
3.3: מאפיינים ייחודיים של תאים פרוקריוטים
- Page ID
- 208909

מטרות למידה
- הסבר את המאפיינים המבדילים של תאים פרוקריוטים
- תאר מורפולוגיות תאים נפוצות וסידורים תאיים האופייניים לתאים פרוקריוטים והסבר כיצד תאים שומרים על המורפולוגיה שלהם
- תאר מבנים פנימיים וחיצוניים של תאים פרוקריוטים במונחים של המבנה הפיזי שלהם, המבנה הכימי ותפקודם
- השווה את המאפיינים המבדילים של תאים חיידקיים וארכאיים
תורת התא קובעת שהתא הוא יחידת החיים הבסיסית. עם זאת, התאים משתנים באופן משמעותי בגודל, צורה, מבנה ותפקוד. ברמת הבנייה הפשוטה ביותר, לכל התאים יש כמה מרכיבים בסיסיים. אלה כוללים ציטופלזמה (חומר דמוי ג'ל המורכב ממים וכימיקלים מומסים הדרושים לצמיחה), הכלול בתוך קרום פלזמה (נקרא גם קרום תא או קרום ציטופלזמי); כרומוזום אחד או יותר, המכילים את השרטוטים הגנטיים של התא; וריבוזומים, אברונים המשמשים לייצור חלבונים.
מעבר למרכיבים הבסיסיים הללו, התאים יכולים להשתנות מאוד בין אורגניזמים, ואפילו בתוך אותו אורגניזם רב תאי. שתי הקטגוריות הגדולות ביותר של תאים - תאים פרוקריוטים ותאים אוקריוטיים - מוגדרות על ידי הבדלים גדולים במספר מבני תאים. לתאים פרוקריוטים חסר גרעין המוקף בקרום גרעיני מורכב ובדרך כלל יש להם כרומוזום יחיד ומעגלי הממוקם בנוקלואיד. לתאים אוקריוטיים יש גרעין המוקף בקרום גרעיני מורכב המכיל כרומוזומים מרובים בצורת מוט. 1
כל תאי הצמח ותאי בעלי החיים הם אוקריוטים. חלק מהמיקרואורגניזמים מורכבים מתאים פרוקריוטים, בעוד שאחרים מורכבים מתאים אוקריוטיים. מיקרואורגניזמים פרוקריוטיים מסווגים בתוך התחומים ארכאה וחיידקים, ואילו אורגניזמים אוקריוטיים מסווגים בתחום Eukarya.
