
15.4: עיבוד RNA באיקריוטים
- Page ID
- 205955

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מיומנויות לפיתוח
- תאר את השלבים השונים בעיבוד RNA
- להבין את המשמעות של אקסונים, אינטרונים וחבור
- הסבר כיצד מעובדים tRNAs ו- rRNA
לאחר שעתוק, pre-mRNAs אוקריוטיים חייבים לעבור מספר שלבי עיבוד לפני שניתן יהיה לתרגם אותם. tRNAs ו-rRNA אוקריוטיים (ופרוקריוטים) עוברים גם עיבוד לפני שהם יכולים לתפקד כרכיבים במנגנון סינתזת החלבון.
עיבוד mRNA
הפרה-mRNA האוקריוטי עובר עיבוד נרחב לפני שהוא מוכן לתרגום. השלבים הנוספים המעורבים בהבשלת mRNA אוקריוטית יוצרים מולקולה עם זמן מחצית חיים ארוך בהרבה מאשר mRNA פרוקריוטי. mRNA אוקריוטי נמשך מספר שעות, ואילו ה- mRNA הטיפוסי E. coli נמשך לא יותר מחמש שניות.
טרום mRNA מצופים לראשונה בחלבונים מייצבי RNA; אלה מגנים על הפרה-mRNA מפני פירוק בזמן שהוא מעובד ומיוצא מהגרעין. שלושת השלבים החשובים ביותר של עיבוד טרום mRNA הם הוספת גורמי ייצוב ואיתות בקצוות 5' ו-3' של המולקולה, והסרת רצפים מתערבים שאינם מציינים את חומצות האמינו המתאימות. במקרים נדירים, ניתן "לערוך" את תמליל ה-mRNA לאחר תעתוקו.
חיבור אבולוציה: עריכת RNA בטריפנוזומים
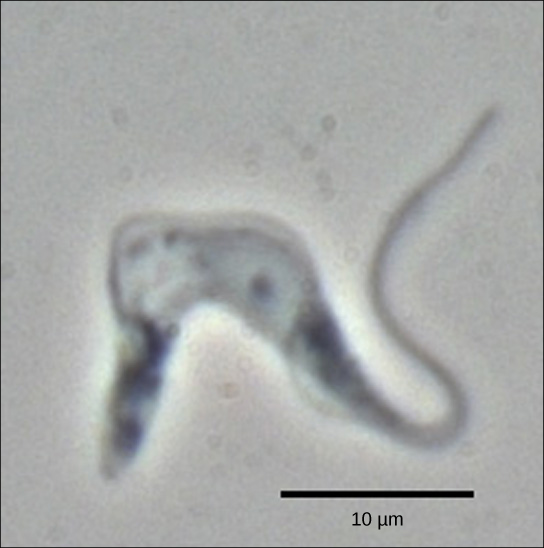
הטריפנוזומים הם קבוצה של פרוטוזואה הכוללת את הפתוגן Trypanosoma brucei, הגורם למחלת שינה בבני אדם (איור). \(\PageIndex{1}\) לטריפנוזומים, ולמעשה לכל שאר האיקריוטים, יש אברונים הנקראים מיטוכונדריה המספקים לתא אנרגיה כימית. מיטוכונדריה הם אברונים המבטאים את ה-DNA שלהם ומאמינים שהם שרידים לקשר סימביוטי בין אוקריוט לפרוקריוט נבלע. ה-DNA המיטוכונדריאלי של טריפנוזומים מציג חריג מעניין מהדוגמה המרכזית: ל-pre-mRNA שלהם אין את המידע הנכון לציין חלבון פונקציונלי. בדרך כלל, הסיבה לכך היא של-mRNA חסרים מספר נוקלאוטידים U. התא מבצע שלב עיבוד RNA נוסף הנקרא עריכת RNA כדי לתקן זאת.
