
15.2: תמלול פרוקריוטי
- Page ID
- 205962

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מיומנויות לפיתוח
- רשום את השלבים השונים בתעתיק פרוקריוטי
- דון בתפקידם של מקדמים בתעתיק פרוקריוטי
- תאר כיצד ומתי מסתיים התמלול
הפרוקריוטים, הכוללים חיידקים וארכאים, הם בעיקר אורגניזמים חד תאיים, שבהגדרה, חסרים גרעינים הקשורים לממברנה ואברונים אחרים. כרומוזום חיידקי הוא מעגל סגור קוולנטית שבניגוד לכרומוזומים אוקריוטיים אינו מאורגן סביב חלבוני היסטון. האזור המרכזי של התא בו שוכן ה- DNA הפרוקריוטי נקרא נוקלאואיד. בנוסף, לפרוקריוטים יש לעתים קרובות פלסמידים בשפע, שהם מולקולות DNA מעגליות קצרות יותר שעשויות להכיל רק גן אחד או כמה גנים. ניתן להעביר פלסמידים ללא תלות בכרומוזום החיידקי במהלך חלוקת התא ולעתים קרובות נושאים תכונות כגון עמידות לאנטיביוטיקה.
שעתוק בפרוקריוטים (ובאוקריוטים) מחייב את הסליל הכפול של ה-DNA להירגע חלקית באזור סינתזת ה-mRNA. אזור ההתרה נקרא בועת שעתוק. התעתיק תמיד יוצא מאותו גדיל DNA עבור כל גן, הנקרא גדיל התבנית. תוצר ה-mRNA משלים לגדיל התבנית והוא כמעט זהה לגדיל ה-DNA השני, הנקרא גדיל nontemplate. ההבדל היחיד הוא שב- mRNA, כל נוקלאוטידים T מוחלפים בנוקלאוטידים U. בסליל כפול של RNA, A יכול לקשור U באמצעות שני קשרי מימן, בדיוק כמו בזיווג A — T בסליל כפול של DNA.
צמד הנוקלאוטידים בסליל הכפול של ה-DNA המתאים לאתר שממנו מתועתק נוקלאוטיד ה-mRNA הראשון של 5' נקרא אתר +1, או אתר ההתחלה. נוקלאוטידים שקדמו לאתר ההתחלה מקבלים מספרים שליליים והם מיועדים במעלה הזרם. לעומת זאת, נוקלאוטידים העוקבים אחר אתר ההתחלה מסומנים במספור "+" ונקראים נוקלאוטידים במורד הזרם.
התחלת תמלול בפרוקריוטים
לפרוקריוטים אין גרעינים סגורים בקרום. לכן, תהליכי התעתיק, התרגום ופירוק ה- mRNA יכולים להתרחש כולם בו זמנית. ניתן להגביר במהירות את הרמה התוך תאית של חלבון חיידקי על ידי אירועי שעתוק ותרגום מרובים המתרחשים במקביל באותה תבנית DNA. שעתוק פרוקריוטי מכסה לעתים קרובות יותר מגן אחד ומייצר mRNA פוליציסטרוני המציינים יותר מחלבון אחד.
הדיון שלנו כאן ידגים שעתוק על ידי תיאור תהליך זה ב Escherichia coli, מין חיידקי שנחקר היטב. למרות שקיימים כמה הבדלים בין שעתוק ב אי - קולי ותעתיק בארכאה, ניתן ליישם הבנה של שעתוק E. coli כמעט על כל מיני החיידקים.
פולימראז RNA פרוקריוטי
פרוקריוטים משתמשים באותו פולימראז RNA כדי לתמלל את כל הגנים שלהם. ב אי - קולי, הפולימראז מורכב מחמש יחידות משנה של פוליפפטיד, שתיים מהן זהות. ארבע מיחידות המשנה הללו, המסומנות α, α, β ו- β' מהוות את אנזים הליבה של הפולימראז. יחידות משנה אלה מתאספות בכל פעם שגן מתועתק, והן מתפרקות לאחר השלמת התעתיק. לכל יחידת משנה יש תפקיד ייחודי; שתי יחידות המשנה α נחוצות להרכבת הפולימראז על ה-DNA; יחידת המשנה β נקשרת לטריפוספט הריבונוקלאוזיד שיהפוך לחלק ממולקולת ה-mRNA המתהווה "שנולדה לאחרונה"; וה-β' קושר את גדיל תבנית ה-DNA. יחידת המשנה החמישית, σ, מעורבת רק בהתחלת שעתוק. זה מקנה ספציפיות תעתיק כך שהפולימראז מתחיל לסנתז mRNA מאתר התחלה מתאים. ללא σ, אנזים הליבה יתמלל מאתרים אקראיים וייצר מולקולות mRNA שציינו ג'יבריש חלבון. הפולימראז המורכב מכל חמש יחידות המשנה נקרא הולואנזים.
