
8.3: נשימה סלולרית
- Page ID
- 209093

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מטרות למידה
- השווה והשוואה בין המיקום והתפקוד של מערכת הובלת האלקטרונים בתא פרוקריוטי ובתא אוקריוטי
- השווה והשוואה בין ההבדלים בין רמת המצע לבין זרחון חמצוני
- הסבר את הקשר בין כימיוזמוזה לכוח המניע של הפרוטונים
- תאר את התפקוד והמיקום של סינתאז ATP בתא פרוקריוטי לעומת אוקריוטי
- השווה והשוואה בין נשימה אירובית ואנאירובית
זה עתה דנו בשני מסלולים בקטבוליזם של גלוקוז - גליקוליזה ומחזור קרבס - המייצרים ATP על ידי זרחון ברמת המצע. אולם רוב ה- ATP נוצר במהלך תהליך נפרד הנקרא זרחון חמצוני, המתרחש במהלך הנשימה התאית. הנשימה התאית מתחילה כאשר אלקטרונים מועברים מ- NADH ו- FADH 2 - המיוצרים בגליקוליזה, בתגובת המעבר ומחזור קרבס - באמצעות סדרה של תגובות כימיות למקבל אלקטרונים אנאורגני סופי (חמצן בנשימה אירובית או מולקולות אנאורגניות שאינן חמצן בנשימה אנאירובית). העברות אלקטרונים אלה מתרחשות בחלק הפנימי של קרום התא של תאים פרוקריוטים או במתחמי חלבון מיוחדים בקרום הפנימי של המיטוכונדריה של תאים אוקריוטיים. האנרגיה של האלקטרונים נקצרת ליצירת שיפוע אלקטרוכימי על פני הממברנה, המשמש לייצור ATP על ידי זרחון חמצוני.
מערכת הובלת אלקטרונים
מערכת הובלת האלקטרונים (ETS) היא המרכיב האחרון המעורב בתהליך הנשימה התאית; הוא כולל סדרה של מתחמי חלבון הקשורים לממברנה ונושאי אלקטרונים נלווים נלווים. הובלת אלקטרונים היא סדרה של תגובות כימיות הדומות לחטיבת דלי בכך שאלקטרונים מ- NADH ו- FADH 2 מועברים במהירות ממוביל אלקטרונים ETS אחד למשנהו. נשאים אלה יכולים להעביר אלקטרונים ב- ETS בגלל פוטנציאל החיזור שלהם. כדי שחלבון או כימיקל יקבלו אלקטרונים, עליו להיות בעל פוטנציאל חיזור חיובי יותר מתורם האלקטרונים. לכן, אלקטרונים עוברים מנשאי אלקטרונים בעלי פוטנציאל חיזור שלילי יותר לאלה עם פוטנציאל חיזור חיובי יותר. ארבעת המחלקות העיקריות של נשאי אלקטרונים המעורבים במערכות הובלת אלקטרונים אוקריוטיות ופרוקריוטיות הן הציטוכרומים, הפלבופרוטאינים, חלבוני ברזל-גופרית והקינונים.
בנשימה אירובית, מקבל האלקטרונים הסופי (כלומר, זה בעל פוטנציאל החיזור החיובי ביותר) בסוף ה-ETS הוא מולקולת חמצן (O 2) שהופכת מופחתת למים (H 2 O) על ידי נושא ה-ETS הסופי. נושא אלקטרונים זה, ציטוכרום אוקסידאז, שונה בין סוגי חיידקים וניתן להשתמש בו כדי להבדיל בין חיידקים קרובים לאבחנות. לדוגמה, האופורטוניסט הגראם-שלילי Pseudomonas aeruginosa וה-Vibrio cholerae הגורם לגרם שלילי משתמשים בציטוכרום c אוקסידאז, שניתן לזהות על ידי בדיקת האוקסידאז, בעוד Enterobacteriaceae גרם שליליים, כמו אי - קולי, שליליים לבדיקה זו מכיוון שהם מייצרים סוגים שונים של ציטוכרום אוקסידאז.