גנים אחרים בגנום המיטוכונדריאלי מקודדים ל- RNA מנחה של 40 עד 80 נוקלאוטידים. אחת או יותר מהמולקולות הללו מקיימות אינטראקציה על ידי זיווג בסיסים משלים עם חלק מהנוקלאוטידים בתמליל הפרה-mRNA. עם זאת, ל-RNA המדריך יש יותר נוקלאוטידים A מאשר ל-pre-mRNA יש נוקלאוטידים U להיקשר איתם. באזורים אלה, ה- RNA המדריך מתפתל החוצה. לקצוות 3' של RNAs מדריכים יש זנב פולי-U ארוך, ובסיסי U אלה מוכנסים באזורים של תמליל הפרה-mRNA שבו לולאות ה-RNA המדריכים. תהליך זה מתווך כולו על ידי מולקולות RNA. כלומר, RNAs מדריכים - ולא חלבונים - משמשים כזרזים בעריכת RNA.
עריכת RNA אינה רק תופעה של טריפנוזומים. במיטוכונדריה של כמה צמחים, כמעט כל pre-mRNA נערכים. עריכת RNA זוהתה גם ביונקים כמו חולדות, ארנבות ואפילו בני אדם. מה יכולה להיות הסיבה האבולוציונית לשלב נוסף זה בעיבוד טרום mRNA? אפשרות אחת היא שלמיטוכונדריה, בהיותן שרידים של פרוקריוטים עתיקים, יש שיטה מבוססת RNA עתיקה לא פחות לוויסות ביטוי גנים. לתמיכה בהשערה זו, עריכות שבוצעו ב- pre-mRNAs שונות בהתאם לתנאי הסלולר. למרות ספקולטיבי, תהליך עריכת ה- RNA עשוי להיות עיכוב מתקופה ראשונית שבה מולקולות RNA, במקום חלבונים, היו אחראיות לזרז תגובות.
מכסה 5 '
בעוד הפרה-mRNA עדיין מסונתז, מכסה 7-methylguanosine מתווסף לקצה 5' של התמליל הגדל על ידי קישור פוספט. חלק זה (קבוצה פונקציונלית) מגן על ה-mRNA המתהווה מפני פירוק. בנוסף, גורמים המעורבים בסינתזת חלבון מזהים את המכסה כדי לסייע ליזום תרגום על ידי ריבוזומים.
זנב פולי-A 3 '
לאחר השלמת ההתארכות, הפרה-mRNA מבוקע על ידי אנדונוקלאז בין רצף קונצנזוס AAUAAA לרצף עשיר ב-GU, ומשאיר את רצף AAUAAA על הפרה-mRNA. אנזים הנקרא פולי-A פולימראז מוסיף לאחר מכן מחרוזת של כ-200 שאריות A, הנקראות זנב פולי-A. שינוי זה מגן עוד יותר על הפרה-mRNA מפני פירוק ומאותת על ייצוא הגורמים התאיים שהתמליל צריך לציטופלזמה.
שחבור טרום mRNA
גנים אוקריוטיים מורכבים מאקסונים, התואמים לרצפים מקודדי חלבון (למשל מסמל שהם לחוצים לשעבר), ורצפי int ervening הנקראים אינטרונים (int- ron מציין את תפקידם המחולל), שעשויים להיות מעורבים בוויסות גנים אך מוסרים מהפרה-mRNA במהלך העיבוד. רצפי אינטרון ב-mRNA אינם מקודדים לחלבונים פונקציונליים.
גילוי האינטרונים הפתיע חוקרים בשנות ה-70 שציפו ש-pre-mRNA יציינו רצפי חלבון ללא עיבוד נוסף, כפי שצפו בפרוקריוטים. הגנים של אוקריוטים גבוהים מכילים לעתים קרובות אינטרון אחד או יותר. אזורים אלה עשויים להתאים לרצפים רגולטוריים; עם זאת, המשמעות הביולוגית של אינטרונים רבים או שיש אינטרונים ארוכים מאוד בגן אינה ברורה. יתכן שאינטרונים מאטים את ביטוי הגנים מכיוון שלוקח יותר זמן לתמלל pre-mRNA עם הרבה אינטרונים. לחלופין, אינטרונים עשויים להיות שרידי רצף לא פונקציונליים שנותרו מהתמזגות של גנים עתיקים לאורך האבולוציה. זה נתמך על ידי העובדה שאקסונים נפרדים מקודדים לעתים קרובות תת-יחידות חלבון או תחומים נפרדים. לרוב, ניתן לבצע מוטציה של רצפי האינטרונים מבלי להשפיע בסופו של דבר על מוצר החלבון.