מקדמים פרוקריוטיים
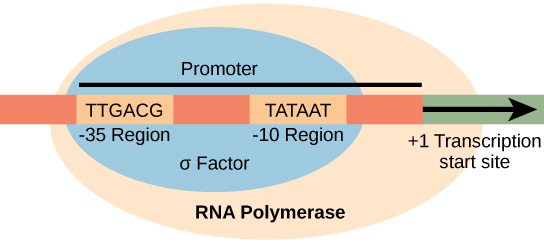
מקדם הוא רצף DNA שאליו נקשרת מנגנון השעתוק ויוזם שעתוק. ברוב המקרים, מקדמים קיימים במעלה הזרם של הגנים שהם מווסתים. הרצף הספציפי של מקדם חשוב מאוד מכיוון שהוא קובע אם הגן המתאים מועתק כל הזמן, חלק מהזמן או לעתים רחוקות. למרות שמקדמים משתנים בין הגנום הפרוקריוטי, כמה אלמנטים נשמרים. באזורים -10 ו-35 במעלה הזרם של אתר ההתחלה, ישנם שני רצפי קונצנזוס פרומטור, או אזורים דומים בכל המקדמים ועל פני מיני חיידקים שונים (איור). \(\PageIndex{1}\) רצף הקונצנזוס -10, הנקרא אזור -10, הוא TATAAT. רצף -35, TTGACA, מזוהה ומחויב על ידי σ. לאחר יצירת אינטראקציה זו, יחידות המשנה של אנזים הליבה נקשרות לאתר. האזור העשיר ב- A- T -10 מקל על פירוק תבנית ה- DNA, ונוצרים מספר קשרי פוספודיסטר. שלב התחלת השעתוק מסתיים בייצור תמלילי הפלה, שהם פולימרים של כ-10 נוקלאוטידים המיוצרים ומשתחררים.
קישור ללמידה
צפה באנימציה זו של MolecularMovies כדי לראות את החלק הראשון של התמלול ואת החזרה על רצף הבסיס של תיבת TATA.
התארכות וסיום בפרוקריוטים
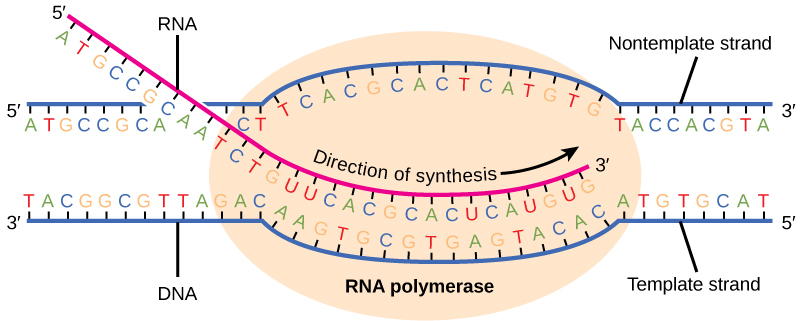
שלב התארכות השעתוק מתחיל בשחרור תת-היחידה σ מהפולימראז. הדיסוציאציה של σ מאפשרת לאנזים הליבה להמשיך לאורך תבנית ה-DNA, לסנתז mRNA בכיוון 5' עד 3' בקצב של כ-40 נוקלאוטידים בשנייה. ככל שההתארכות מתקדמת, ה-DNA מתפרק ברציפות לפני אנזים הליבה ומתפתל מאחוריו (איור). \(\PageIndex{2}\) זיווג הבסיס בין DNA ל- RNA אינו יציב מספיק כדי לשמור על יציבות רכיבי סינתזת ה- mRNA. במקום זאת, ה-RNA פולימראז פועל כמקשר יציב בין תבנית ה-DNA לבין גדילי ה-RNA המתהווים כדי להבטיח שההתארכות לא תופרע בטרם עת.
אותות סיום פרוקריוטיים
לאחר שעתוק גן, יש להנחות את הפולימראז הפרוקריוטי להתנתק מתבנית ה-DNA ולשחרר את ה-mRNA החדש שנוצר. בהתאם לגן המתועתק, ישנם שני סוגים של אותות סיום. האחד מבוסס על חלבון והשני מבוסס RNA. סיום תלוי Rho נשלט על ידי חלבון ה- rho, העוקב מאחורי הפולימראז בשרשרת ה- mRNA הגדלה. לקראת סוף הגן, הפולימראז נתקל בריצה של נוקלאוטידים G על תבנית ה-DNA והוא נעצר. כתוצאה מכך, חלבון ה- rho מתנגש בפולימראז. האינטראקציה עם rho משחררת את ה- mRNA מבועת התעתיק.