המבנים בתוך התא מקבילים לאיברים בתוך גוף האדם, עם מבנים ייחודיים המתאימים לתפקודים ספציפיים. חלק מהמבנים המצויים בתאים פרוקריוטים דומים לאלה המצויים בחלק מהתאים האיקריוטים; אחרים ייחודיים לפרוקריוטים. למרות שיש כמה יוצאים מן הכלל, תאים אוקריוטיים נוטים להיות גדולים יותר מתאים פרוקריוטים. הגודל הגדול יחסית של תאים אוקריוטיים מכתיב את הצורך למדר תהליכים כימיים שונים באזורים שונים של התא, באמצעות אברונים מורכבים הקשורים לממברנה. לעומת זאת, לתאים פרוקריוטים חסרים בדרך כלל אברונים הקשורים לממברנה; עם זאת, לעתים קרובות הם מכילים תכלילים המחלקים את הציטופלזמה שלהם. איור \(\PageIndex{1}\) ממחיש מבנים הקשורים בדרך כלל לתאים פרוקריוטים. מבנים אלה מתוארים ביתר פירוט בסעיף הבא.
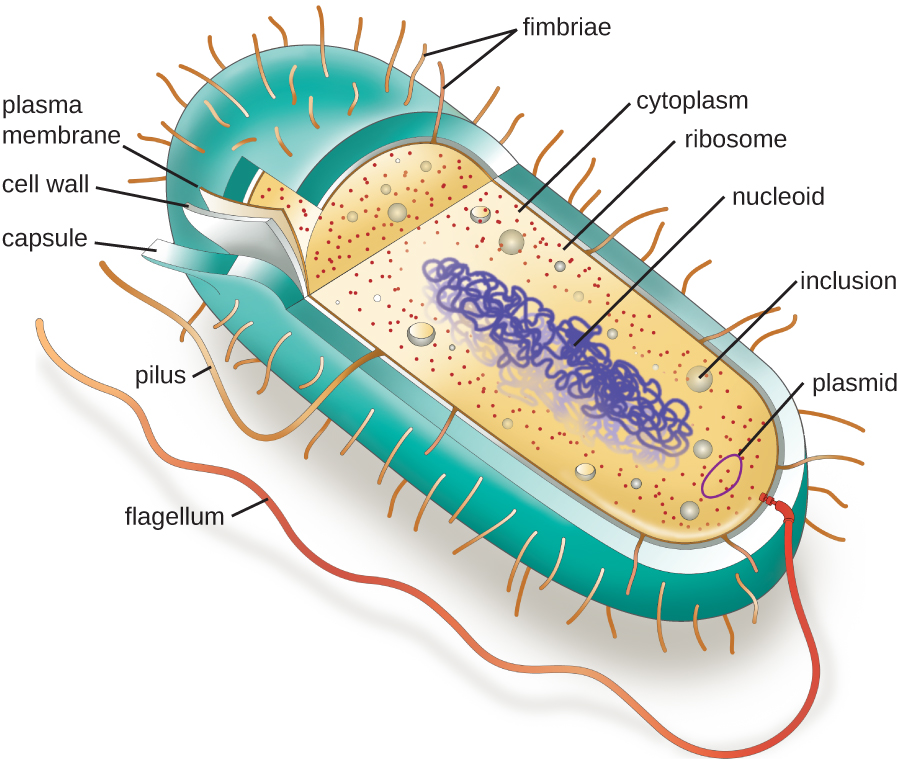
מורפולוגיות וסידורים נפוצים של תאים
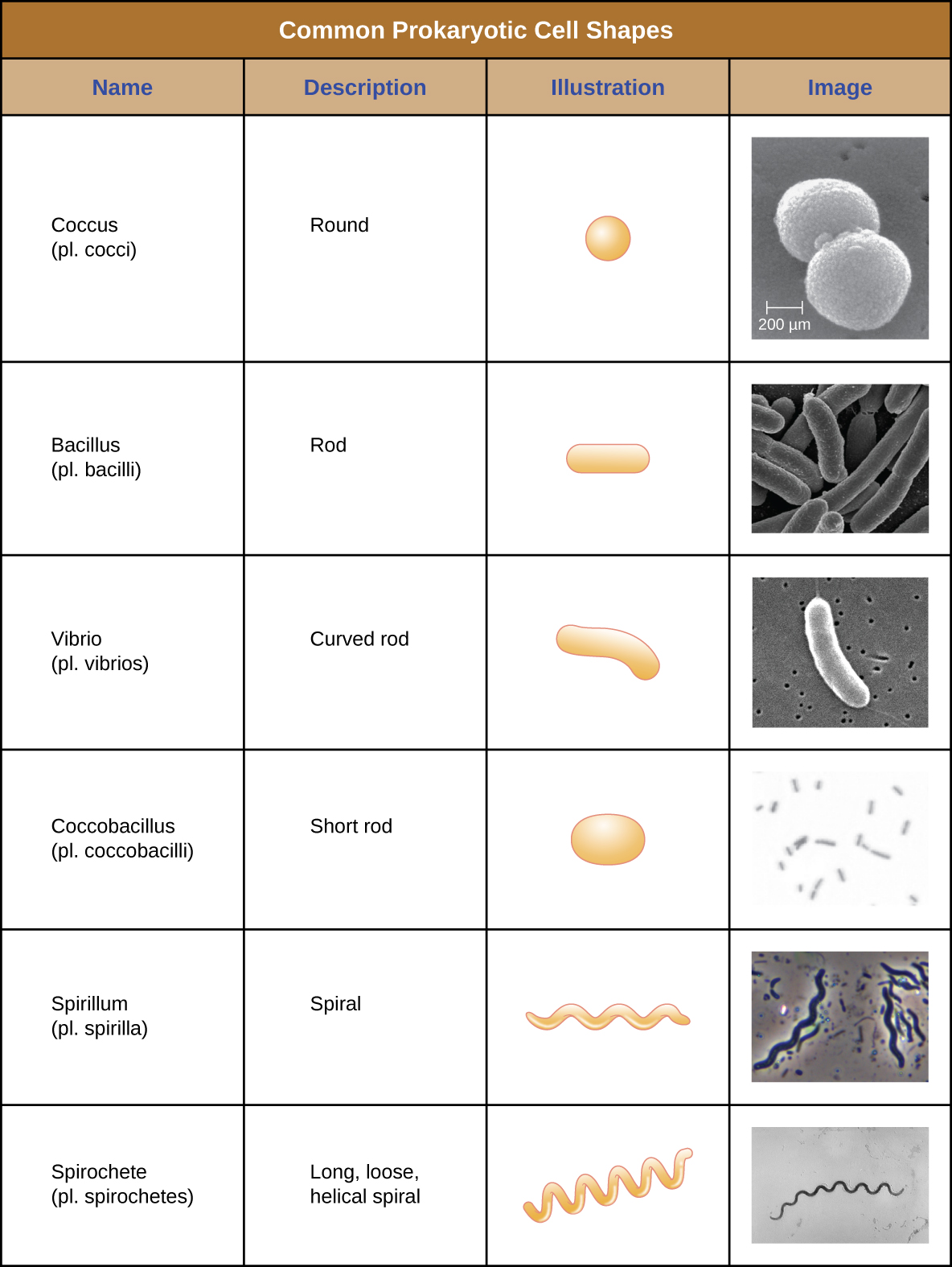
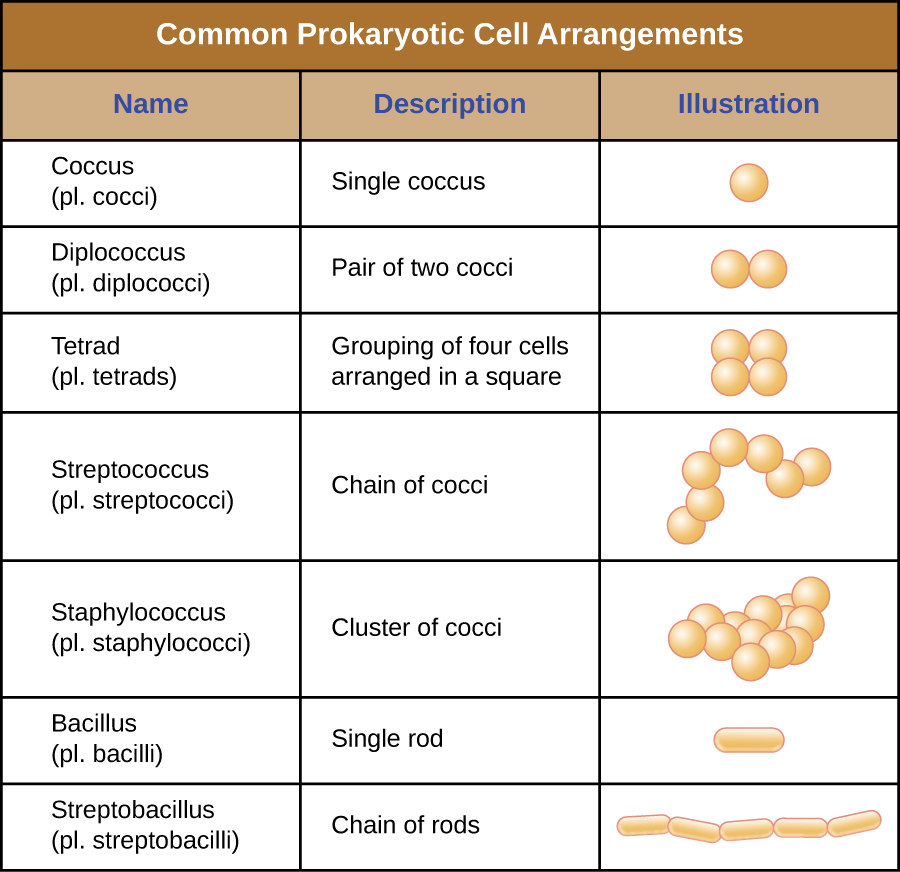
תאים בודדים של אורגניזם פרוקריוטי מסוים דומים בדרך כלל בצורתם, או במורפולוגיה של התא. למרות שזוהו אלפי אורגניזמים פרוקריוטים, רק קומץ מורפולוגיות תאים נראות בדרך כלל במיקרוסקופ. איור \(\PageIndex{2}\) שם וממחיש מורפולוגיות תאים הנפוצות בתאים פרוקריוטים. בנוסף לצורה התאית, תאים פרוקריוטים מאותו מין עשויים להתקבץ יחד בסידורים ייחודיים מסוימים בהתאם למישור חלוקת התא. כמה סידורים נפוצים מוצגים באיור\(\PageIndex{3}\).
