
15.3: תמלול אוקריוטי
- Page ID
- 205946

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מיומנויות לפיתוח
- רשום את השלבים בתעתיק אוקריוטי
- דון בתפקידם של פולימראזות RNA בתעתיק
- השווה והשוואה בין שלושת פולימראזות ה-RNA
- הסבר את המשמעות של גורמי שעתוק
פרוקריוטים ואיקריוטים מבצעים ביסודו את אותו תהליך של שעתוק, עם כמה הבדלים מרכזיים. ההבדל החשוב ביותר בין פרוקריוטים לאיקריוטים הוא הגרעין והאברונים הקשורים לממברנה של האחרון. כאשר הגנים קשורים בגרעין, התא האוקריוטי חייב להיות מסוגל להעביר את ה- mRNA שלו לציטופלזמה ועליו להגן על ה- mRNA שלו מפני פירוק לפני שהוא מתורגם. אוקריוטים משתמשים גם בשלושה פולימראזות שונות שכל אחת מהן מתעתקת תת-קבוצה אחרת של גנים. mRNA אוקריוטי הם בדרך כלל מונוגניים, כלומר הם מציינים חלבון בודד.
התחלת תמלול באיקריוטים
בניגוד לפולימראז הפרוקריוטי שיכול להיקשר לתבנית DNA בפני עצמו, אוקריוטים דורשים מספר חלבונים אחרים, הנקראים גורמי שעתוק, כדי להיקשר תחילה לאזור האמרגן ולאחר מכן לסייע בגיוס הפולימראז המתאים.
שלושת פולימראזות ה- RNA האוקריוטיות
התכונות של סינתזת mRNA אוקריוטית מורכבות במידה ניכרת מאלו של פרוקריוטים. במקום פולימראז בודד הכולל חמש יחידות משנה, לאאוקריוטים יש שלוש פולימראזות שכל אחת מהן מורכבת מ-10 יחידות משנה או יותר. כל פולימראז אוקריוטי דורש גם קבוצה מובהקת של גורמי שעתוק כדי להביא אותו לתבנית ה-DNA.
RNA פולימראז I ממוקם בגרעין, תת-מבנה גרעיני מיוחד שבו RNA ריבוזומלי (rRNA) מתועתק, מעובד ומורכב לריבוזומים (טבלה). \(\PageIndex{1}\) מולקולות ה-rRNA נחשבות ל-RNA מבניים מכיוון שיש להן תפקיד תאי אך אינן מתורגמות לחלבון. ה- rRNAs הם מרכיבים של הריבוזום והם חיוניים לתהליך התרגום. RNA פולימראז I מסנתז את כל ה- rRNA למעט מולקולת ה- rRNA 5S. הכינוי "S" חל על יחידות "Svedberg", ערך לא-תוסף המאפיין את המהירות שבה משקע חלקיק במהלך צנטריפוגה.
טבלה\(\PageIndex{1}\): מיקומים, מוצרים ורגישויות של שלושת פולימראזות ה- RNA האוקריוטיות
| RNA פולימראז | תא סלולרי | תוצר של תמלול | α-amanitin רגישות |
|---|---|---|---|
| I | נוקלאולוס | כל ה-rRNA למעט 5S rRNA | חסר רגישות |
| II | גרעין | כל הפרה-mRNA הגרעיני המקודד לחלבון | רגיש במיוחד |
| III | גרעין | 5S rRNA, tRNAs ו-RNA גרעיניים קטנים | רגיש בינוני |
RNA פולימראז II ממוקם בגרעין ומסנתז את כל הפרה-mRNA הגרעיני המקודד לחלבון. טרום mRNA אוקריוטיים עוברים עיבוד נרחב לאחר שעתוק אך לפני התרגום. לשם הבהרה, הדיון של מודול זה על שעתוק ותרגום באיקריוטים ישתמש במונח "mRNAs" כדי לתאר רק את המולקולות הבוגרות והמעובדות שמוכנות לתרגום. RNA פולימראז II אחראי לתמלול הרוב המכריע של הגנים האוקריוטיים.
RNA פולימראז III ממוקם גם בגרעין. פולימראז זה מתמלל מגוון של RNAs מבניים הכוללים את 5S pre-rRNA, העברת pre-RNAs (pre-tRNAs), ו pre-RNAs גרעיניים קטנים. ל- tRNAs יש תפקיד קריטי בתרגום; הם משמשים כמולקולות המתאם בין תבנית ה- mRNA לשרשרת הפוליפפטיד ההולכת וגדלה. ל-RNAs גרעיניים קטנים יש מגוון פונקציות, כולל "שחבור" pre-mRNAs וויסות גורמי שעתוק.
