
11.3B: מערכת המשלים
- Page ID
- 209008

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)תאר בקצרה כיצד מופעל מסלול המשלים הקלאסי.תאר בקצרה את ההשפעות המועילות של מוצרי מסלול המשלים הבאים: - C5a
- C3a
- C3b
- C4b
- C3d
- ג5B6789 n (מק)
תאר בקצרה כיצד מסלול הלקטין מופעל. תאר בקצרה כיצד מופעל מסלול המשלים האלטרנטיבי.
- C5a
- C3a
- C3b
- C4b
- C3d
- ג5B6789 n (מק)
בחלק זה נבחן כיצד מערכת המשלים של הגוף מתפקדת להסרת חומרים זיהומיים. מערכת המשלים מתייחסת לסדרה של יותר מ-30 חלבונים מסיסים ומעוצבים מראש המסתובבים בדם ורוחצים את הנוזלים המקיפים את הרקמות. החלבונים מסתובבים בצורה לא פעילה, אך בתגובה להכרה במרכיבים מולקולריים של מיקרואורגניזם, הם מופעלים ברצף, עובדים במפל שבו בקשירה של חלבון אחד מקדם את הקישור של החלבון הבא במפל. ישנם 3 מסלולי השלמה המרכיבים את מערכת המשלים: מסלול המשלים הקלאסי, מסלול הלקטין ומסלול המשלים האלטרנטיבי. המסלולים שונים באופן שבו הם מתחילים ובסופו של דבר מייצרים אנזים מפתח הנקרא C3 convertase:
- מסלול המשלים הקלאסי יוזם על ידי הפעלה של C1. C1 מופעל בעיקר על ידי אינטראקציה עם חלק ה- Fc של מולקולות הנוגדנים IgG או IgM לאחר שהם נקשרו לאנטיגן הספציפי שלהם. C1 מסוגל גם להיקשר ישירות למשטחים של כמה פתוגנים וכן לחלבון C-reactive (CRP) המיוצר במהלך תגובת השלב החריף של חסינות מולדת.
- מסלול הלקטין מופעל על ידי אינטראקציה של פחמימות מיקרוביאליות (לקטינים) עם לקטין מחייב מנוז (MBL) או פיקולינים הנמצאים בפלזמה ובנוזלי הרקמות.
- מסלול המשלים האלטרנטיבי מופעל על ידי קישור C3b למשטחים מיקרוביאליים ולמולקולות נוגדנים.
התוצאות הסופיות ויתרונות ההגנה של כל מסלול, לעומת זאת, זהים. כל מסלולי המשלים מבצעים 6 פונקציות הגנה מולדות מועילות. חלבונים המיוצרים על ידי מסלולי המשלים:
- דלקת טריגר,
- מושכים פגוציטים כימוטקטית לאתר הזיהום,
- לקדם את ההתקשרות של אנטיגנים לפגוציטים (התקשרות משופרת או אופסוניזציה),
- לגרום לתמוגה של חיידקים גראם שליליים, תאים אנושיים המציגים אפיטופים זרים ומעטפות ויראליות,
- לשחק תפקיד בהפעלה של לימפוציטים B נאיביים במהלך חסינות אדפטיבית, ו
- הסר מתחמי חיסון מזיקים מהגוף.
כעת נבחן כל אחד ממסלולי המשלים הללו ונראה כיצד הם מתפקדים כדי להגן על הגוף.
מסלול ההשלמה הקלאסי
מסלול המשלים הקלאסי מופעל בעיקר כאשר קומפלקס חלבון משלים הנקרא C1 מקיים אינטראקציה עם חלק ה- Fc של מולקולות הנוגדנים IgG או IgM לאחר שהם נקשרו לאנטיגן הספציפי שלהם דרך חלק ה- Fab שלהם. C1 מסוגל גם להיקשר ישירות למשטחים של כמה פתוגנים וכן לחלבון C-reactive (CRP) המיוצר במהלך תגובת השלב החריף של חסינות מולדת. קומפלקס C1 מורכב משלושה חלבונים משלימים הנקראים C1q, C1r ו- C1s.