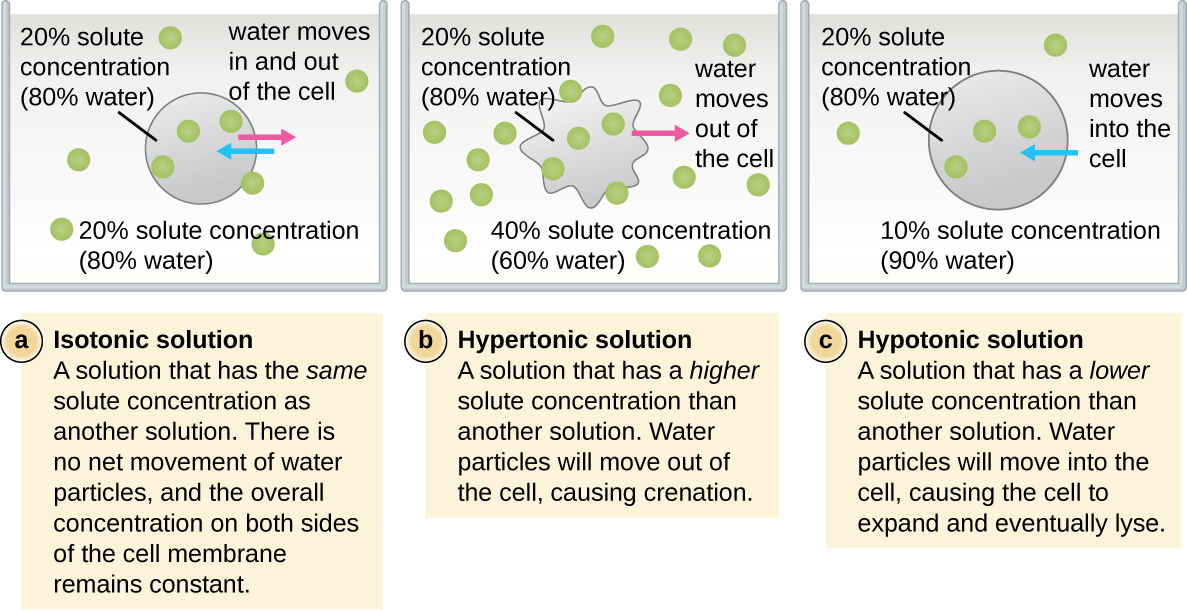
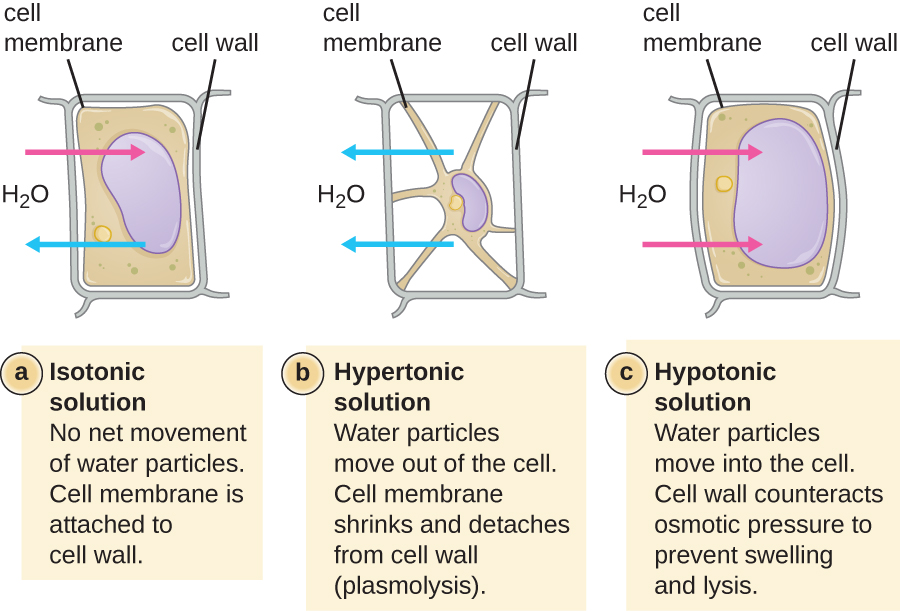
ברוב התאים הפרוקריוטים, המורפולוגיה נשמרת על ידי דופן התא בשילוב עם אלמנטים ציטו-שלד. דופן התא היא מבנה המצוי ברוב הפרוקריוטים ובחלק מהאוקריוטים; הוא עוטף את קרום התא, ומגן על התא מפני שינויים בלחץ האוסמוטי (איור\(\PageIndex{4}\)). לחץ אוסמוטי מתרחש בגלל הבדלים בריכוז המומסים בצדדים מנוגדים של קרום חדיר למחצה. מים מסוגלים לעבור דרך קרום חדיר למחצה, אך מומסים (מולקולות מומסות כמו מלחים, סוכרים ותרכובות אחרות) אינם יכולים. כאשר ריכוז המומסים גדול יותר בצד אחד של הממברנה, מים מתפזרים על פני הממברנה מהצד עם הריכוז הנמוך יותר (יותר מים) לצד עם הריכוז הגבוה יותר (פחות מים) עד שהריכוזים משני הצדדים הופכים שווים. דיפוזיה זו של מים נקראת אוסמוזה, והיא עלולה לגרום ללחץ אוסמוטי קיצוני על התא כאשר סביבתו החיצונית משתנה.