מדען המאפיין גן חדש יכול לקבוע איזה פולימראז מתמלל אותו על ידי בדיקה האם הגן מתבטא בנוכחות רעל פטריות מסוים, α-amanitin (טבלה למעלה). מעניין לציין כי α-amanitin המיוצר על ידי אמניטה פאלואידים, פטריית כובע המוות, משפיע על שלושת הפולימראזות בצורה שונה מאוד. RNA פולימראז I אינו רגיש לחלוטין ל-α-amanitin, כלומר הפולימראז יכול לתמלל DNA בַּמַבחֵנָה בנוכחות רעל זה. לעומת זאת, RNA פולימראז II רגיש ביותר ל-α-amanitin, ו-RNA פולימראז III רגיש במידה בינונית. הכרת הפולימראז המתעתק יכולה לרמז על חוקר לתפקוד הכללי של הגן הנחקר. מכיוון ש-RNA פולימראז II מתמלל את הרוב המכריע של הגנים, נתמקד בפולימראז זה בדיונים הבאים שלנו על גורמי שעתוק אוקריוטיים ומקדמים.
מבנה של מקדם RNA פולימראז II
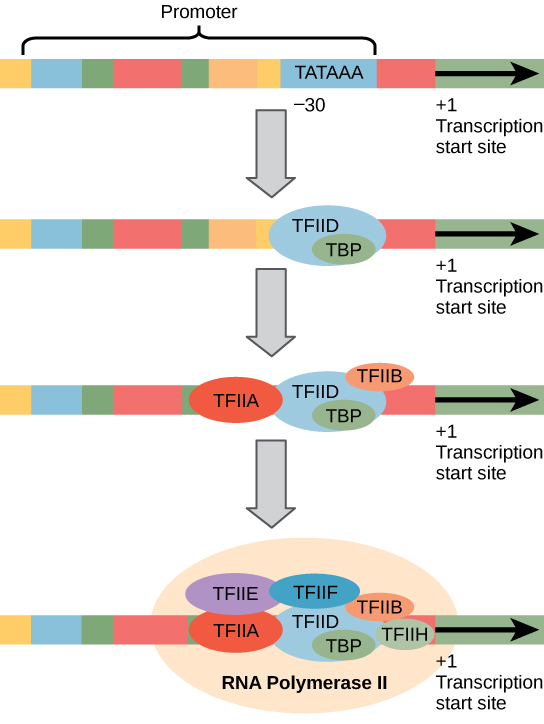
מקדמים אוקריוטיים גדולים ומורכבים בהרבה ממקדמים פרוקריוטים, אך לשניהם יש תיבת TATA. לדוגמה, בגן התימידין קינאז של העכבר, תיבת TATA ממוקמת בערך -30 ביחס לאתר ההתחלה (+1) (איור). \(\PageIndex{1}\) עבור גן זה, רצף תיבות TATA המדויק הוא TATAAAA, כפי שנקרא בכיוון 5' עד 3' בגדיל הלא תבנית. רצף זה אינו זהה לתיבת E. coli TATA, אך הוא משמר את האלמנט העשיר A — T. היציבות התרמית של קשרי A-T נמוכה וזה עוזר לתבנית ה- DNA להירגע מקומית לקראת שעתוק.
חיבור אמנות

מדען מחבר מקדם אוקריוטי מול גן חיידקי ומכניס את הגן לכרומוזום חיידקי. האם היית מצפה שהחיידקים יתמללו את הגן?
הגנום של העכבר כולל גן אחד ושני פסאודוגנים לתימידין קינאז ציטופלזמי. פסאודוגנים הם גנים שאיבדו את יכולת קידוד החלבון שלהם או שאינם באים לידי ביטוי עוד על ידי התא. פסאודוגנים אלה מועתקים מ- mRNA ומשולבים בכרומוזום. לדוגמה, למקדם התימידין קינאז של העכבר יש גם תיבת CAAT שמורה (GGCCAATCT) בערך -80. רצף זה חיוני ומעורב בגורמי שעתוק מחייבים. בהמשך הזרם של תיבת TATA, מקדמים אוקריוטיים עשויים להכיל גם קופסה אחת או יותר עשירה ב- GC (GGCG) או תיבות אוקטמר (ATTTGCAT). אלמנטים אלה קושרים גורמים תאיים המגבירים את היעילות של התחלת שעתוק ולעתים קרובות מזוהים בגנים "פעילים" יותר המתבטאים כל הזמן על ידי התא.
גורמי שעתוק עבור RNA פולימראז II
המורכבות של שעתוק אוקריוטי אינה מסתיימת בפולימראזות ובמקדמים. צבא של גורמי שעתוק בסיסיים, משפרים ומשתיקי קול מסייע גם לווסת את התדירות שבה מסונתז פרה-mRNA מגן. משפרים ומשתיקים משפיעים על יעילות התעתיק אך אינם נחוצים להמשך התמלול. גורמי שעתוק בסיסיים הם מכריעים ביצירת קומפלקס התחלה מוקדמת על תבנית ה-DNA המגייס לאחר מכן RNA פולימראז II לתחילת שעתוק.