ישנן נסיבות רבות בהן נשימה אירובית אינה אפשרית, כולל אחד או יותר מהבאים:
- לתא חסרים גנים המקודדים לציטוכרום אוקסידאז מתאים להעברת אלקטרונים לחמצן בקצה מערכת הובלת האלקטרונים.
- לתא חסרים גנים המקודדים לאנזימים כדי למזער את ההשפעות המזיקות קשות של רדיקלי חמצן מסוכנים המיוצרים במהלך הנשימה האירובית, כגון מי חמצן (H 2 O 2) או סופראוקסיד\((\ce{O2-})\).
- לתא חסר כמות מספקת של חמצן לביצוע נשימה אירובית.
חלופה אפשרית אחת לנשימה אירובית היא נשימה אנאירובית, תוך שימוש במולקולה אנאורגנית שאינה חמצן כמקבל אלקטרונים סופי. ישנם סוגים רבים של נשימה אנאירובית המצויים בחיידקים ובארכיאה. דניטריפירים הם חיידקי קרקע חשובים המשתמשים בחנקה \((\ce{NO3-})\) ובניטריט \((\ce{NO2-})\) כמקבלי אלקטרונים סופיים, המייצרים גז חנקן (N 2). חיידקים רבים נושמים אירובית, כולל E. coli, עוברים לשימוש בחנקה כמקבל אלקטרונים סופי ומייצרים ניטריט כאשר רמות החמצן התרוקנו.
לחיידקים המשתמשים בנשימה אנאירובית יש בדרך כלל מחזור קרבס שלם, כך שאורגניזמים אלה יכולים לגשת לאנרגיה של מולקולות NADH ו- FADH 2 שנוצרו. עם זאת, נושמים אנאירוביים משתמשים בנשאי ETS משתנים המקודדים על ידי הגנום שלהם, כולל קומפלקסים נפרדים להעברת אלקטרונים למקבלי האלקטרונים הסופיים שלהם. שיפועים אלקטרוכימיים קטנים יותר נוצרים ממערכות העברת אלקטרונים אלה, כך שפחות ATP נוצר באמצעות נשימה אנאירובית.
תרגיל \(\PageIndex{1}\)
האם גם נשימה אירובית וגם נשימה אנאירובית משתמשים בשרשרת הובלת אלקטרונים?
כימיוזמוזה, כוח מניע פרוטון וזרחון חמצוני
בכל העברה של אלקטרון דרך ה-ETS, האלקטרון מאבד אנרגיה, אך עם העברות מסוימות, האנרגיה מאוחסנת כאנרגיה פוטנציאלית על ידי שימוש בה לשאיבת יוני מימן (H +) על פני ממברנה. בתאים פרוקריוטים, H+ נשאב אל החלק החיצוני של הממברנה הציטופלזמית (הנקרא החלל הפריפלזמי בחיידקים גרם שליליים וגרם חיוביים), ובתאים אוקריוטיים הם נשאבים מהמטריצה המיטוכונדריאלית על פני הממברנה המיטוכונדריאלית הפנימית אל החלל הבין-ממברני. יש התפלגות לא אחידה של H+ על פני הממברנה שמבססת שיפוע אלקטרוכימי מכיוון שיוני H+ טעונים חיובי (חשמלי) ויש ריכוז גבוה יותר (כימי) בצד אחד של הממברנה. שיפוע אלקטרוכימי זה שנוצר על ידי הצטברות של H + (המכונה גם פרוטון) בצד אחד של הממברנה בהשוואה לצד השני מכונה כוח המניע הפרוטוני (PMF). מכיוון שהיונים המעורבים הם H +, נוצר גם שיפוע pH, כאשר בצד הממברנה הריכוז הגבוה יותר של H+ הוא חומצי יותר. מעבר לשימוש ב- PMF לייצור ATP, כפי שנדון בפרק זה, ניתן להשתמש ב- PMF גם להנעת תהליכים שליליים אחרים מבחינה אנרגטית, כולל הובלת חומרים מזינים וסיבוב דגלים לתנועתיות.