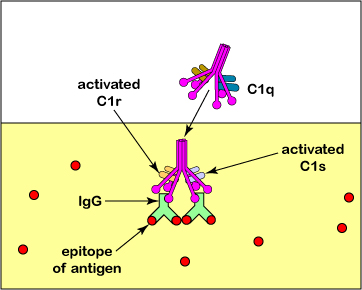
C1 מסוגל גם להיקשר ישירות למשטחים של כמה פתוגנים וכן לחלבון C-reactive (CRP) המיוצר במהלך תגובת השלב החריף של חסינות מולדת.
2. הקישור של C1q מפעיל את החלק C1r של C1 אשר, בתורו, מפעיל C1s. הפעלה זו נותנת C1s פעילות אנזימטית כדי לבקע חלבון משלים C4 לתוך C4a ו- C4b (ראה איור A ואיור B). \(\PageIndex{2}\) \(\PageIndex{2}\)
3. לאחר מכן C2 נקשר ל-C4b והוא מבוקע על ידי C1 לתוך C2a ו-C2b (ראה איור A ואיור B). \(\PageIndex{3}\) \(\PageIndex{3}\)
4. C4b ו- C2a משתלבים ויוצרים C4B2a, ה- C3 המרה. C3 convertase יכול כעת לבקע מאות מולקולות של C3 לתוך C3a ו-C3b (ראה איור). \(\PageIndex{4}\)
6. [1]C5b נקשר לפני השטח של תא המטרה ולאחר מכן קושר את C6, C7, C8 ומספר מונומרים של C9 ליצירת C5B6789 n, מתחם התקפת הממברנה (MAC) (ראה איור ואיור). \(\PageIndex{6}\) \(\PageIndex{7}\)
כפי שצוין לעיל, חלבונים של מסלולי המשלים מבצעים 6 פונקציות הגנה מולדות מועילות. אלה כוללים:
1. דלקת מעוררת: C5a הוא חלבון המשלים החזק ביותר המעורר דלקת. הוא מגיב עם כלי דם הגורמים להתרחבות כלי דם. זה גם גורם לתאי פיטום לשחרר מרחיבי כלי דם כמו היסטמין, הגברת חדירות כלי הדם וכן הגברת הביטוי של מולקולות הדבקה על לויקוציטים ואנדותל כלי הדם, כך שלוקוציטים יכולים להידחק מכלי הדם ולהיכנס לרקמה (דיאפדיזיס). C5a גורם גם לנויטרופילים לשחרר רדיקלי חמצן רעילים להרג חוץ תאי ומעורר חום. במידה פחותה C3a ו- C4a מקדמים גם דלקת. כפי שנראה בהמשך ביחידה זו, דלקת היא תהליך בו כלי הדם מתרחבים והופכים לחדירים יותר, ובכך מאפשרים לתאי הגנה בגוף וכימיקלים להגנה לעזוב את הדם ולהיכנס לרקמות.
2. משיכת פגוציטים כימוטקטית לאתר ההדבקה: C5a מתפקד גם ככימו-מושך לפגוציטים. פגוציטים ינועו לעבר ריכוזים הולכים וגדלים של C5a ולאחר מכן יתחברו, דרך קולטני CR1 שלהם למולקולות C3b המחוברות לאנטיגן. זה יידון בפירוט רב יותר בהמשך יחידה זו תחת phagocytosis.
