
4.8: 我们的远古过去-最早的人类
- Page ID
- 204682

在本节结束时,您将能够:
用两只脚行走
人参一词是指所有被认为与人类有直接谱系的物种,包括人类属、南方古猿属、P aranthropus 属和 Ardipith ecus 属。 人类是指所有现代和灭绝的大猿,包括人类、大猩猩、黑猩猩和猩猩及其祖先。 多年来,人们认为这些术语代表不同的东西,但此处提供的定义是最新的。 尽管所有的人参素可能在不同方面有所不同,但它们都具有一个解剖学行为复合体:双足运动。
科学家可以通过分析生物形态的多个方面来假设生物是如何移动的。 Brachiators 是通过在树枝之间摆动来移动的动物,通常手臂很长,而跳跃者是通过下肢力量推动身体的动物,腿相对较长。 树栖灵长类动物的手臂和腿的长度相等。 在双足运动中,一条腿被称为姿态腿,另一条腿被称为踏步腿。 当姿态腿在地面上时,阶梯腿离开地面向前迈进。 在正常行走过程中,只有大约 25% 的时间双脚都在地面上。 随着运动速度的提高,双脚在地面上的时间百分比会降低。 因此,在双足生物移动的大部分时间里,它们的身体只有一条腿(姿态腿)保持平衡。 为了确保双足生物在保持姿势平衡时不会摔倒,自最早的人类祖先以来,它们经历了许多解剖学变化。
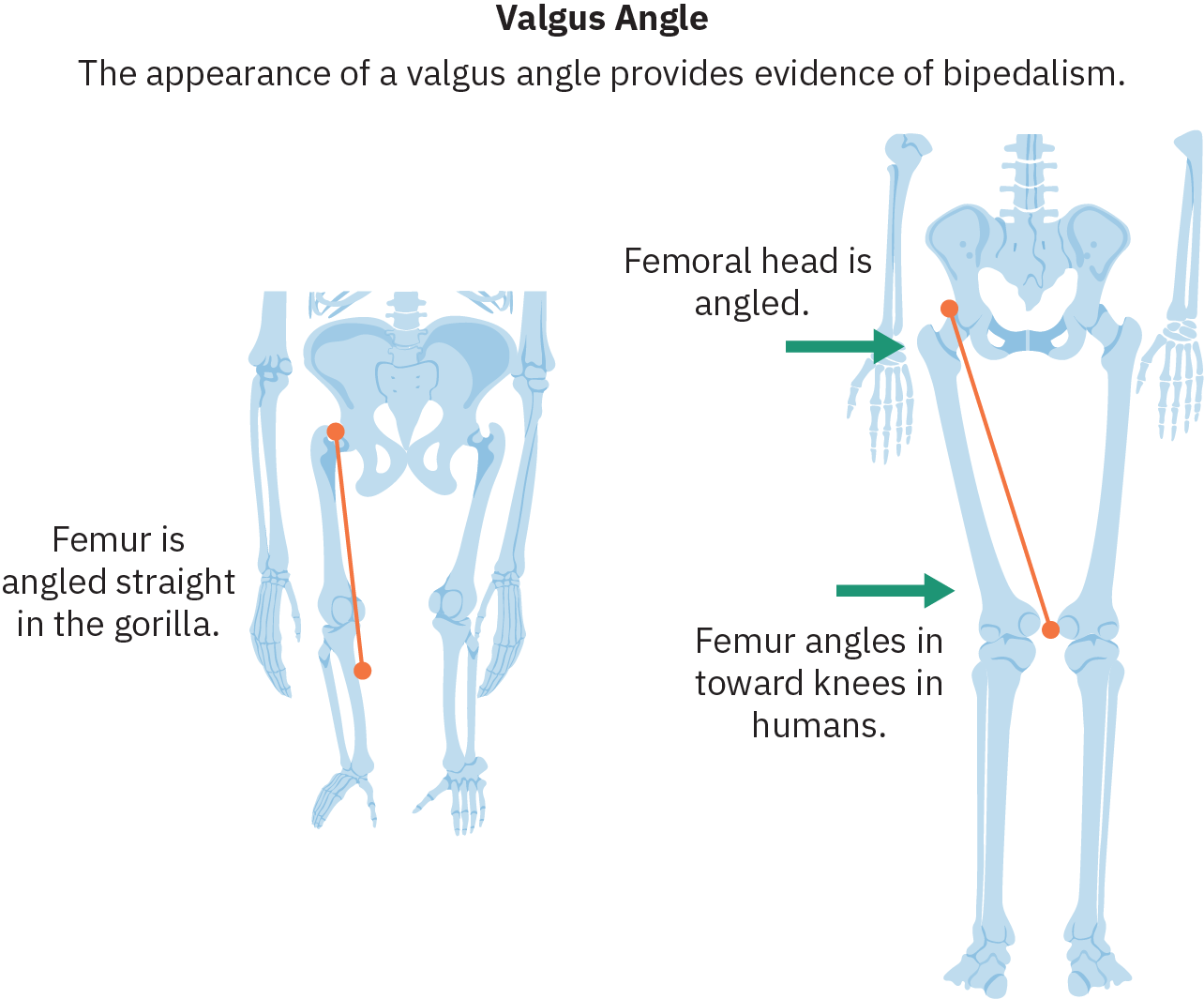
促进成功进行双足运动的最重要的解剖学变化之一是股骨(大腿骨)以所谓的外翻角向内倾斜,将膝盖和脚置于骨盆中心以下。 双足人参素还进化出脊柱曲线,使臀部有可能平衡上半身的重量。 足弓的演变以及大脚趾的重新调整使其与其他脚趾平行,也有助于在双足运动的步步阶段传递重量。
英国古人类学家玛丽·利基(Mary Leakey)的著作提供了早期人参两足动物的最重要证据。 20 世纪 80 年代,玛丽·利基发现了一条 75 英尺长的足迹,这些足迹由三只双足动物在大约 3.5 MYA 的湿火山灰层中穿过。 这些足迹是在东非的莱托利遗址发现的。 根据日期和地点,这些足迹很可能是南方古猿阿法伦西斯制作的。 对Laetoli足迹的分析表明了现代的步态。

人参素两足动物的演变需要复杂的解剖学重组。 要使自然选择产生如此巨大的变化,这些变化的好处肯定是巨大的。 这些变化有几十种假设,从腾出双手携带工具、食物或后代,到通过暴露更多人体表面来提高能效或体温调节(维持人体温度的能力)。 所有这些假设都无法检验,这使得理解人类祖先为何做出如此巨大的行为转变确实具有挑战性。 接下来的章节将探讨早期人参化石的一些关键发现,在这些发现中,人类学家看到了人类故事中两足动物适应的最早迹象。
中新世原始人
第一批人类化石出现在中新世晚期,即 10 至 5 MYA。 在 7 MYA 到 4 MYA 之间的某个时候,原始人从树上移出,开始更充分地适应地面生活利基市场。 不幸的是,这段时期的化石证据极为稀少,但新的发现仍在继续。