סיום בלתי תלוי ב- Rho נשלט על ידי רצפים ספציפיים בגדיל תבנית ה- DNA. כאשר הפולימראז מתקרב לסוף הגן המתועתק, הוא נתקל באזור עשיר בנוקלאוטידים C-G. ה-mRNA מתקפל בחזרה על עצמו, והנוקלאוטידים C-G המשלימים נקשרים יחד. התוצאה היא סיכת שיער יציבה הגורמת לפולימראז להיעצר ברגע שהוא מתחיל לתמלל אזור עשיר בנוקלאוטידים A-T. אזור U- A המשלים של תמליל ה- mRNA יוצר רק אינטראקציה חלשה עם ה- DNA של התבנית. זה, יחד עם הפולימראז שנתקע, גורם לחוסר יציבות מספיק כדי שאנזים הליבה יתנתק וישחרר את תמליל ה-mRNA החדש.
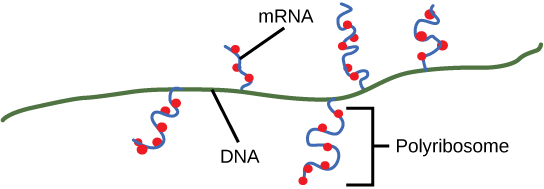
עם סיום, תהליך התמלול הושלם. עד לסיום, התמליל הפרוקריוטי כבר היה משמש כדי להתחיל בסינתזה של עותקים רבים של החלבון המקודד מכיוון שתהליכים אלה יכולים להתרחש במקביל. האיחוד של שעתוק, תרגום ואפילו פירוק mRNA אפשרי מכיוון שכל התהליכים הללו מתרחשים באותו כיוון 5' עד 3', ומכיוון שאין מידור קרומי בתא הפרוקריוטי (איור). \(\PageIndex{3}\) לעומת זאת, נוכחותו של גרעין בתאים אוקריוטיים מונעת שעתוק ותרגום בו זמנית.
קישור ללמידה
בקר באנימציה זו של BioStudio כדי לראות את תהליך התעתיק הפרוקריוטי.
סיכום
בפרוקריוטים, סינתזת mRNA מתחילה ברצף פרומטור בתבנית ה-DNA הכוללת שני רצפי קונצנזוס המגייסים RNA פולימראז. הפולימראז הפרוקריוטי מורכב מאנזים ליבה של ארבע יחידות משנה של חלבון וחלבון σ המסייע רק בהתחלה. התארכות מסנתזת mRNA בכיוון 5' עד 3' בקצב של 40 נוקלאוטידים לשנייה. סיום משחרר את ה- mRNA ומתרחש על ידי אינטראקציה של חלבון rho או על ידי יצירת סיכת ראש mRNA.
רשימת מילים
- הסכמה
- רצף DNA המשמש מינים רבים לביצוע פונקציות זהות או דומות
- אנזים ליבה
- פולימראז RNA פרוקריוטי המורכב מ-α, α, β ו-β' אך חסר σ; קומפלקס זה מבצע התארכות
- במורד הזרם
- נוקלאוטידים העוקבים אחר אתר ההתחלה לכיוון שעתוק mRNA; באופן כללי, רצפים שנמצאים לקראת סוף 3 'ביחס לאתר ב- mRNA
- סיכת ראש
- מבנה ה- RNA כאשר הוא מתקפל על עצמו ויוצר קשרי מימן תוך מולקולריים בין נוקלאוטידים משלימים
- הולואנזים
- פולימראז RNA פרוקריוטי המורכב מ-α, α, β, β' ו-σ; קומפלקס זה אחראי לתחילת השעתוק
- אתר חניכה
- נוקלאוטיד שממנו סינתזת mRNA ממשיכה בכיוון 5 'עד 3'; מסומן עם "+1"
- גדיל שאינו תבנית
- גדיל DNA שאינו משמש לתמלול mRNA; גדיל זה זהה ל-mRNA פרט לכך שנוקלאוטידים T ב-DNA מוחלפים בנוקלאוטידים U ב-mRNA
- פלסמיד
- מולקולת DNA מעגלית אקסטרכרומוזומלית, סגורה קוולנטית, שעשויה להכיל רק גן אחד או כמה גנים; נפוץ בפרוקריוטים
- מקדם
- רצף DNA שאליו RNA פולימראז וגורמים קשורים נקשרים ומתחילים שעתוק
- סיום תלוי Rho
- בפרוקריוטים, הפסקת שעתוק על ידי אינטראקציה בין RNA פולימראז לחלבון rho בריצה של נוקלאוטידים G בתבנית ה-DNA
- עצמאי Rho
- סיום תלוי רצף סיום של סינתזת mRNA פרוקריוטית; נגרמת על ידי היווצרות סיכת ראש ב-mRNA שעוצרת את הפולימראז
- תיבת טאטא
- רצף פרומטור שמור באאוקריוטים ופרוקריוטים המסייע לבסס את אתר ההתחלה לתעתוק
- גדיל תבנית
- גדיל DNA המציין את מולקולת ה- mRNA המשלימה
- בועת תמלול
- אזור של DNA לא מפותל מקומי המאפשר שעתוק של mRNA
- במעלה הזרם
- נוקלאוטידים שקדמו לאתר החניכה; באופן כללי, רצפים לקראת קצה 5' ביחס לאתר ב- mRNA