3. קידום התקשרות אנטיגנים לפגוציטים (התקשרות משופרת או אופסוניזציה): C3b ובמידה פחותה, C4b יכול לתפקד כאופסונינים, כלומר הם יכולים לחבר אנטיגנים לפגוציטים. חלק אחד של C3b נקשר לחלבונים ופוליסכרידים על משטחים מיקרוביאליים; חלק אחר מתחבר לקולטני CR1 על פגוציטים, לימפוציטים B ותאים דנדריטים לפגוציטוזיס משופר. (ראה איור\(\PageIndex{8}\)). למעשה, מולקולת C3b יכולה להיקשר כמעט לכל חלבון או פוליסכריד. תאים אנושיים, לעומת זאת, מייצרים פקטור H הנקשר ל-C3b ומאפשר לפקטור I להשבית את ה-C3b. מצד שני, חומרים כמו LPS על תאי חיידקים מקלים על הקישור של פקטור B ל- C3b וזה מגן על ה- C3b מפני השבתה של גורם I. באופן זה, C3b אינו מתקשר עם התאים שלנו אלא מסוגל לקיים אינטראקציה עם תאים מיקרוביאליים. C3a ו-C5a מגבירים את הביטוי של קולטני C3b על פגוציטים ומגבירים את הפעילות המטבולית שלהם.
4. גרימת תמוגה של חיידקים גראם שליליים, תאים אנושיים המציגים אפיטופים זרים ומעטפות ויראליות: C5B6789 n, מתפקד כמתחם התקפת ממברנה (MAC). [2]זה עוזר להרוס חיידקים גרם שליליים כמו גם תאים אנושיים המציגים אנטיגנים זרים (תאים נגועים בנגיף, תאי גידול וכו') על ידי גרימת תמוגה שלהם; ראה איור ואיור. \(\PageIndex{6}\) \(\PageIndex{7}\) זה יכול גם לפגוע במעטפה של וירוסים עטופים.
5. משמש כאות שני להפעלת לימפוציטים B תמימים במהלך חסינות אדפטיבית: חלק מ- C3b מומר ל- C3d. C3d נקשר לקולטני CR2 על לימפוציטים B. זה משמש אות שני להפעלה של לימפוציטים מסוג B שקולטני תאי B שלהם התקשרו זה עתה עם האנטיגן המקביל להם.
6. הסרת מתחמי חיסון מזיקים מהגוף: C3b ובמידה פחותה, C4b עוזרים בהסרת מתחמי חיסון מזיקים מהגוף. ה- C3b ו- C4b מחברים את מתחמי החיסון לקולטני CR1 על אריתרוציטים. האריתרוציטים מעבירים את המתחמים למקרופאגים קבועים בתוך הטחול והכבד להשמדה. קומפלקסים חיסוניים עלולים להוביל לרגישות יתר מזיקה מסוג III, כפי שיידון בהמשך ביחידה 5 תחת רגישות יתר.
חלק מהכמוסות החיידקיות עשירות בחומצה סיאלית, מרכיב נפוץ בגליקופרוטאין של התא המארח, שיש לו זיקה לחלבון H בסרום, חלבון רגולטורי משלים המוביל לפירוק C3b.
תאר איזו משמעות יש לזה בחיידק המתנגד לפגוציטוזיס ומדוע.
- S. pyogenes מייצר פרוטאז שמבקע את החלבון המשלים C5a.
תאר איזו משמעות יש לזה בחיידק המתנגד לפגוציטוזיס ומדוע.
מסלול לקטין
מסלול הלקטין מופעל על ידי אינטראקציה של פחמימות מיקרוביאליות עם לקטין מחייב מנוז (MBL) או פיקולינים הנמצאים בפלזמה ובנוזלי הרקמות. (לקטינים הם חלבונים המחייבים פחמימות.) מסלול הלקטין מתווך על ידי שתי קבוצות של חלבונים המצויים בפלסמת הדם ובנוזלי רקמות:
1. לקטין מחייב מנוז (MBL) - הידוע גם בשם חלבון מחייב מנוז או MBP. MBL הוא קולטן מסיס לזיהוי דפוסים הנקשר לפחמימות מיקרוביאליות שונות כגון אלו העשירות במנוז או בפוקוז, ול- N-acetylglucosamine (NAG). גליקנים אלה נפוצים בגליקופרוטאינים מיקרוביאליים וגליקוליפידים אך נדירים באלה של בני אדם. MBL מסונתז על ידי הכבד ומשתחרר לזרם הדם כחלק מתגובת השלב החריף שתידון בהמשך יחידה זו. ה- MBL שווה ערך ל- C1q במסלול המשלים הקלאסי.
