
4.7: 灵长类动物的起源和分类
- Page ID
- 204646

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)在本节结束时,您将能够:
理解时间概念
地质学家将深厚的历史划分为称为时代的时期。 时代通常以观察到的化石生命形式为基础。 最古老的地质时代是Eoarchean,它始于大约四十亿年前。 我们掌握的灵长类动物进化的大部分化石证据来自新生代时代——当前的地质时代,其历史可追溯至6500万年前(MYA)至今。 新生代分为一系列时代。 每个时代都与在该时期进化的特定形式的灵长类动物有关。
化石和定年方法
生物人类学家主要研究化石文物,但不限于研究化石文物。 化石是指保存在地球中的植物或动物的任何剩余物。 生物死亡后,它的身体会慢慢分解,直到剩下的只是牙齿和骨头,或者仅仅是对该生物体原始形态的印象。 在大多数情况下,牙齿和骨骼以及印象最终也会恶化。 但是,有时条件有利于保存。 有利于化石形成的材料的例子包括火山灰、石灰石和矿化地下水。 科学家没有过去生活过的所有东西的化石,在某些情况下,只发现了某个物种中少数个体的遗骸。 化石记录非常不完整。 芝加哥菲尔德自然历史博物馆馆长罗伯特·马丁估计,已经有6,000多种灵长类动物物种,而仅发现了3%的遗骸。 化石非常罕见,但它们对人类生物进化的信息非常丰富。
理解化石
了解化石的一个重要部分是确定它们的年龄并按时间顺序排列。 为了使用灵长类化石来重建灵长类动物的进化史,人类学家必须首先能够大致估计该特定化石的年龄。 一段时间以来,相对定年方法是化石定年的唯一可用方法。 相对定年法计算出化石与其他化石标本相比的大概年龄. 在过去的半个世纪中,绝对定年法取得了重要进展,包括使灵长类动物进化早期阶段的定年成为可能的技术。 绝对定年法计算出化石在一定年内以年为单位的实际生物学年龄.
相对约会技巧
地层学是最著名和最常用的相对测年方法. 地层学基于这样的观察,即土壤沉积在连续的层或地层中。 最古老的土壤层(以及其中的任何文物或化石)将出现在最近的土壤层(以及其中的任何文物或化石)之下。 除了利用土壤层的位置来确定沉积在这些层中的化石的日期外,生物人类学家有时还会使用在特定土壤层中持续发现的其他物品。 这些物品之所以被称为指示文物,是因为它们有助于表明化石和其他文物的相对年龄。 最好的指示文物是那些地理分布广泛、在地质时间很短的时间内存在和/或来自经历快速进化变化的物种的文物。 不同的指标伪像已被用来确定世界不同地区的相对年龄。 在非洲,大象、猪和马被用来确定不同地质地层的相对日期。 例如,东非奥尔杜瓦伊峡谷的地层学是根据化石猪建立的。 连续地层中的各种猪种类是不同且截然不同的,这使古人类学家能够根据其中的猪种类来区分地层。 一旦确定了某个地区的地层学, 不同地点的两种不同化石的相对年龄可以通过相关的指示人工制品来确定.
如果某个地点受到干扰,地层学将不是确定相对年龄的令人满意的方法。 在这种情况下, 可以使用绝对测年方法来估计在受干扰地点一起发现的化石的年龄.
绝对约会技巧
许多绝对测年方法都是基于放射性同位素的衰变速率. 放射性同位素是一种通过自发发射辐射消散多余能量的化学元素。 这些排放以已知且稳定的速率发生。 一旦确定了放射性同位素的衰变率, 可以在可能的误差范围内估算出含有该同位素的标本的年龄.
C-14
确定化石绝对年龄的最著名方法是碳14或 14 C (发音为 “C-14”) 测年. 所有动植物都含有同位素碳 14 (14 C)。 植物从空气中吸收 14 摄氏度,动物摄取含有同位素的植物。 由于植物仅在活着时吸收 14 摄氏度,而动物只在活着时消耗植物,因此科学家可以根据其细胞中残留的 14 摄氏度来确定动物或植物在多久之前死亡。 已知的碳14的半衰期为5,730年。 这意味着生物体中最初的 14 摄氏度中大约有一半将在其死亡后的5,730年内被消除。 例如,如果一个生物体的原始 14 摄氏度值为100,那么在5,730年后,将只有50个单位的 14 摄氏度存在。
热发光
古人类学家经常使用的另一种绝对测年技术是热发光测年. 热发光测年法要求对化石进行测年或化石所在的沉积物必须暴露在高温事件中,例如火山爆炸。 在这种高温事件中,材料中的所有放射性元素都会被释放。 因此,自高温事件发生以来在人工制品中积聚的放射性元素的数量可用于计算人工制品的年龄。
