
7.4: גודל וצורה
- Page ID
- 207513

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)הסוג הסופי של דפוס התפתחותי שהאבולוציה יכולה לפעול עליו הוא הגודל והצורה של רקמות או איברים. אלה נחשבים בדרך כלל לבעיות קנה מידה "מורפומטריות" ומסווגים כשינויים "אלומטריים". מורפומטריה היא המחקר כיצד ניתן לעוות גיאומטריה רציפה (כמו המשטח החיצוני של גוף). אלומטריה חוקרת זאת בהקשר של אבולוציה והתפתחות. אחד האנשים שהגדירו את השדה, ד'ארסי וונטוורת 'תומפסון, העלה שינויים מורפומטריים המאפשרים עיוות של מין דגים אחד ידוע למין ידוע אחר (איור 7).
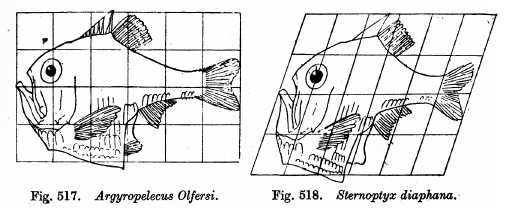
אמנם היקף התפקיד שממלא האלומטריה באבולוציה אינו ברור לחלוטין, אך אנו יודעים מדוגמאות שהיא אכן ממלאת תפקיד כלשהו. קל לראות זאת אם נסתכל סביב על מינים קשורים. לדוגמה, גולגולות הומינידיות דומות מאוד ולעתים קרובות שונות בדרכים שמוסברות בקלות על ידי וריאציות קטנות בדפוסי הצמיחה ההתפתחותיים. התפשטות תאים נוספת או מוקדמת יותר כאן, פחות או מאוחר יותר התפשטות תאים שם. למרות שאנו יודעים שסמנים גנטיים רבים של התפשטות תאים (למשל מווסת מחזור התא Cyclin D1) מגבירים את התפשטות התאים, ישנם מפעילים רבים ושונים במעלה הזרם של התפשטות בהתאם לרקמה/סוג התא ולשלב ההתפתחותי. כדי להתחיל להבין את תפקידה של האלומטריה באבולוציה, חוקרים משווים את צמיחת הרקמות ואת הרגולטורים של צמיחה זו בין מינים קרובים למדי.
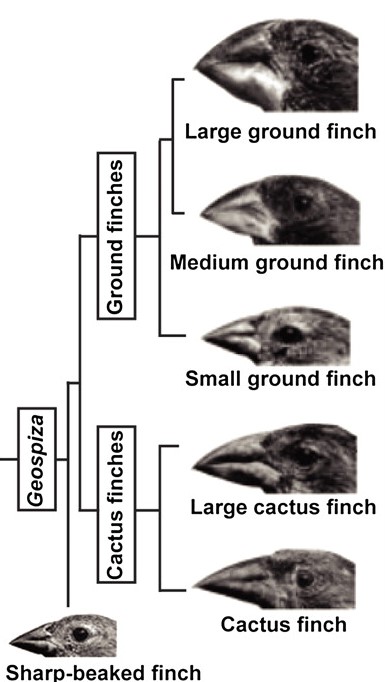
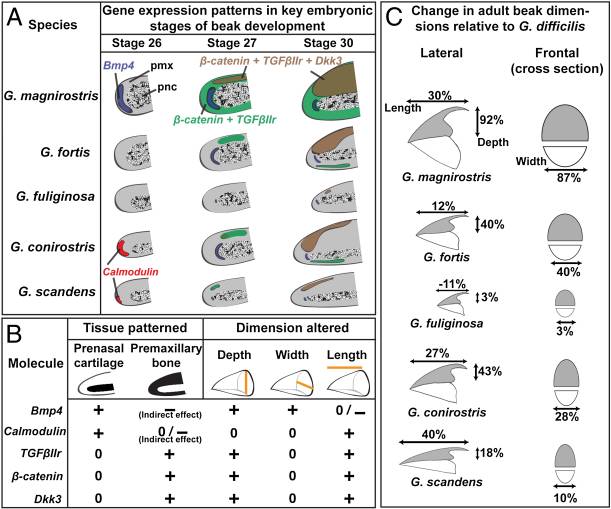
שני מקומות שבהם ההתפתחות והאבולוציה של שינויים בגודל ובצורה מובנים למדי הם צורות מקור בציפורים ואליטרה בחיפושיות. אחת הדוגמאות המפורסמות ביותר לאבולוציה היא החוחיות של דרווין. קבוצת חוחיות זו עברה קרינה אדפטיבית באיי גלפגוס החל לפני כ -2.3 מיליון שנה 12. מאז, החוחיות הסתגלו לנישות אי שונות ל -14 מינים שונים. השוואת מורפולוגיה של מקור בין מינים אלה חשפה שתי תוכניות אלומטריות שמורות לשינוי דפוסי התפשטות תאים. בהתפתחות מוקדמת, רוחב המקור מוסדר על ידי רמות BMP4 - BMP4 גבוה יותר פירושו מקור עמוק ורחב יותר. לדוגמה, לחוחיות קרקע יש ביטוי BMP4 גבוה באזור המסמן לאזור ההחלקה השולט של המקור המוקדם - הסחוס שלפני האף (pnc) 13. באופן דומה, רמות גבוהות של קלמודולין ליד ה-pnc קשורות למקור ארוך יותר, כמו מקורם של חוחיות קקטוס (איור 8).