2002 年,法国古人类学家米歇尔·布鲁内特及其团队在西非乍得发现了完整的 Sahelanthropus tchadensis 颅骨。 Sahelanthropus 是一种化石猿,活了大约 7 MYA,一些研究人员声称它是人类和黑猩猩的最后共同祖先。 遗传研究表明,人类和黑猩猩在5 MYA和7 MYA之间存在分歧,因此该物种在分歧时正好生活。 颅骨容量仅为350立方厘米(cc),相当于黑猩猩;现代人类的颅骨容量约为1,400 cc。 Sahelanthropus 还有一个非常大的眉脊(眼睛上方的大骨头),而孔大酒瓶的位置,即脊柱进入头骨的头部开口,表明它的头部没有被固定在脊柱上,因此它不是双足动物。
Orrorin tugenensis 是法兰西学院的地质学家马丁·皮克福德和法国国家自然历史博物馆的古生物学家布里吉特·塞努特于 2001 年在肯尼亚发现的。 Orrorin tugenensis 的历史可追溯到大约 6 MYA。 Orrorin 之所以被提议为人参素,是因为解剖学特征暗示了两足动物。 例如,股骨头(位于腿骨顶部的连接腿部和臀部的大圆球)比四足猿大得多,这表明股骨被用来支撑上半身的重量。 附着在股骨上的肌肉也提示双足运动。 另一个表明 Orrorin 确实是人参素的特征是牙齿,它表现出厚厚的牙釉质和小的方形磨牙,就像现代人类一样。
上新世人参素
上新世时代从 5 MYA 延伸到 1.8 MYA。 来自上新世的化石显示了显然是双足的人类素进化的证据。 它们还显示了明确但原始的文化行为的证据。 从气候上讲,上新世比之前的中新世更冷,这导致海平面变化和两极冰层增加,打开了一些以前无法进入的地区。 在此期间,北美和南美通过巴拿马地峡连接起来,阿拉斯加和西伯利亚之间出现了一座横跨白令海峡的陆桥。
Ardipithecus ramidus
美国古人类学家蒂姆·怀特于 1992 年在埃塞俄比亚发现了 Ardipithecus ramidus,其历史可追溯到大约 4.4 MYA。 这是可追溯到上新世时代的第一个发现的人参物种。 根据孔大酒瓶的向前位置,可以得出结论 Ardipithecus 是双足动物。 此外,上臂骨头很小,这表明手臂在运动过程中没有用来支撑重量。 Ardipithecus 具有许多特征,例如牙釉质薄、犬类减少的证据和对立的大脚趾。 由于后一种特征,许多人认为 Ardipithecus 在地面上是双足动物,在树上是四足动物。 这一假设得到了以下事实的支持:化石骨骼是在森林相对密集的环境中发现的。 缩小犬是一种衍生特征,出现的时间甚至比 A. ramidus 还要早,这不是我们通常在拥有大型恐吓犬类的非洲猿雄性身上看到的那样。 目前的假设表明,随着时间的推移,当人们不再需要表现出攻击性并且雌性偏爱性质较温和的雄性时,较小的犬类就会占主导地位(Suwa,G. 等人,2021)。