פיקולינים דומים במבנה שלהם ל- MBL ונקשרים לפחמימות מיקרוביאליות כגון N-acetylglucosamine (NAG), חומצות ליפוטייכואיות וליפופוליסכריד (LPS). פיקולין שווה גם ל- C1q במסלול המשלים הקלאסי.
2. הן לקטין מחייב מנוז (MBL) והן פיקולין יוצרים קומפלקסים עם פרוטאזות סרין הקשורות ל- MBL הנקראות MASP1 ו- MASP2, המקבילות ל- C1r ו - C1s של המסלול הקלאסי.
א. הקישור של ה-MBL (או הפיקולין) לפחמימה המיקרוביאלית מפעיל את ה-MASP2 המשויך ומעניק לו את הפעילות האנזימטית לפצל C4 ל-C4a ו-C4b (ראה איור A ואיור B) . \(\PageIndex{9}\) \(\PageIndex{9}\)
לאחר מכן C2 נקשר ל-C4b ונבקע על ידי MASP2 לתוך C2a ו-C2b (ראה איור A ואיור B ) . \(\PageIndex{10}\) \(\PageIndex{10}\)
ג. C4b ו- C2a משתלבים ויוצרים C4B2a, ה- C3 המרה. C3 convertase יכול כעת לבקע מאות מולקולות של C3 לתוך C3a ו-C3b (ראה איור). \(\PageIndex{11}\)
חלק מהמולקולות של C3b נקשרות ל-C4B2a, ה-C3 קונברטאז, ליצירת C4ב2A3b, קומרטאז C5 שמבקע C5 לתוך C5a ו-C5b (ראה איור). \(\PageIndex{12}\)
ה. [3]C5b נקשר לפני השטח של תא המטרה ולאחר מכן קושר את C6, C7, C8 ומספר מונומרים של C9 ליצירת C5B6789 n, מתחם התקפת הממברנה (MAC) (ראה איור ואיור). \(\PageIndex{6}\) \(\PageIndex{7}\)
התוצאות המועילות של חלבוני המשלים המופעלים זהות למסלול המשלים הקלאסי לעיל. חלבוני המשלים:
1. דלקת טריגר: C5A> C3A> C4a;
2. מושכים פגוציטים כימוטקטית לאתר ההדבקה: C5a;
3. לקדם את ההתקשרות של אנטיגנים לפגוציטים באמצעות התקשרות משופרת או אופסוניזציה: C3B> C4b;
4. לגרום לתמוגה של חיידקים גראם שליליים ותאים אנושיים המציגים אפיטופים זרים: MAC;
5. לשמש אות שני להפעלת לימפוציטים B נאיביים): C3d; ו
6 הסר מתחמי חיסון מזיקים מהגוף: C3B>C4b.
מסלול ההשלמה האלטרנטיבי
מסלול המשלים האלטרנטיבי מתווך על ידי C3b, המיוצר על ידי המסלולים הקלאסיים או הלקטין או מהידרוליזה C3 על ידי מים. (מים יכולים לבצע הידרוליזה של C3 וליצור C3i, מולקולה שמתפקדת באופן דומה ל-C3b.)
הפעלת מסלול המשלים האלטרנטיבי מתחילה כאשר C3b (או C3i) נקשר לדופן התא ולרכיבי שטח אחרים של חיידקים. C3b יכול גם להיקשר לנוגדני IgG. חלבון מסלול חלופי פקטור B משתלב לאחר מכן עם C3b הקשור לתא ליצירת C3bB. לאחר מכן גורם D מפצל את פקטור B הקשור ל- Bb ו- Ba ויוצר C3bbB. חלבון בסרום הנקרא פרופרדין נקשר לאחר מכן ל-Bb ליצירת C3bbBP שמתפקד כ-C3 convertase (ראה איור\(\PageIndex{13}\)) המסוגל לפצל אנזימטית מאות מולקולות של C3 ל-C3a ו-C3b. מסלול המשלים האלטרנטיבי מופעל כעת.