ניתן לתאר את הסביבה החיצונית של התא כמדיום איזוטוני, היפרטוני או היפוטוני. במדיום איזוטוני, ריכוזי המומסים בתוך התא ומחוצה לו שווים בערך, כך שאין תנועה נטו של מים על פני קרום התא. במדיום היפרטוני, ריכוז המומסים מחוץ לתא עולה על זה שבתוך התא, ולכן מים מתפזרים החוצה מהתא ואל המדיום החיצוני. במדיום היפוטוני, ריכוז המומסים בתוך התא עולה על זה שמחוץ לתא, כך שהמים ינועו על ידי אוסמוזה לתוך התא. זה גורם לתא להתנפח ועלול להתפוגג, או להתפוצץ.
המידה שבה תא מסוים מסוגל לעמוד בשינויים בלחץ האוסמוטי נקראת טוניקות. תאים בעלי דופן תא מסוגלים לעמוד טוב יותר בשינויים עדינים בלחץ האוסמוטי ולשמור על צורתם. בסביבות היפרטוניות, תאים חסרי דופן תא יכולים להתייבש, לגרום להתכווצות או להתכווצות של התא; קרום הפלזמה מתכווץ ונראה מסולסל או מחורץ (איור). \(\PageIndex{4}\) לעומת זאת, תאים בעלי דופן תא עוברים פלסמוליזה ולא קרנציה. בפלסמוליזה, קרום הפלזמה מתכווץ ומתנתק מדופן התא, ויש ירידה בנפח הפנימי, אך דופן התא נשארת שלמה, ובכך מאפשרת לתא לשמור על צורה ושלמות מסוימת למשך פרק זמן (איור). \(\PageIndex{5}\) באופן דומה, תאים חסרי דופן תא מועדים יותר לתמוגה בסביבות היפוטוניות. הנוכחות של דופן תא מאפשרת לתא לשמור על צורתו ושלמותו למשך זמן רב יותר לפני הליסינג (איור\(\PageIndex{5}\)).
תרגיל \(\PageIndex{1}\)
- הסבר את ההבדל בין מורפולוגיה של התא לסידור.
- אילו יתרונות מספקים קירות התא לתאים פרוקריוטים?
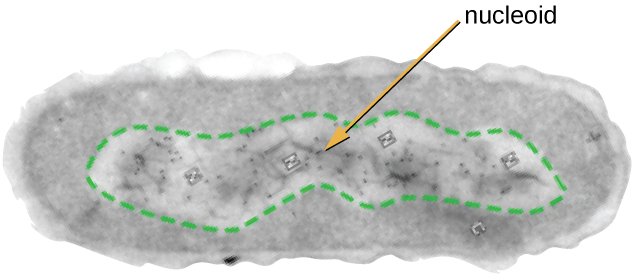
הנוקלאואיד
לכל חיי התא יש גנום DNA המאורגן לכרומוזום אחד או יותר. כרומוזומים פרוקריוטיים הם בדרך כלל מעגליים, הפלואידים (לא מזווגים) ואינם קשורים לממברנה גרעינית מורכבת. DNA פרוקריוטי וחלבונים הקשורים ל-DNA מרוכזים באזור הנוקלאואיד של התא (איור). \(\PageIndex{6}\) באופן כללי, DNA פרוקריוטי מקיים אינטראקציה עם חלבונים הקשורים לנוקלאואידים (NAP) המסייעים בארגון ובאריזה של הכרומוזום. בחיידקים, NAPs מתפקדים בדומה להיסטונים, שהם החלבונים המארגנים DNA המצויים בתאים אוקריוטיים. בארכאה, הנוקלאואיד מאורגן על ידי NAPs או חלבונים המארגנים DNA דמוי היסטון.
פלסמידים
תאים פרוקריוטים עשויים להכיל גם DNA אקסטרכרומוזומלי, או DNA שאינו חלק מהכרומוזום. DNA חוץ-כרומוזומלי זה נמצא בפלסמידים, שהם מולקולות DNA קטנות, מעגליות, דו-גדיליות. לתאים שיש להם פלסמידים יש לעתים קרובות מאות מהם בתוך תא בודד. פלסמידים נמצאים בדרך כלל יותר בחיידקים; עם זאת, פלסמידים נמצאו באורגניזמים ארכאים ואוקריוטיים. פלסמידים נושאים לעתים קרובות גנים המעניקים תכונות מועילות כגון עמידות לאנטיביוטיקה; לפיכך, הם חשובים להישרדות האורגניזם. נדון בפלסמידים ביתר פירוט במנגנונים של גנטיקה מיקרוביאלית.