האנרגיה הפוטנציאלית של שיפוע אלקטרוכימי זה שנוצר על ידי ה-ETS גורמת ל-H+ להתפזר על פני ממברנה (קרום הפלזמה בתאים פרוקריוטים והקרום הפנימי במיטוכונדריה בתאים אוקריוטיים). זרימה זו של יוני מימן על פני הממברנה, הנקראת כימיוזמוזה, חייבת להתרחש דרך תעלה בממברנה באמצעות קומפלקס אנזים הקשור לממברנה הנקרא ATP synthase (איור). \(\PageIndex{1}\) הנטייה לתנועה בדרך זו דומה מאוד למים שהצטברו בצד אחד של סכר, הנעים דרך הסכר בעת פתיחתם. סינתאז ATP (כמו שילוב של צריכת וגנרטור של סכר הידרואלקטרי) הוא חלבון מורכב הפועל כמחולל זעיר, המסתובב בכוח ה-H+ המתפזר דרך האנזים, במורד השיפוע האלקטרוכימי שלהם מהמקום שבו יש הרבה הדדיות דוחה H+ למקום שבו יש פחות H +. בתאים פרוקריוטים, H+ זורם מהחלק החיצוני של הממברנה הציטופלזמית לתוך הציטופלזמה, ואילו במיטוכונדריה אוקריוטית, H+ זורם מהחלל הבין-ממברני למטריצה המיטוכונדריאלית. הפיכת חלקי המכונה המולקולרית הזו מחדשת ATP מ- ADP ופוספט אנאורגני (P i) על ידי זרחון חמצוני, מנגנון שני לייצור ATP הקוצר את האנרגיה הפוטנציאלית המאוחסנת בתוך שיפוע אלקטרוכימי.
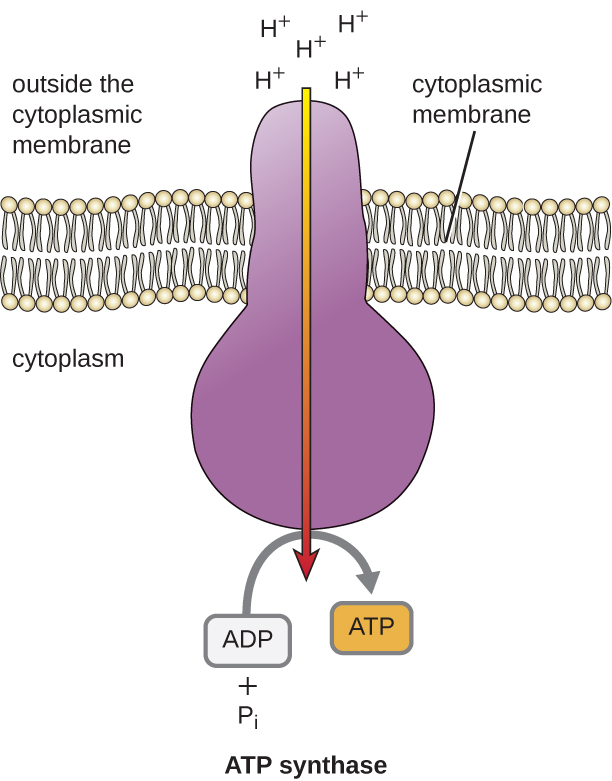
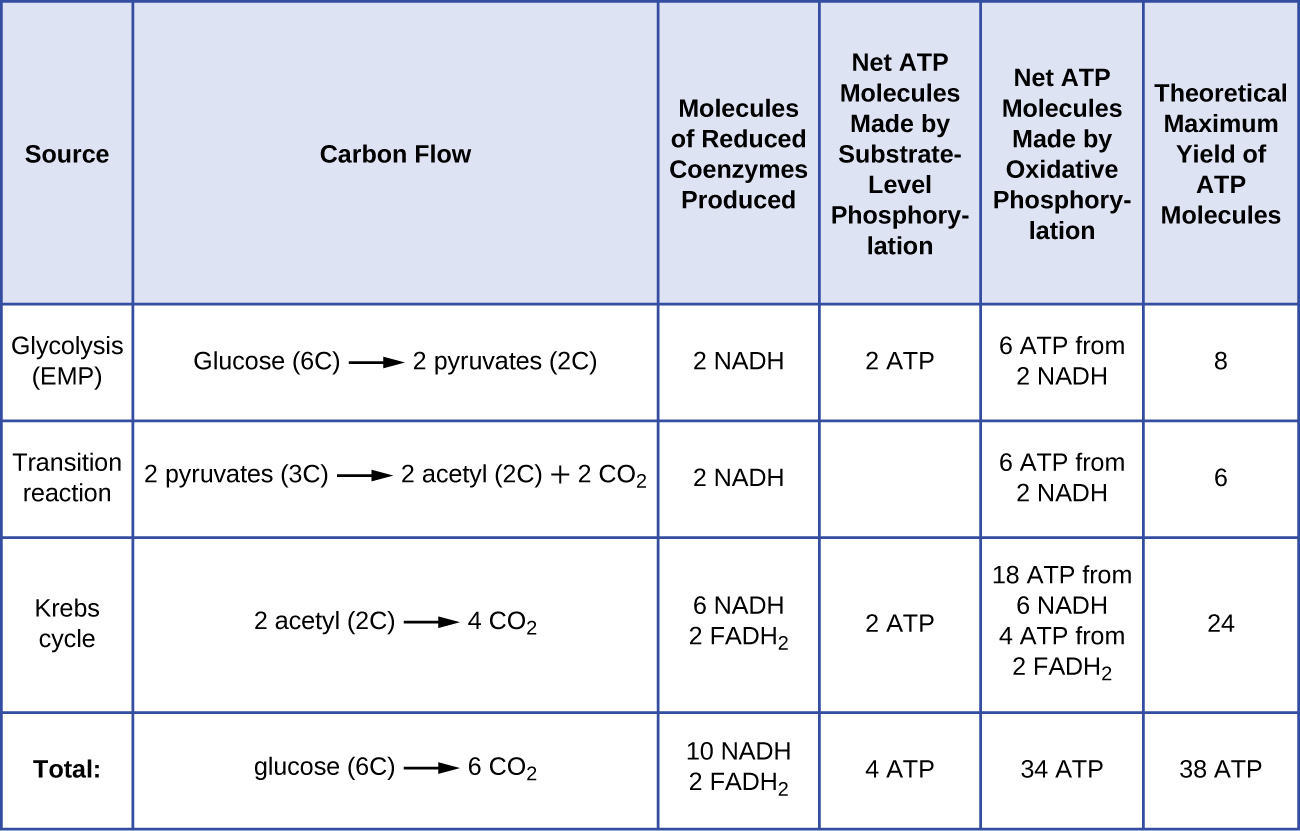