坚固而亲切的南方古猿
接下来的几节将研究具有与牙齿和头骨形态相关的不同物理特征的各种南方古猿物种。 根据这些特征,古人类学家将这些物种分为温柔而健壮的形态,如图 4.33 所示。 Gracile 物种的下巴投射更明显(prognathism),张开的脸颊较少,没有矢状波峰,牙齿和下巴较小。 坚固的南方古猿中的矢状波峰可容纳较大的颌骨肌肉,用于咀嚼坚硬的植物材料。
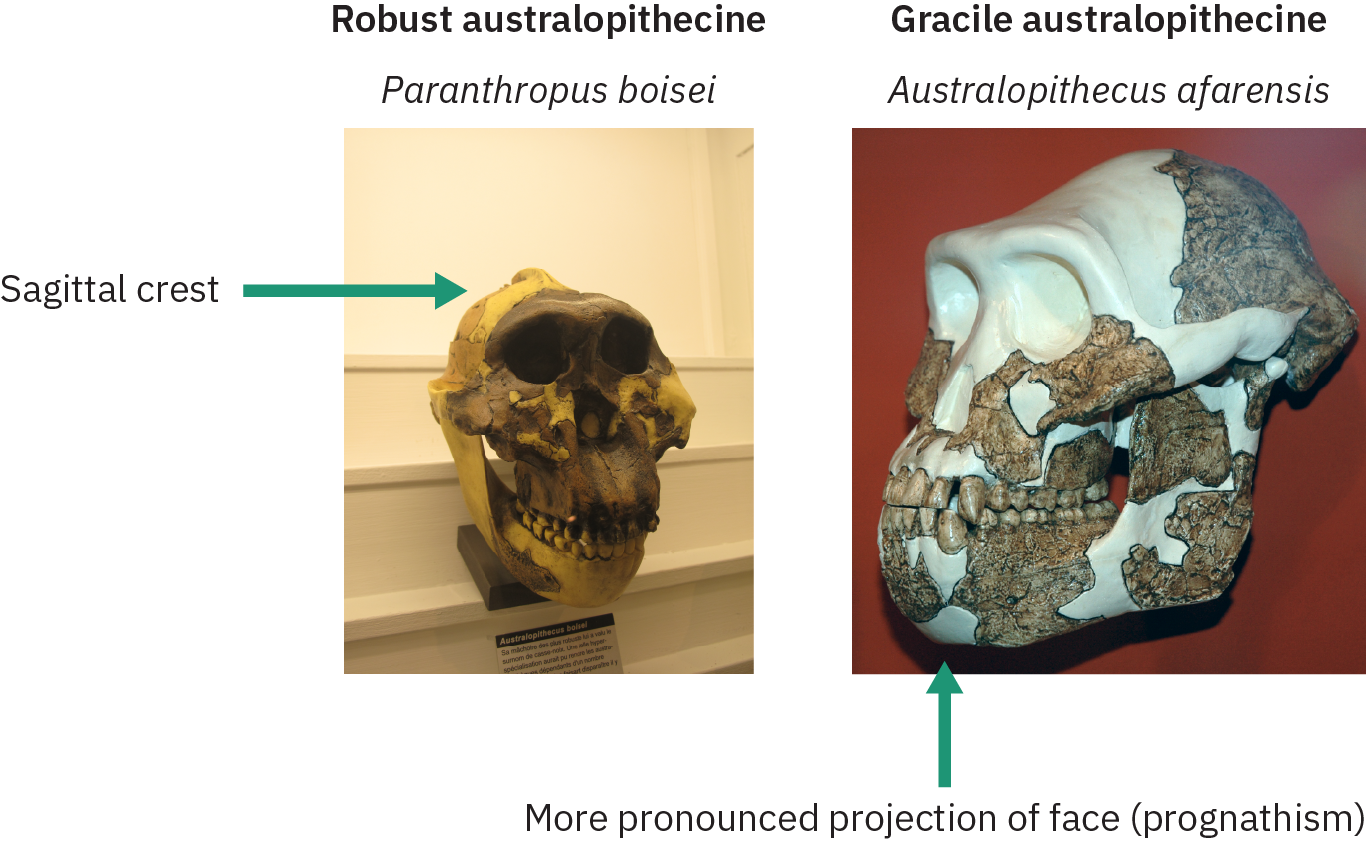
被认为是 gracile 的物种包括 A ustralopithecus anamensis、A. afransis、A. africanus、A. garhi 和 A. sediba。 强大的南方古猿(归类为 Paranthropus 属)包括 Paran thropus robustus、P. boisei 和 P. aethiopicus。 温柔的物种出现在 4 MYA 左右,消失了 2 MYA,而健壮的物种又存在了一百万年。 接下来的章节将首先介绍南方古猿的一些温和形式,然后是强壮的形式。
Australopithecus africanus
Australopithecus africanus 是 1924 年发现的第一个南方古猿,澳大利亚解剖学家和人类学家雷蒙德·达特对此进行了描述,他在南非一个名为 Taung 的石灰采石场工人寄给他的一盒化石中发现了这种化石。 盒子里最引人注目的标本是一个孩子的头骨,达特必须将其从嵌入的石头上砍掉。 达特花了四年时间才把牙齿分开。 头骨现在被称为 Taung 头骨或 Taung 小孩。 达特认为,唐的孩子代表着 “介于活类人猿和人类之间的灭绝类人猿种族”(Wayman 2011)。 他指出,头骨又长又窄,不像现代人类那样圆润,它的大脑平均只有422 cc,相当于黑猩猩。 但是,Taung 的孩子没有眉脊,有圆形的轨道,前体力很小,犬也很小,没有腹泻(闭上嘴巴时下巴有空间可以放置大型犬类)。 后面的这些特征都类似于现代人类。 最重要的是,达特指出,孔大酒瓶的向前位置表明头骨处于椎柱的顶部,这表明两足动作和直立姿势。

南方古猿 afarensis
1973年,美国古人类学家唐纳德·约翰逊在埃塞俄比亚的阿法尔地区发现了很大一部分骨骼(约40%)。 他用甲壳虫乐队的一首歌叫骷髅露西。 它的历史可追溯到大约 3.75—2.8 MYA,被确定为南方古猿 afarensis 物种的成员。 像最近发现的所有化石一样,露西被赋予了一个识别号或入藏号 KNM-AL-288。 KNM 的首字母缩写代表存放化石的肯尼亚国家博物馆,AL 代表发现化石的阿法尔地区。 从那时起,在肯尼亚、坦桑尼亚和埃塞俄比亚以及东非发现了更多该物种的标本。