שמות גורמי השעתוק הבסיסיים מתחילים ב- "TFII" (זהו גורם השעתוק של RNA פולימראז II) ומצוינים באותיות A—J גורמי השעתוק נופלים במקומם באופן שיטתי על תבנית ה- DNA, כאשר כל אחד מהם מייצב עוד יותר את קומפלקס ההתחלה ותורם לגיוס RNA פולימראז II.
התהליכים של הבאת פולימראזות RNA I ו-III לתבנית ה-DNA כוללים אוספים מעט פחות מורכבים של גורמי שעתוק, אך הנושא הכללי זהה. שעתוק אוקריוטי הוא תהליך מוסדר היטב הדורש מגוון חלבונים כדי לקיים אינטראקציה זה עם זה ועם גדיל ה-DNA. למרות שתהליך השעתוק באאוקריוטים כרוך בהשקעה מטבולית גדולה יותר מאשר בפרוקריוטים, הוא מבטיח שהתא מתמלל בדיוק את הפרה-mRNA שהוא זקוק לסינתזת חלבון.
חיבור אבולוציה: האבולוציה של מקדמים
התפתחות הגנים עשויה להיות מושג מוכר. מוטציות יכולות להתרחש בגנים במהלך שכפול ה-DNA, והתוצאה עשויה להועיל לתא או לא. על ידי שינוי אנזים, חלבון מבני או גורם אחר, תהליך המוטציה יכול לשנות פונקציות או תכונות פיזיות. עם זאת, מקדמים אוקריוטיים ורצפים רגולטוריים אחרים של גנים עשויים להתפתח גם כן. לדוגמה, שקול גן שבמשך דורות רבים הופך להיות בעל ערך רב יותר לתא. אולי הגן מקודד לחלבון מבני שהתא צריך לסנתז בשפע לתפקוד מסוים. אם זה המקרה, יהיה מועיל לתא שהמקדם של הגן הזה יגייס גורמי שעתוק ביעילות רבה יותר ויגביר את ביטוי הגנים.
מדענים שבדקו את האבולוציה של רצפי פרומטור דיווחו על תוצאות שונות. בין השאר, הסיבה לכך היא שקשה להסיק בדיוק היכן מתחיל ונגמר מקדם אוקריוטי. חלק מהמקדמים מתרחשים בתוך גנים; אחרים ממוקמים רחוק מאוד במעלה הזרם, או אפילו במורד הזרם, של הגנים שהם מווסתים. עם זאת, כאשר חוקרים הגבילו את בדיקתם לרצפי מקדם ליבה אנושיים שהוגדרו בניסוי כרצפים הקושרים את קומפלקס ההתחלה המוקדמת, הם גילו שמקדמים מתפתחים אפילו מהר יותר מגנים מקודדי חלבון.
עדיין לא ברור כיצד התפתחות האמרגן עשויה להתאים לאבולוציה של בני אדם או אורגניזמים גבוהים אחרים. עם זאת, האבולוציה של מקדם לייצור יעיל פחות או יותר מתוצר גנטי נתון היא חלופה מסקרנת לאבולוציה של הגנים עצמם. 1
מבני מקדם לפולימראזות RNA I ו-III
באאוקריוטים, יסודות האמרגן השמורים שונים עבור גנים המתועתקים על ידי פולימראזות RNA I, II ו- III. RNA פולימראז I מתמלל גנים שיש להם שני רצפי פרומטור עשירים ב-GC באזור -45 עד +20. רצפים אלה לבדם מספיקים להתחלת שעתוק להתרחש, אך מקדמים עם רצפים נוספים באזור מ -180 עד -105 במעלה הזרם של אתר ההתחלה ישפרו עוד יותר את ההתחלה. לגנים המתועתקים על ידי RNA פולימראז III יש מקדמים או מקדמים במעלה הזרם המתרחשים בתוך הגנים עצמם.
התארכות וסיום אוקריוטיים
לאחר היווצרות קומפלקס ההתחלה, הפולימראז משתחרר מגורמי השעתוק האחרים, והתארכות מותרת להתקדם כפי שהיא מתרחשת בפרוקריוטים כאשר הפולימראז מסנתז פרה-mRNA בכיוון 5' עד 3'. כפי שנדון קודם לכן, RNA פולימראז II מתמלל את החלק העיקרי של גנים אוקריוטיים, כך שחלק זה יתמקד באופן שבו פולימראז זה משיג התארכות וסיום.