ריבוזומים
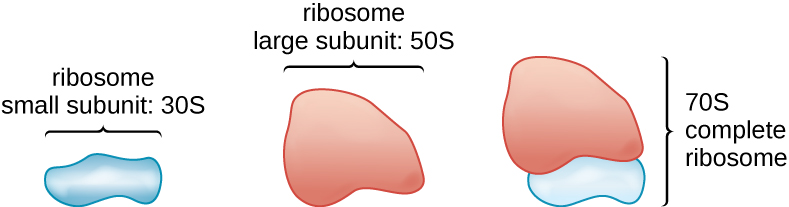
כל החיים התאיים מסנתזים חלבונים, ולאורגניזמים בכל שלושת תחומי החיים יש ריבוזומים, מבנים האחראים לסינתזת חלבון. עם זאת, הריבוזומים בכל אחד משלושת התחומים שונים מבחינה מבנית. הריבוזומים, עצמם, בנויים מחלבונים, יחד עם RNA ריבוזומלי (rRNA). ריבוזומים פרוקריוטיים נמצאים בציטופלזמה. הם נקראים ריבוזומים 70S מכיוון שיש להם גודל של 70S (איור\(\PageIndex{7}\)), ואילו לריבוזומים ציטופלסמיים אוקריוטיים יש גודל של 80S. (ה- S מייצג יחידת סוודברג, מדד שקיעה באולטרה-צנטריפוגה, המבוסס על גודל, צורה ואיכויות פני השטח של המבנה המנותח). למרות שהם באותו גודל, לריבוזומים חיידקיים וארכאיים יש חלבונים ומולקולות rRNA שונות, והגרסאות הארכאיות דומות יותר למקביליהם האוקריוטיים מאשר לאלה המצויים בחיידקים.
תכלילים
כאורגניזמים חד-תאיים החיים בסביבות לא יציבות, לחלק מהתאים הפרוקריוטים יש את היכולת לאחסן עודפי חומרים מזינים בתוך מבנים ציטופלזמיים הנקראים תכלילים. אחסון חומרים מזינים בצורה פולימרית הוא יתרון מכיוון שהוא מפחית את הצטברות הלחץ האוסמוטי המתרחש כאשר תא צובר מומסים. סוגים שונים של תכלילים אוגרים גליקוגן ועמילנים, המכילים פחמן שתאים יכולים לגשת אליו לאנרגיה. גרגירי וולוטין, הנקראים גם גרגירים מטכרומטיים בגלל מאפייני הצביעה שלהם, הם תכלילים המאחסנים פוספט אנאורגני מפולימר שניתן להשתמש בהם בחילוף חומרים ולסייע ביצירת ביופילמים. חיידקים הידועים כמכילים גרגירי וולטין כוללים את הארכיאה מתנוסארצ'ינה, החיידק Corynebacterium diphtheriae והאצה האוקריוטית החד-תאית כלמידומונס. גרגרי גופרית, סוג אחר של הכללה, נמצאים בחיידקי גופרית מהסוג Thiobacillus; גרגירים אלה אוגרים גופרית יסודית, בה משתמשים החיידקים למטבוליזם.
מדי פעם, סוגים מסוימים של תכלילים מוקפים בשכבה חד-שכבתית פוספוליפידית המוטמעת בחלבון. Polyhydroxybutyrate (PHB), אשר יכול להיות מיוצר על ידי מינים של Bacillus ו Pseudomonas, הוא דוגמה להכללה המציגה סוג זה של מבנה חד-שכבתי. מבחינה תעשייתית, PHB שימש גם כמקור לפולימרים מתכלים לביו-פלסטיקה. מספר סוגים שונים של תכלילים מוצגים באיור\(\PageIndex{8}\).

לחלק מהתאים הפרוקריוטים יש סוגים אחרים של תכלילים המשרתים מטרות אחרות מלבד אחסון חומרים מזינים. לדוגמה, חלק מהתאים הפרוקריוטים מייצרים ואקום גז, הצטברות של שלפוחיות גז קטנות ומרופדות חלבון. שואבי גז אלה מאפשרים לתאים הפרוקריוטים המסנתזים אותם לשנות את הציפה שלהם כך שיוכלו להתאים את מיקומם בעמודת המים. חיידקים מגנטוטקטיים, כמו Magnetospirillum magnetotacticum, מכילים מגנטוזומים, שהם תכלילים של תחמוצת ברזל מגנטית או ברזל גופרתי המוקפים בשכבת שומנים. אלה מאפשרים לתאים להתיישר לאורך שדה מגנטי, ומסייעים לתנועתם (איור\(\PageIndex{8}\)). ציאנובקטריה כגון Anabaena cylindrica וחיידקים כגון Halothiobacillus neapolitanus מייצרים תכלילים קרבוקסיזומים. קרבוקסיזומים מורכבים מקליפות חיצוניות של אלפי יחידות משנה של חלבון. החלק הפנימי שלהם מלא בריבולוז-1,5-ביספוספט קרבוקסילאז/חמצן (RuBisCO) ואנהידראז פחמני. שתי התרכובות הללו משמשות למטבוליזם של פחמן. לחלק מהתאים הפרוקריוטים יש גם קרבוקסיזומים הסוגרים אנזימים הקשורים לתפקוד במיקום אחד. מבנים אלה נחשבים לפרוטו-אברונים מכיוון שהם מחלקים תרכובות חשובות או תגובות כימיות, בדומה לאברונים אוקריוטיים רבים.
אנדוספורים
תאים חיידקיים נצפים בדרך כלל כתאים וגטטיביים, אך לחלק מהסוגים של חיידקים יש את היכולת ליצור אנדוספורים, מבנים המגנים בעצם על הגנום החיידקי במצב רדומה כאשר תנאי הסביבה אינם נוחים. אנדוספורים (אין להתבלבל עם נבגי הרבייה הנוצרים על ידי פטריות) מאפשרים לתאי חיידקים מסוימים לשרוד תקופות ארוכות ללא מזון או מים, כמו גם חשיפה לכימיקלים, טמפרטורות קיצוניות ואפילו קרינה. הטבלה \(\PageIndex{1}\) משווה את המאפיינים של תאים וגטטיביים ואנדוספורים.