יש להסיר את כל האינטרונים של פרה-mRNA לחלוטין ומדויק לפני סינתזת החלבון. אם התהליך טועה אפילו על ידי נוקלאוטיד בודד, מסגרת הקריאה של האקסונים המחוברים מחדש תשתנה, והחלבון המתקבל יהיה לא מתפקד. תהליך הסרת האינטרונים וחיבור האקסונים מחדש נקרא שחבור (איור). \(\PageIndex{2}\) אינטרונים מוסרים ומתפרקים בעוד הפרה-mRNA עדיין בגרעין. שחבור מתרחש על ידי מנגנון ספציפי לרצף המבטיח שהאינטרונים יוסרו ואקסונים יצטרפו מחדש עם הדיוק והדיוק של נוקלאוטיד בודד. השחבור של טרום mRNA מתבצע על ידי קומפלקסים של חלבונים ומולקולות RNA הנקראות spliceosomes.
חיבור אמנות
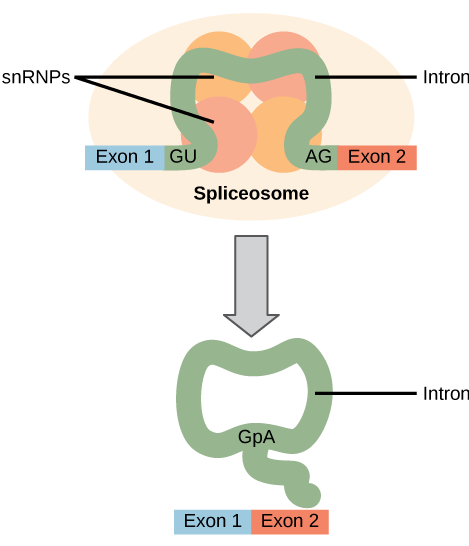
טעויות בחבור מעורבות בסרטן ובמחלות אנושיות אחרות. אילו סוגים של מוטציות עלולים להוביל לשגיאות שחבור? חשבו על תוצאות אפשריות שונות אם מתרחשות שגיאות שחבור.
שים לב שיותר מ-70 אינטרונים בודדים יכולים להיות נוכחים, וכל אחד מהם צריך לעבור את תהליך השחבור - בנוסף למכסת 5' ותוספת של זנב פולי-A - רק כדי ליצור מולקולת mRNA אחת הניתנת לתרגום.
קישור ללמידה
ראה כיצד אינטרונים מוסרים במהלך שחבור RNA באתר זה.
עיבוד של tRNAs ו- rRNA
ה- tRNAs ו- rRNAs הם מולקולות מבניות שיש להן תפקידים בסינתזת חלבון; עם זאת, RNA אלה אינם מתורגמים בעצמם. טרום rRNA מתועתקים, מעובדים ומורכבים לריבוזומים בגרעין. טרום tRNA מתועתקים ומעובדים בגרעין ולאחר מכן משתחררים לציטופלזמה שם הם מקושרים לחומצות אמינו חופשיות לסינתזת חלבון.
רוב ה-tRNA וה-rRNAs באאוקריוטים ובפרוקריוטים מתועתקים לראשונה כמולקולת מבשר ארוכה המשתרעת על פני מספר rRNAs או tRNAs. לאחר מכן אנזימים מבקעים את המבשרים ליחידות משנה המתאימות לכל RNA מבני. חלק מהבסיסים של pre-rRNAs מתילטים; כלומר, קבוצה —CH 3 (קבוצה פונקציונלית מתיל) מתווספת ליציבות. מולקולות טרום tRNA עוברות גם מתילציה. בדומה ל- pre-mRNAs, כריתה של יחידת משנה מתרחשת ב- pre-RNAs אוקריוטיים המיועדים להפוך ל- tRNAs או rRNAs.