מספר מולקולות ה- ATP הנוצרות מקטבוליזם של גלוקוז משתנה. לדוגמה, מספר יוני המימן שמתחמי מערכת הובלת האלקטרונים יכולים לשאוב דרך הממברנה משתנה בין מינים שונים של אורגניזמים. בנשימה אירובית במיטוכונדריה, מעבר אלקטרונים ממולקולה אחת של NADH מייצר מספיק כוח מניע פרוטון כדי ליצור שלוש מולקולות ATP על ידי זרחון חמצוני, בעוד שמעבר אלקטרונים ממולקולה אחת של FADH 2 מייצר מספיק כוח מניע פרוטון כדי ליצור רק שתי מולקולות ATP. לפיכך, 10 מולקולות NADH המיוצרות לכל גלוקוז במהלך הגליקוליזה, תגובת המעבר ומחזור קרבס נושאות מספיק אנרגיה ליצירת 30 מולקולות ATP, בעוד ששתי מולקולות FADH 2 המיוצרות לכל גלוקוז במהלך תהליכים אלה מספקות מספיק אנרגיה ליצירת ארבע מולקולות ATP. בסך הכל, התשואה המקסימלית התיאורטית של ATP המיוצרת במהלך הנשימה האירובית המלאה של גלוקוז היא 38 מולקולות, כאשר ארבע נוצרות על ידי זרחון ברמת המצע ו -34 נוצרות על ידי זרחון חמצוני (איור). \(\PageIndex{2}\) במציאות, תפוקת ה-ATP הכוללת היא בדרך כלל פחות, ונעה בין אחת ל-34 מולקולות ATP, תלוי אם התא משתמש בנשימה אירובית או בנשימה אנאירובית; בתאים אוקריוטיים, אנרגיה מסוימת מושקעת להובלת תוצרי ביניים מהציטופלזמה לתוך המיטוכונדריה, ומשפיעה על תפוקת ATP.