| תאים צמחיים | אנדוספורים |
|---|---|
| רגיש לטמפרטורות קיצוניות וקרינה | עמיד בפני טמפרטורות קיצוניות וקרינה |
| גראם חיובי | אין לספוג כתם גראם, רק כתמי אנדוספור מיוחדים (ראה צביעת דגימות מיקרוסקופיות) |
| תכולת מים רגילה ופעילות אנזימטית | מיובש; ללא פעילות מטבולית |
| מסוגל לצמיחה פעילה ומטבוליזם | רָדוֹם; ללא צמיחה או פעילות מטבולית |
התהליך שבו תאים וגטטיביים הופכים לאנדוספורים נקרא ספורולציה, והוא מתחיל בדרך כלל כאשר חומרים מזינים מתרוקנים או שתנאי הסביבה הופכים לבלתי נוחים אחרת (איור). \(\PageIndex{9}\) התהליך מתחיל עם היווצרות של מחיצות בתא חיידקי הצומח. המחיצה מחלקת את התא בצורה לא סימטרית, ומפרידה בין תא DNA לתא האם. ה-forespore, שיהווה את ליבת האנדוספור, הוא בעצם עותק של הכרומוזומים של התא, ומופרד מתא האם על ידי קרום שני. קליפת המוח נוצרת בהדרגה סביב פורפורפור על ידי הנחת שכבות של סידן וחומצה דיפיקולינית בין הממברנות. לאחר מכן נוצר מעטפת נבג חלבון סביב קליפת המוח בעוד ה- DNA של תא האם מתפרק. התבגרות נוספת של האנדוספור מתרחשת עם היווצרות אקסוספוריום חיצוני ביותר. האנדוספור משתחרר עם התפוררות תא האם, ומשלים ספורולציה.
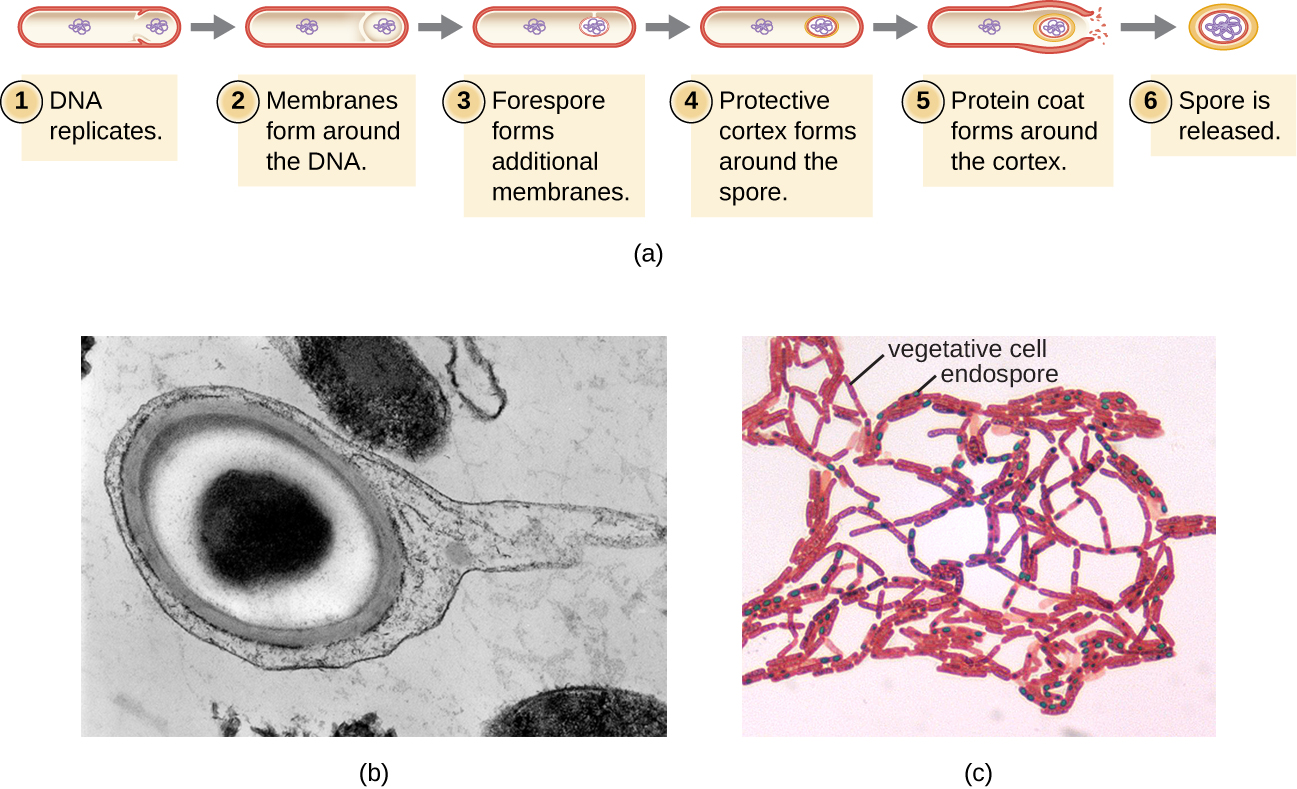
הוכח כי אנדוספורים ממינים מסוימים נמשכים במצב רדומה לפרקי זמן ממושכים, עד אלפי שנים. 2 עם זאת, כאשר תנאי החיים משתפרים, האנדוספורים עוברים נביטה, ונכנסים שוב למצב וגטטיבי. לאחר הנביטה, התא הופך שוב לפעיל מטבולית ומסוגל לבצע את כל תפקידיו הרגילים, כולל צמיחה וחלוקת תאים.
לא לכל החיידקים יש את היכולת ליצור אנדוספורים; עם זאת, ישנם מספר חיידקים חיוביים לגרם חיוביים היוצרים אנדוספור בעלי משמעות קלינית מהסוגים Bacillus ו- Clostridium. אלה כוללים B. anthracis, הסוכן הסיבתי של אנתרקס, המייצר אנדוספורים המסוגלים לשרוד במשך עשורים רבים 3; C. טטני (גורם לטטנוס); C. difficile (גורם לקוליטיס פסאודוממברנית); C. perfringens (גורם לגנגרן בגז); ו- C. botulinum (גורם לבוטוליזם). קשה במיוחד להילחם בפתוגנים כמו אלה מכיוון שקשה להרוג את האנדוספורים שלהם. שיטות עיקור מיוחדות לחיידקים יוצרי אנדוספור נדונות בשליטה על צמיחה מיקרוביאלית.