A@@ ustralopithecus afarensis 的年代为 3.9 至 2.9 MYA,颅内容量约为 400 cc,与普通黑猩猩大致相同。 有两个形态学特征提供了证据,表明 A. afarenis 的移动方式更像是大猿而不是人类。 首先,它的手臂比现代人类长得多。 长臂通常出现在悬挂在树枝上的动物身上,这表明 A. afarensis 也表现出这种行为。 此外,A. afarensis 拥有长而弯曲的手指和脚趾骨,这是悬挂在树枝上的动物的另一个特征。 但是,A. afarensis 有一个重要的形态学特征表明该物种的移动可能有点像现代人类。 A. afarens is 的骨盆(臀骨)的形状看起来更像现代人的骨盆,而不是猿的骨盆。髋骨不是又长又窄,而是又短又宽。 大多数古人类学家认为,骨盆形状的这种变化表明 A. afarensis 像现代人类一样用两条腿移动。 虽然 A. afarensis 可能是双足运动的,但 A. afarensis 和现代人类之间的形态差异表明它们的移动方式并不完全相同。 目前的共识是,A. afarensis 既是树栖又是双足动物。 两足动物的其他解剖学证据包括孔大酒瓶更靠前的位置以及股骨头部和颈部的角度。
南方古猿 garhi
同样在埃塞俄比亚发现的 Australopithecus garhi 的历史可追溯到大约 2.5 MYA。 它的颅骨容量略高于 A. afarensis,为 450 cc。 Australopithecus garhi 的门牙比任何已知的南方古猿或 Homo 的门牙都要大。 大型门牙的功能尚不清楚。 A. garhi 最令人兴奋的方面是,它提供了人类最早使用石器工具的证据。 具体而言,A. garhi 化石被发现带有反刍动物(例如羚羊)的化石骨头,上面有许多切割痕迹。 切割痕迹是通过用石头或金属工具去除骨头上的肉的过程在骨头上留下的。 基于这一发现,生物人类学家假设 A. garhi 使用了某种类型的石器进行屠杀。
南方古猿 sediba
2008 年,美国古生物学家李·伯杰的九岁儿子马修·伯杰在南非马拉帕发现了南方古猿塞迪巴的锁骨骨。 在洞穴特征中进一步挖掘发现了两个部分骨骼,一个是成年雌性,另一个是年幼的少年。 A. sediba 被认为是重要的物种,因为它出现在化石记录中,大约在 2 mya 左右 Homo 属首次出现的时候。 A. sediba 的分类最初很难确定,因为它具有复杂的重叠特征,包括类似人类的脊柱、骨盆、手和牙齿以及类似黑猩猩的脚。 这种特征的组合表明了爬树和双足动物的适应能力。 在集体研究了这些特征之后,人类学家将 A. sediba 归类为南方古猿的一种。 它被认为是直立人和 Homo ergaster 的直接祖先,这两者将在第 5 章《人属与我们的出现》中讨论。 据信 A. sediba 可能是 A. africanus 的后代,这表明该物种可能是人类谱系中的死胡同。 它的分类和与 Homo 属的关系可能仍将备受争议。