חלק מה-C3b נקשר לאחר מכן לחלק מה-C3bBb ליצירת C3BBb3b, C5 המרה המסוגלת לפצל מולקולות של C5 ל-C5a ו-C5b (ראה איור). \(\PageIndex{14}\) מכאן, מסלול המשלים האלטרנטיבי זהה למסלולי המשלים האחרים.
התוצאות המועילות זהות למסלול המשלים הקלאסי לעיל. חלבוני המשלים:
- דלקת טריגר: C5A> C3A> C4a;
- מושכים פגוציטים כימוטקטית לאתר ההדבקה: C5a;
- לקדם את ההתקשרות של אנטיגנים לפגוציטים באמצעות התקשרות משופרת או אופסוניזציה: C3B> C4b;
- לגרום לתמוגה של חיידקים גראם שליליים, תאים אנושיים המציגים אפיטופים זרים ומעטפות ויראליות: MAC; ו
- לשמש אות שני להפעלת לימפוציטים B נאיביים): C3d;
- הסר מתחמי חיסון מזיקים מהגוף: C3B>C4b.
זכור כי ביחידה 3 למדנו מספר מנגנונים בהם משתמשים חיידקים שונים כדי להתנגד למסלולי המשלים של הגוף. על ידי התנגדות להגנות החיסון המולדות המיידיות הללו, לחיידקים מסוימים יש סיכוי טוב יותר ליישב את המארח שלהם.
סיכום
- החלבונים של מערכת המשלים מסתובבים בצורה לא פעילה, אך בתגובה להכרה במרכיבים מולקולריים של מיקרואורגניזם, הם מופעלים ברצף, עובדים במפל שבו בקשירה של חלבון אחד מקדם את הקישור של החלבון הבא במפל.
- ישנם 3 מסלולי השלמה המרכיבים את מערכת המשלים: מסלול המשלים הקלאסי, מסלול הלקטין ומסלול המשלים האלטרנטיבי.
- מסלול המשלים הקלאסי יוזם על ידי הפעלה של C1. C1 מופעל בעיקר על ידי אינטראקציה עם חלק ה- Fc של מולקולות הנוגדנים IgG או IgM לאחר שהם נקשרו לאנטיגן הספציפי שלהם. C1 מסוגל גם להיקשר ישירות למשטחים של כמה פתוגנים וכן לחלבון C-reactive (CRP) המיוצר במהלך תגובת השלב החריף של חסינות מולדת.
- מסלול הלקטין מופעל על ידי אינטראקציה של פחמימות מיקרוביאליות (לקטינים) עם לקטין מחייב מנוז (MBL) או פיקולינים הנמצאים בפלזמה ובנוזלי הרקמות.
- מסלול המשלים האלטרנטיבי מופעל על ידי קישור C3b למשטחים מיקרוביאליים ולמולקולות נוגדנים.
- כל מסלולי המשלים מבצעים את אותן 6 פונקציות הגנה מולדות מועילות.
- חלבוני המשלים C5a, ובמידה פחותה, C3a ו-C4a מעוררים הרחבת כלי דם ודלקת על מנת להעביר תאי הגנה וכימיקלים הגנה לאתר ההדבקה.
- חלבון המשלים C5a מתפקד גם ככימו-מושך לפגוציטים.
- חלבוני המשלים C3b ובמידה פחותה, C4b יכולים לתפקד כאופסונינים, כלומר הם יכולים לחבר אנטיגנים לפגוציטים.
- חלבוני המשלים C5B6789n, מתפקד כמתחם התקפת ממברנה (MAC) הגורם לתמוגה של חיידקים גראם שליליים, תאים אנושיים המציגים אפיטופים זרים ומעטפות ויראליות.
- חלבון המשלים C3d משמש אות שני להפעלת לימפוציטים B נאיביים במהלך חסינות אדפטיבית.
- חלבוני המשלים C3b ובמידה פחותה, C4b עוזרים בהסרת מתחמי חיסון מזיקים מהגוף.