למרות שתהליך ההתארכות האנזימטי זהה במהותו באאוקריוטים ובפרוקריוטים, תבנית ה- DNA מורכבת יותר. כאשר תאים אוקריוטיים אינם מתחלקים, הגנים שלהם קיימים כמסה מפוזרת של DNA וחלבונים הנקראים כרומטין. ה-DNA ארוז היטב סביב חלבוני היסטון טעונים במרווחים חוזרים ונשנים. מתחמי DNA -היסטון אלה, הנקראים ביחד נוקלאוזומים, מרווחים באופן קבוע וכוללים 146 נוקלאוטידים של DNA הפצועים סביב שמונה היסטונים כמו חוט סביב סליל.
כדי שתתרחש סינתזת פולינוקלאוטידים, מכונות השעתוק צריכות להזיז היסטונים מהדרך בכל פעם שהיא נתקלת בנוקלאוזום. זה מושג על ידי קומפלקס חלבונים מיוחד בשם FACT, המייצג "מקל על שעתוק הכרומטין". קומפלקס זה מושך היסטונים מתבנית ה-DNA כאשר הפולימראז נע לאורכו. ברגע שהפרה-mRNA מסונתז, קומפלקס FACT מחליף את ההיסטונים כדי ליצור מחדש את הנוקלאוזומים.
הפסקת השעתוק שונה עבור הפולימראזות השונות. שלא כמו בפרוקריוטים, התארכות על ידי RNA פולימראז II באאוקריוטים מתרחשת 1,000-2,000 נוקלאוטידים מעבר לקצה הגן המתועתק. זנב קדם-mRNA זה מוסר לאחר מכן על ידי מחשוף במהלך עיבוד mRNA. מצד שני, פולימראזות RNA I ו-III דורשות אותות סיום. גנים המתועתקים על ידי RNA פולימראז I מכילים רצף ספציפי של 18 נוקלאוטידים המוכר על ידי חלבון סיום. תהליך הסיום ב-RNA פולימראז III כולל סיכת ראש mRNA הדומה לסיום בלתי תלוי ב-rho של שעתוק בפרוקריוטים.
סיכום
שעתוק באאוקריוטים כולל אחד משלושה סוגים של פולימראזות, בהתאם לגן המתועתק. RNA פולימראז II מתמלל את כל הגנים המקודדים לחלבון, בעוד ש-RNA פולימראז I מתמלל גנים של rRNA, ו-RNA פולימראז III מתמלל גנים rRNA, tRNA וגנים גרעיניים קטנים של RNA. התחלת השעתוק באאוקריוטים כרוכה בקשירה של מספר גורמי שעתוק לרצפי פרומטור מורכבים הממוקמים בדרך כלל במעלה הזרם של הגן המועתק. ה-mRNA מסונתז בכיוון 5' עד 3', וקומפלקס FACT נע ומרכיב מחדש נוקלאוזומים כשהפולימראז עובר. בעוד שפולימראזות RNA I ו-III מפסיקות את השעתוק בשיטות תלויות חלבון או RNA, RNA פולימראז II מתמלל עבור 1,000 נוקלאוטידים או יותר מעבר לתבנית הגן ומבקע את העודפים במהלך עיבוד טרום mRNA.
חיבורי אמנות
איור\(\PageIndex{2}\): מדען מחבר מקדם אוקריוטי מול גן חיידקי ומכניס את הגן לכרומוזום חיידקי. האם היית מצפה שהחיידקים יתמללו את הגן?
- תשובה
-
לא. פרוקריוטים משתמשים במקדמים שונים מאשר אוקריוטים.
הערות שוליים
- 1 H Liang et al., "אבולוציה מהירה של מקדמי ליבה בגנום פרימטים", ביולוגיה מולקולרית ואבולוציה 25 (2008): 1239—44.
רשימת מילים
- תיבת CAAT
- (GGCCAATCT) רצף מקדם אוקריוטי חיוני המעורב בגורמי שעתוק מחייבים
- עובדה
- קומפלקס ש"מקל על שעתוק הכרומטין" על ידי פירוק נוקלאוזומים לפני תמלול RNA פולימראז II והרכבתם מחדש לאחר שהפולימראז עובר
- תיבה עשירה ב- GC
- (GGCG) רצף מקדם אוקריוטי לא חיוני הקושר גורמים סלולריים כדי להגביר את יעילות התעתיק; עשוי להיות נוכח מספר פעמים בפרומוטור
- תיבת אוקטמר
- (ATTTGCAT) רצף מקדם אוקריוטי לא חיוני הקושר גורמים סלולריים כדי להגביר את יעילות התעתיק; עשוי להיות נוכח מספר פעמים בפרומוטור
- קומפלקס ייזום מראש
- מקבץ של גורמי שעתוק וחלבונים אחרים המגייסים RNA פולימראז II לשעתוק של תבנית DNA
- RNA גרעיני קטן
- מולקולות המסונתזות על ידי RNA פולימראז III בעלות מגוון פונקציות, כולל שחבור טרום mRNA וויסות גורמי שעתוק