תרגיל \(\PageIndex{2}\)
- מהי הכללה?
- מה תפקידו של אנדוספור?
ממברנה פלזמה
מבנים הסוגרים את הציטופלזמה והמבנים הפנימיים של התא ידועים ביחד בשם מעטפת התא. בתאים פרוקריוטים, המבנים של מעטפת התא משתנים בהתאם לסוג התא והאורגניזם. לרוב התאים הפרוקריוטים (אך לא לכולם) יש דופן תא, אך ההרכב של דופן התא הזה משתנה. לכל התאים (פרוקריוטים ואוקריוטיים) יש קרום פלזמה (הנקרא גם קרום ציטופלזמי או קרום תא) המציג חדירות סלקטיבית, המאפשר לחלק מהמולקולות להיכנס לתא או לצאת ממנו תוך הגבלת מעבר של אחרים.
מבנה קרום הפלזמה מתואר לעתים קרובות במונחים של מודל הפסיפס הנוזלי, המתייחס ליכולתם של רכיבי הממברנה לנוע בצורה נוזלית בתוך מישור הממברנה, כמו גם ההרכב דמוי הפסיפס של הרכיבים, הכוללים מגוון מגוון של רכיבי שומנים וחלבונים (איור). \(\PageIndex{10}\) מבנה קרום הפלזמה של רוב סוגי התאים החיידקיים והאוקריוטיים הוא דו שכבתי המורכב בעיקר מפוספוליפידים הנוצרים עם קישורי אסטר וחלבונים. לפוספוליפידים וחלבונים אלה יש את היכולת לנוע לרוחב בתוך מישור הממברנות וכן בין שתי שכבות הפוספוליפידים.

ממברנות ארכאיות שונות מהותית מממברנות חיידקיות ואיקריוטיות בכמה דרכים משמעותיות. ראשית, פוספוליפידים ממברנה ארכאית נוצרים עם קישורי אתר, בניגוד לקישורי האסטר המצויים בממברנות תאים חיידקיות או אוקריוטיות. שנית, לפוספוליפידים ארכאיים יש שרשראות מסועפות, ואילו אלה של תאים חיידקיים ואיקריוטים קשורים ישר. לבסוף, למרות שכמה ממברנות ארכאיות יכולות להיווצר מדו-שכבות כמו אלה המצויות בחיידקים ובאוקריוטים, ממברנות פלזמה ארכאיות אחרות הן חד-שכבות שומנים.
חלבונים על פני התא חשובים למגוון פונקציות, כולל תקשורת בין תא לתא, וחישת תנאים סביבתיים וגורמי ארסיות פתוגניים. לחלבוני ממברנה ופוספוליפידים עשויים להיות פחמימות (סוכרים) הקשורים אליהם ונקראים גליקופרוטאינים או גליקוליפידים, בהתאמה. מתחמי גליקופרוטאין וגליקוליפידים אלה משתרעים מפני השטח של התא, ומאפשרים לתא לקיים אינטראקציה עם הסביבה החיצונית (איור\(\PageIndex{10}\)). גליקופרוטאינים וגליקוליפידים בקרום הפלזמה יכולים להשתנות במידה ניכרת בהרכב הכימי בין ארכאים, חיידקים ואיקריוטים, מה שמאפשר למדענים להשתמש בהם כדי לאפיין מינים ייחודיים.
ממברנות פלזמה מסוגי תאים שונים מכילות גם פוספוליפידים ייחודיים, המכילים חומצות שומן. כפי שמתואר בשימוש בביוכימיה לזיהוי מיקרואורגניזמים, ניתן להשתמש בפרופילי ניתוח חומצות שומן (PLFA) שמקורם בפוספוליפידים לזיהוי סוגים ייחודיים של תאים המבוססים על הבדלים בחומצות שומן. לארכיאה, לחיידקים ולאוקריוטים יש פרופיל PFLA ייחודי.
מנגנוני הובלת ממברנה
אחד התפקידים החשובים ביותר של קרום הפלזמה הוא לשלוט בהובלת מולקולות לתא ומחוצה לו. יש לשמור על תנאים פנימיים בטווח מסוים למרות כל שינוי בסביבה החיצונית. הובלת חומרים על פני קרום הפלזמה מאפשרת לתאים לעשות זאת.
