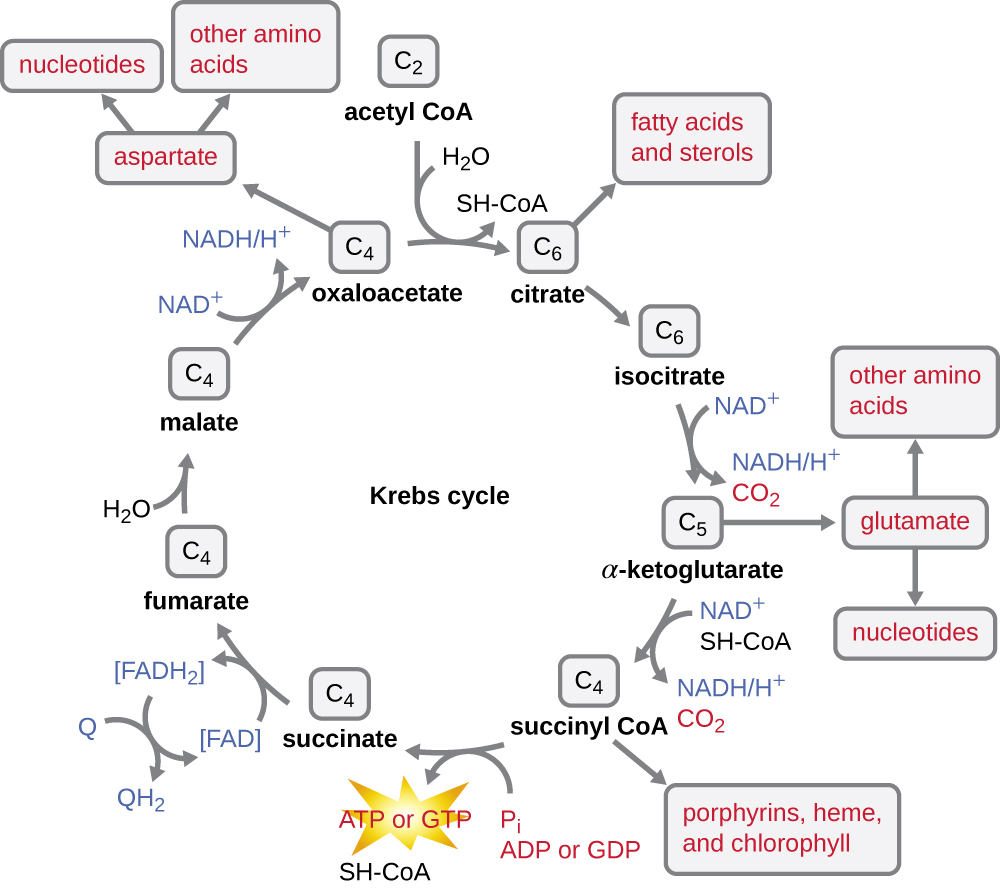
מחזור קרבס מעביר אלקטרונים שנותרו מקבוצת האצטיל המיוצרת במהלך תגובת המעבר למולקולות נושאות אלקטרונים, ובכך מפחית אותן. מחזור קרבס מתרחש גם בציטופלזמה של פרוקריוטים יחד עם הגליקוליזה ותגובת המעבר, אך הוא מתרחש במטריצה המיטוכונדריאלית של תאים אוקריוטיים שם מתרחשת גם תגובת המעבר. מחזור קרבס נקרא על שם מגלהו, המדען הבריטי הנס אדולף קרבס (1900—1981) והוא נקרא גם מחזור חומצת לימון, או מחזור החומצה הטריקרבוקסילית (TCA) מכיוון שלחומצת לימון יש שלוש קבוצות קרבוקסיל במבנה שלה. בניגוד לגליקוליזה, מחזור קרבס הוא לולאה סגורה: החלק האחרון של המסלול מחדש את התרכובת המשמשת בשלב הראשון (איור). \(\PageIndex{4}\) שמונת השלבים של המחזור הם סדרה של תגובות כימיות הלוכדות את קבוצת האצטיל הדו-פחמנית (נשא ה-CoA אינו נכנס למחזור קרבס) מתגובת המעבר, המתווספת לתווך בעל ארבעה פחמנים במחזור קרבס, המייצרת את חומצת הלימון הבינונית בעלת שישה פחמנים (נותנת את השם החלופי למחזור זה). כאשר סיבוב אחד של המחזור חוזר לנקודת ההתחלה של תוצר הביניים בעל ארבעה פחמנים, המחזור מייצר שתי מולקולות CO 2, מולקולת ATP אחת (או מקבילה, כגון גואנוסין טריפוספט [GTP]) המיוצרת על ידי זרחון ברמת המצע, ושלוש מולקולות של NADH ואחת של FADH 2. (דיון והמחשה מפורטת של מחזור קרבס המלא מופיעים בנספח ג ').
למרות שאורגניזמים רבים משתמשים במחזור קרבס כמתואר כחלק ממטבוליזם הגלוקוז, ניתן להשתמש בכמה מתרכובות הביניים במחזור קרבס בסינתזה של מגוון רחב של מולקולות תאיות חשובות, כולל חומצות אמינו, כלורופילים, חומצות שומן ונוקלאוטידים; לכן, המחזור הוא אנבוליים וקטבוליים כאחד (איור). \(\PageIndex{5}\)
מושגי מפתח וסיכום
- גליקוליזה היא השלב הראשון בפירוק הגלוקוז, וכתוצאה מכך נוצר ATP, המיוצר על ידי זרחון ברמת המצע; NADH; ושתי מולקולות פירובט. הגליקוליזה אינה משתמשת בחמצן ואינה תלויה בחמצן.
- לאחר הגליקוליזה, פירובט בעל שלושה פחמנים עובר דה-קרבוקסילציה ליצירת קבוצת אצטיל דו-פחמנית, יחד עם היווצרות NADH. קבוצת האצטיל מחוברת לתרכובת נשא גדולה הנקראת קואנזים A.
- לאחר שלב המעבר, קואנזים A מעביר את האצטיל הדו-פחמני למחזור קרבס, שם שני הפחמנים נכנסים למחזור. בכל סיבוב המחזור, קבוצת אצטיל אחת הנגזרת מגליקוליזה מתחמצנת עוד יותר, מייצרת שלוש מולקולות NADH, אחת FADH 2 ו- ATP אחד על ידי זרחון ברמת המצע, ומשחררת שתי מולקולות CO 2.
- מחזור קרבס עשוי לשמש למטרות אחרות. רבים מתוצרי הביניים משמשים לסינתזה של מולקולות תאיות חשובות, כולל חומצות אמינו, כלורופילים, חומצות שומן ונוקלאוטידים.