古新世时代的灵长类动物
古新世时代始于大约 65 年 MYA,大约 54 MYA 结束。 这是新生代最鲜为人知的时代,因为这是代表它的化石最少的时期。 但是,这个时代被认为对灵长类动物的进化很重要,因为它提供了最早灵长类动物的第一个明确记录。 在美国蒙大拿州的一个可追溯到古新世早期的地质矿床中发现了迄今为止已发现的最原始灵长类动物的证据。 这种生物被称为 Purgatorius。 Purgatorius 与已灭绝的活灵长类动物相似,与其他哺乳动物截然不同,因为最后一个下磨牙很长,中上部门牙增大(导致人们可能认为是 “Bugs Bunny 牙齿”)。 当今所有活灵长类动物都具有这两个特征,这表明 Purgatorius 可能是后来的灵长类动物的共同祖先。

始新世时代的灵长类动物
始新世时代始于大约 54 MYA,大约 34 MYA 结束,其标志是 Purgatorius 的消失以及与现代灵长类动物更相似的灵长类动物首次出现,尤其是它们拥有完全由骨头组成的后轨棒。 postorbital bar 是围绕整个眼眶的骨环。 这与其他哺乳动物形成鲜明对比,后者的 postorbital 棒既是骨骼又是软骨。 一些化石标本还具有牙梳和/或美容爪,这些特征仅存在于当今的链霉素灵长类动物中。 其他重要的解剖学特征是脚踝骨,研究人员认为脚踝骨在灵长类动物的进化成功中起着关键作用。 始新世时期灵长类动物的进化是巨大的。 据推测,始新世时期链球菌灵长类动物的数量是当今活灵长类动物的四倍。 始新世矿床中的化石灵长类动物在北美和欧洲很常见,在亚洲和非洲也越来越广为人知。 但是,目前在南美或南极洲没有已知的始新世化石灵长类动物。
渐新世时代的灵长类动物
渐新世时代始于大约 34 MYA,大约 22 MYA 结束,标志着第一批化石猴的出现。 在距离埃及开罗约60英里的法尤姆考古遗址发现了最早的明确的单洛莱茵化石,如今已成为撒哈拉沙漠的一部分。 Fayum 灵长类动物分为两大类:Parapithecoide a 和 Propliopithecoid ea。 根据它们的牙齿,这些灵长类动物被认为分别是最早的新世界和旧世界猴子。 牙齿通常是根据牙科公式描述的,该公式表示下巴每个象限中每种类型的牙齿的数量。 具有 2.1.2.3 牙科配方的生物在其上下颌的每个象限中有两个门牙、一个犬、两个前磨牙和三个磨牙。 根据第三种前磨牙的存在(所有新世界猴子都存在这种特征),Propliopithecus 很可能代表了最早的新世界猴子,尽管它们最初是在非洲进化的。 同样,propliopithecoids 很可能是最早的卡他莱茵灵长类动物,因为它们是第一批在卡他莱茵灵长类动物中发现的牙科配方为 2.1.2.3 的化石猴。
中新世类人猿
中新世时代包含一些最早的猿类的化石证据,例如 Proconsul africanus a fricanus,它于 23 至 14 年生活在非洲。 在非洲发现的最早的中新世猿是 Proconsul。 与现代猿类不同,总领事缺少长而弯曲的数字,这表明它们能够悬挂在树枝上,但更常见的是四肢移动。 Proconsul 也缺少尾巴,这就是为什么它们被视为猿而不是猴子的原因。 像包括人类在内的所有旧世界猴子和猿类一样,它们的牙齿呈现2.1.2.3的图案。 另一只来自中新世的著名猿是西瓦皮特库斯。 Sivapithecus 化石在整个亚洲非常常见,在土耳其发现了特别多的化石。 像现代人类一样,它们表现出非常厚的牙釉质,这表明这些猿类经常吃非常硬的食物。 Sivapithecus 形态学中最有趣的方面是,头骨在鼻孔高、眼窝高等特征上与活猩猩极为相似。
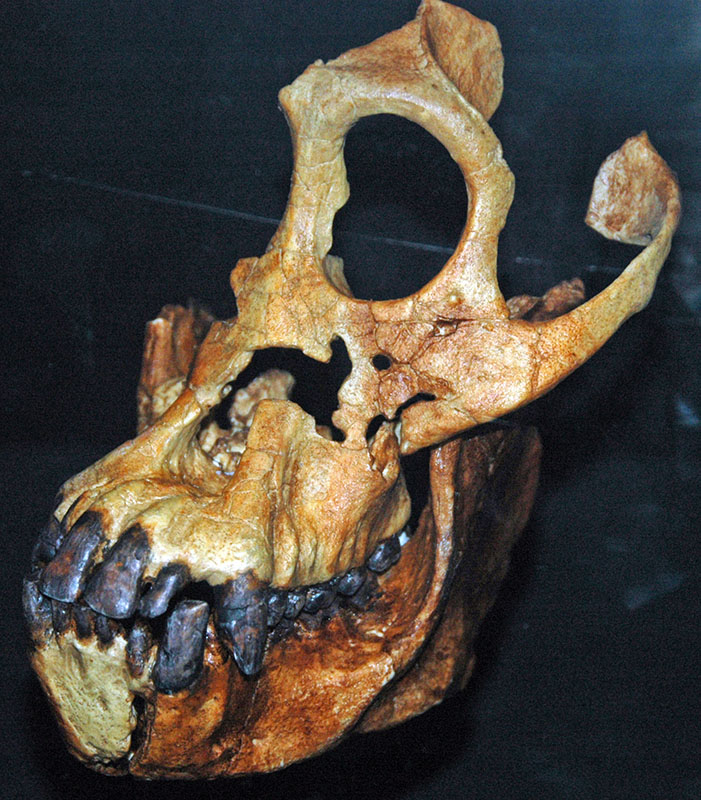
虽然众所周知,猩猩可能是由 Sivapithecus 型猿进化而来的,但现代非洲大猿的祖先没有明确的候选人。 只发现了两种明显而明确地属于现代非洲猿类祖先的化石。 Samburupithecus 是在肯尼亚北部发现的一种中新世晚期的大型猿。 众所周知,它类似于现代非洲类人猿。 它与其他中新世化石的不同之处在于,磨牙从嘴前向后的方向拉长,而不是从脸颊到舌头的方向拉长。 另一种来自中新世晚期(9—10 MYA)的化石有时被认定为现代非洲猿类的祖先,是在希腊发现的 O uranopithecus,其面部形态将其与非洲猿类和人类联系起来。