מעט מאוחר יותר בהתפתחות, מבנה מקור עליון הנקרא עצם טרום מקסילרי (pmx) מתחיל לצמוח. זה יהווה בסופו של דבר את חלקי המקור העליון הנבדלים מבחינה מבנית ותפקודית בין מיני ציפורים רבים. חלק זה של המקור מבטא גם קבוצות שונות של גנים בצורות מקור שונות. קולטן TGFβ סוג II (TGFβIIR), β-catenin ו- Dickkopf-3 (Dkk3) הם כולם גנים של מפל העברת אותות המתבטאים ברמות גבוהות בהרבה במקור החוחית הקרקעית הגדולה בהשוואה לחוחיות מקוריות קטנות יותר. שלושת הגנים הללו באו לידי ביטוי בתחומים רחבים יותר במקור גדול יותר מאשר במקור קטן יותר כאשר מסתכלים על כל חמשת המינים באיור למעלה (איור 9) 14.
ממצאים אלה העלו את הרעיון שצמיחה אלומטרית יכולה לתדלק אבולוציה מהירה על ידי שינוי קנה המידה של תכונה מסוימת. שינוי הסולם מתייחס רק לקנה מידה של משהו למעלה או למטה, מה שהופך אותו לגדול או קטן יותר בממד אחד או בכל הממדים. לדוגמה ריבוע גדול הוא פשוט גרסה מוגדלת של ריבוע קטן. מלבן יהיה ריבוע שגודלו בקנה מידה גדול יותר בממד אחד אך לא בשני. חוקרים הבחינו מיד שפשוט שינוי קנה המידה של המקור העליון יכול להסביר הרבה מהשונות בגיאוספיזה וזה יכול להיות מוסבר על ידי מערכת הקואורדינטות של הגנטיקה שלמעלה (או אפילו מערכת גנטית פשוטה יותר). עם זאת, קנה המידה לא הספיק כדי להסביר את השונות במיני חוחית קשורים אחרים. כדי להסביר את השונות הזו, החוקרים מצאו כי יש לשקול גם קנה מידה וגם גזירה. גזירה מתייחסת לשינוי גיאומטרי כמו זה שנראה בדוגמת הדגים מאת Thompson et al. מֵעַל. בשינוי גזירה, כל נקודה נעה לאורך x במרחק פרופורציונלי לקואורדינטת y שלה (או להיפך) נותנת קו אלכסוני מקו ישר. גזירה פלוס קנה מידה הספיקה כדי להסביר לפחות את צירי האורך והעומק של מקור החוחית שנחקרו (איור 10)
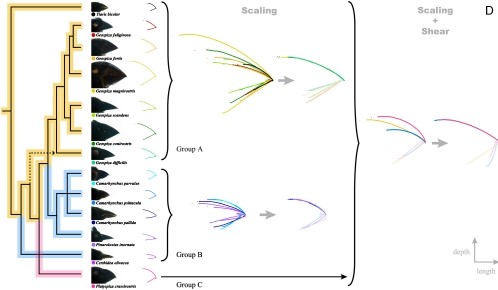
השאלה הבאה שיש לענות עליה היא עד כמה מערכת הקואורדינטות הזו ישימה באופן נרחב לשינוי צורת המקור? הגדלת או הפחתה של רמות הביטוי של גנים אלה אכן משנה את צורת מקור האפרוח בדרכים שחזו Mallarino et al. 14. מידע זה עשוי להוביל אותנו לחזות שהאבולוציה תשתמש במערכת קואורדינטות זו כדי ליצור את המגוון הרחב של המקור שאנו רואים בציפורים חיות ומאובנות. עם זאת, מחקרים על חוחיות שאינן Geospiza מראים שהאבולוציה משתנה יותר ממה שאנחנו יכולים לדמיין. אם יהיה לנו זמן, נדון במאמר: "מיני ציפורים קרובים מפגינים גמישות בין מורפולוגיה של מקור לתוכניות התפתחות בסיסיות".