Paranthropus robustus
雷蒙德·达特被发现十三年后,南非古生物学家兼医生罗伯特·布鲁姆在南非一个名为 K romdraai 的地点发现了 Paranthropus robust us 。 达特和布鲁姆各自的化石 A. africanus 和 P. robustus 之间最明显的区别在于 Broom 化石的形态要大得多。 它的特点包括矢状波峰和喇叭状的颧弓,用于附着一块巨大的腱骨,用于咀嚼依赖硬坚果和种子的饮食。 这种解释得到了扫描电子显微镜(SEM)的进一步支持,扫描电子显微镜(SEM)用于评估蚀刻在牙齿上的标记。 随着牙齿大小的增加,门牙和犬类会缩小,这使 Paranthropus 的脸更平坦,下巴的投射更少。 有人认为,根据环境和地点的不同,有些 Paranthropus 可能是杂食动物,其饮食与 H. ergaster 的饮食相似。 (Lee-Thorp、Thackeray 和 van der Merwe 2000)。
Paranthropus boisei
跟随布鲁姆的脚步,其他科学家开始在东非寻找化石。 从1931年开始,肯尼亚和英国古人类学家路易斯·利基和他的妻子玛丽·利基在所谓的东裂谷工作,这是一个横跨埃塞俄比亚、肯尼亚和坦桑尼亚的1200英里低谷。 他们搜寻了将近 30 年才在 1959 年发现了他们的第一个人参化石 P aranthropus boisei(OH-5)(最初被归类为 Zinjanthropus boisei)。 它通常被称为超强壮的人参素,因为它的头骨顶部有莫霍克族的骨头。 其他特征包括额头低或缺失、脸部扁平、下巴大,以及整个头骨上有较大的附着部位,便于咀嚼肌肉。
Paranthropus aethopicus
我们对P aranthropus aethiopicus(如图 4.37 所示)知之甚少,它的历史可追溯到大约 2.5 MYA,被称为 “黑骷髅”。 据信,该物种介于强壮和温柔的南方古猿之间,具有两者的特征。 该物种是1967年由卡米尔·阿兰堡和伊夫·科本斯率领的法国探险队在埃塞俄比亚发现的。

地标和问题
尽管到目前为止发现的化石为了解人类过去的故事提供了一个小窗口,但它们同时也提出了许多问题。 与系统发育关系和分歧点有关的问题是古人类学家面临的挑战,他们只有零散的化石证据可以围绕这些假设进行假设。 尽管如此,已经取得的发现代表了人类学家理解中的重要里程碑,为人类旅程的下一步提供了线索。
迷你实地考察活动
行人调查
进行行人调查,尝试在你居住的地方附近找到化石(纽约的三叶虫、德克萨斯州的炸药、河床附近的鲨鱼牙齿、箭头)。 想想你最有可能在哪里找到化石以及为什么。 尝试在不破坏周围环境的情况下提取一个,这提供了重要的背景信息。 尝试通过一些互联网研究来弄清楚它是什么样的化石。 你为什么认为这个化石被保存下来? 哪些信息会使寻找化石变得更容易?