איור \(\PageIndex{2}\) מסכם את התשואות המרביות התיאורטיות של ATP מתהליכים שונים במהלך הנשימה האירובית המלאה של מולקולת גלוקוז אחת.
תרגיל \(\PageIndex{1}\)
מה הם הפונקציות של כוח המניע פרוטון?
סיכום
- רוב ה- ATP שנוצר במהלך הנשימה התאית של גלוקוז נוצר על ידי זרחון חמצוני.
- מערכת הובלת אלקטרונים (ETS) מורכבת מסדרה של מתחמי חלבון הקשורים לממברנה ונושאי אלקטרונים נלווים ניידים. ה-ETS מוטבע בממברנה הציטופלזמית של פרוקריוטים ובממברנה המיטוכונדריאלית הפנימית של האאוקריוטים.
- לכל קומפלקס ETS יש פוטנציאל חיזור שונה, ואלקטרונים עוברים מנשאי אלקטרונים בעלי פוטנציאל חיזור שלילי יותר לאלה עם פוטנציאל חיזור חיובי יותר.
- כדי לבצע נשימה אירובית, תא דורש חמצן כמקבל האלקטרונים הסופי. תא זקוק גם למחזור קרבס שלם, ציטוכרום אוקסידאז מתאים ואנזימי ניקוי רעלים מחמצן כדי למנוע את ההשפעות המזיקות של רדיקלי חמצן המיוצרים במהלך הנשימה האירובית.
- אורגניזמים המבצעים נשימה אנאירובית משתמשים בנשאי מערכת הובלת אלקטרונים חלופיים להעברה אולטימטיבית של אלקטרונים למקבלי האלקטרונים הסופיים שאינם חמצן.
- חיידקים מראים שונות רבה בהרכב מערכות הובלת האלקטרונים שלהם, שניתן להשתמש בהן למטרות אבחון כדי לסייע בזיהוי פתוגנים מסוימים.
- כאשר אלקטרונים מועברים מ- NADH ו- FADH 2 דרך ETS, האלקטרון מאבד אנרגיה. אנרגיה זו מאוחסנת באמצעות שאיבת H+ על פני הממברנה, ויוצרת כוח מניע פרוטון.
- ניתן לרתום את האנרגיה של כוח המניע הפרוטוני הזה על ידי מתן אפשרות ליוני מימן להתפזר חזרה דרך הממברנה על ידי כימיוזמוזה באמצעות סינתז ATP. כאשר יוני מימן מתפזרים במורד השיפוע האלקטרוכימי שלהם, מרכיבים של סינתאז ATP מסתובבים, מה שהופך ATP מ- ADP ו- P i על ידי זרחון חמצוני.
- נשימה אירובית יוצרת יותר ATP (מקסימום 34 מולקולות ATP) במהלך זרחון חמצוני מאשר נשימה אנאירובית (בין אחת ל-32 מולקולות ATP).