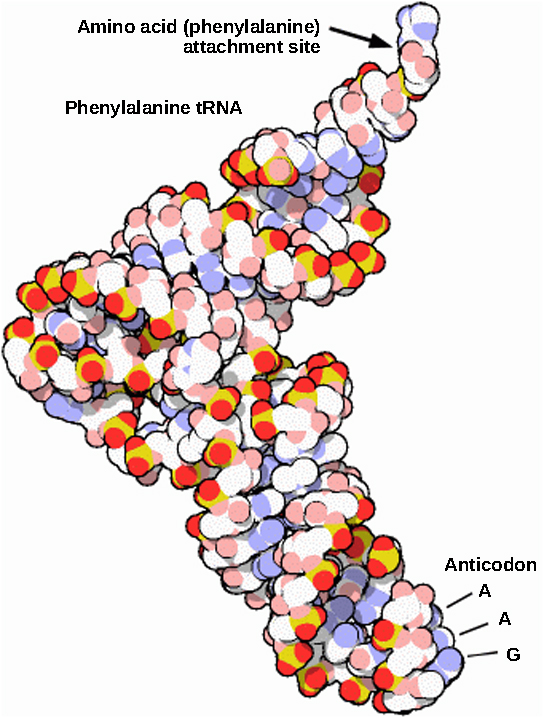
RRNAs בוגרים מהווים כ-50 אחוז מכל ריבוזום. חלק ממולקולות ה-RNA של הריבוזום הן מבניות בלבד, בעוד שלאחרות יש פעילויות קטליטיות או מחייבות. tRNAs בוגרים מקבלים מבנה תלת מימדי באמצעות קישור מימן תוך מולקולרי כדי למקם את אתר הקישור של חומצות אמינו בקצה אחד ואת האנטיקודון בקצה השני (איור). \(\PageIndex{3}\) האנטיקודון הוא רצף של שלושה נוקלאוטידים ב-tRNA המקיים אינטראקציה עם קודון mRNA באמצעות זיווג בסיסים משלים.
סיכום
פרה-mRNA האוקריוטיים משתנים עם מכסה מתילגואנוסין 5' וזנב פולי-A. מבנים אלה מגנים על ה- mRNA הבוגר מפני השפלה ועוזרים לייצא אותו מהגרעין. טרום mRNA עוברים גם שחבור, שבו אינטרונים מוסרים ואקסונים מחוברים מחדש בדיוק של נוקלאוטיד יחיד. רק mRNA מוגמר שעברו מכסה של 5 ', פוליאדנילציה 3' ושחבור אינטרונים מיוצאים מהגרעין לציטופלזמה. ניתן לעבד פרה-RRNAs ו-pre-tRNAs על ידי מחשוף תוך מולקולרי, שחבור, מתילציה והמרה כימית של נוקלאוטידים. לעיתים רחוקות, עריכת RNA מבוצעת גם כדי להכניס בסיסים חסרים לאחר סינתזה של mRNA.
חיבורי אמנות
איור\(\PageIndex{2}\): טעויות בשחבור מעורבות בסרטן ובמחלות אנושיות אחרות. אילו סוגים של מוטציות עלולים להוביל לשגיאות שחבור? חשבו על תוצאות אפשריות שונות אם מתרחשות שגיאות שחבור.
- תשובה
-
מוטציות ברצף זיהוי הספליזוזום בכל קצה של האינטרון, או בחלבונים וב- RNA המרכיבים את הספליזוזום, עלולות לפגוע בשחבור. מוטציות עשויות להוסיף גם אתרי זיהוי ספליזוזומים חדשים. שגיאות שחבור עלולות להוביל לשמירה על אינטרונים ב-RNA שחבור, כריתת אקסונים או שינויים במיקום אתר השחבור.
רשימת מילים
- כובע 7-מתילגואנוסין
- שינוי נוסף לקצה 5 'של pre-mRNA כדי להגן על mRNA מפני השפלה ולסייע בתרגום
- אנטיקודון
- רצף של שלושה נוקלאוטידים במולקולת tRNA המתאימה לקודון mRNA
- אקסון
- רצף קיים ב-mRNA המקודד לחלבון לאחר השלמת שחבור טרום mRNA
- אינטרון
- רצפים מתערבים שאינם מקודדים לחלבון שחוברים מ-mRNA במהלך העיבוד
- זנב פולי-א
- שינוי נוסף לקצה 3 'של pre-mRNA כדי להגן על mRNA מפני פירוק ולסייע לייצוא mRNA מהגרעין
- עריכת RNA
- שינוי ישיר של נוקלאוטידים אחד או יותר ב-mRNA שכבר סונתז
- שחבור
- תהליך של הסרת אינטרונים וחיבור מחדש של אקסונים ב-pre-mRNA