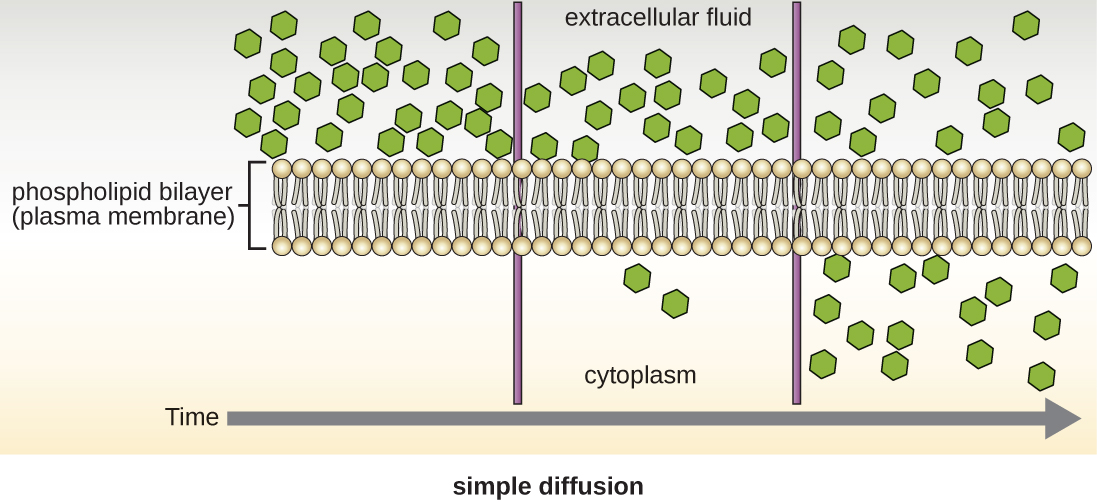
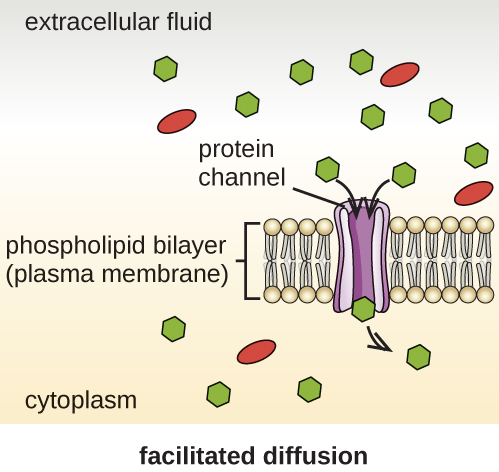
תאים משתמשים באופני הובלה שונים על פני קרום הפלזמה. לדוגמה, מולקולות הנעות מריכוז גבוה יותר לריכוז נמוך יותר עם שיפוע הריכוז מועברות על ידי דיפוזיה פשוטה, המכונה גם הובלה פסיבית (איור\(\PageIndex{11}\)). כמה מולקולות קטנות, כמו פחמן דו חמצני, עשויות לחצות את דו שכבת הממברנה ישירות על ידי דיפוזיה פשוטה. עם זאת, מולקולות טעונות, כמו גם מולקולות גדולות, זקוקות לעזרת נשאים או תעלות בקרום. מבנים אלה מעבירים מולקולות על פני הממברנה, תהליך המכונה דיפוזיה מקלה (איור\(\PageIndex{12}\)).
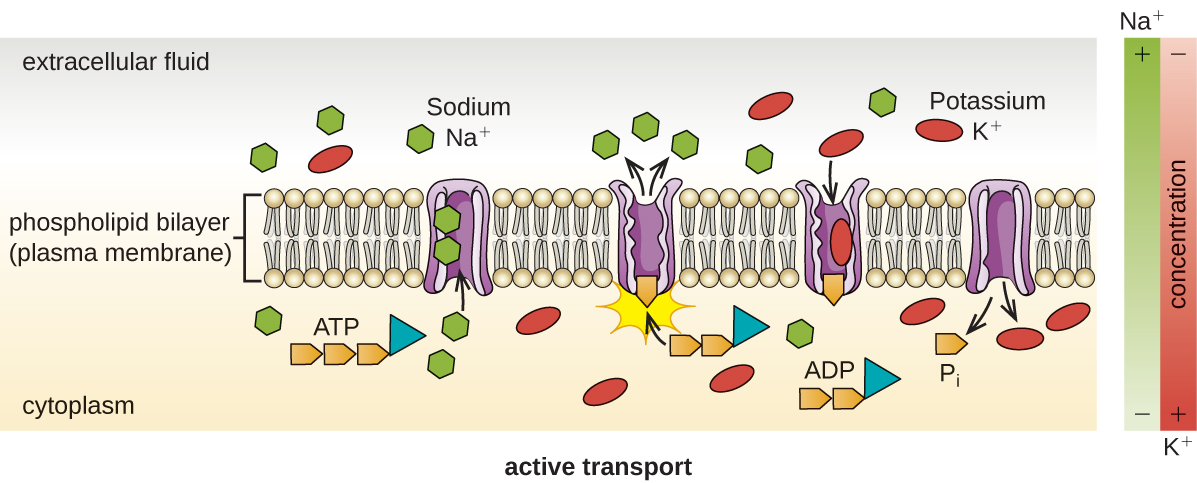
הובלה פעילה מתרחשת כאשר תאים מעבירים מולקולות על פני הממברנה שלהם כנגד שיפועי ריכוז (איור\(\PageIndex{13}\)). ההבדל העיקרי בין הובלה פסיבית לאקטיבית הוא שהובלה פעילה דורשת אדנוסין טריפוספט (ATP) או צורות אחרות של אנרגיה כדי להזיז מולקולות "במעלה ההר". לכן, מבני תחבורה פעילים נקראים לעתים קרובות "משאבות".
טרנסלוקציה קבוצתית מעבירה גם חומרים לתאי חיידקים. במקרה זה, כאשר מולקולה עוברת לתא כנגד שיפוע הריכוז שלה, היא משתנה כימית כך שהיא אינה דורשת הובלה כנגד שיפוע ריכוז שלילי. דוגמה נפוצה לכך היא מערכת הפוספוטרנספראז החיידקית, סדרה של נשאים המזרחים (כלומר, מוסיפים יוני פוספט ל) גלוקוז או סוכרים אחרים עם הכניסה לתאים. מכיוון שנדרש זרחון של סוכרים בשלבים המוקדמים של חילוף החומרים בסוכר, מערכת הפוספוטרנספרז נחשבת למערכת ניטרלית אנרגטית.
מבני ממברנה פוטוסינתטיים
לחלק מהתאים הפרוקריוטים, כלומר ציאנובקטריה וחיידקים פוטוסינתטיים, יש מבני קרום המאפשרים להם לבצע פוטוסינתזה. מבנים אלה מורכבים מקיפול של קרום הפלזמה המקיף פיגמנטים פוטוסינתטיים כגון כלורופילים ירוקים ובקטריוכלורופילים. בציאנובקטריה, מבני הממברנה הללו נקראים תילקואידים; בחיידקים פוטוסינתטיים הם נקראים כרומטופורים, למלות או כלורוזומים.
קיר תא
תפקידו העיקרי של דופן התא הוא להגן על התא מפני תנאים קשים בסביבה החיצונית. כאשר הם קיימים, ישנם קווי דמיון והבדלים בולטים בין דפנות התא של ארכאה, חיידקים ואיקריוטים.
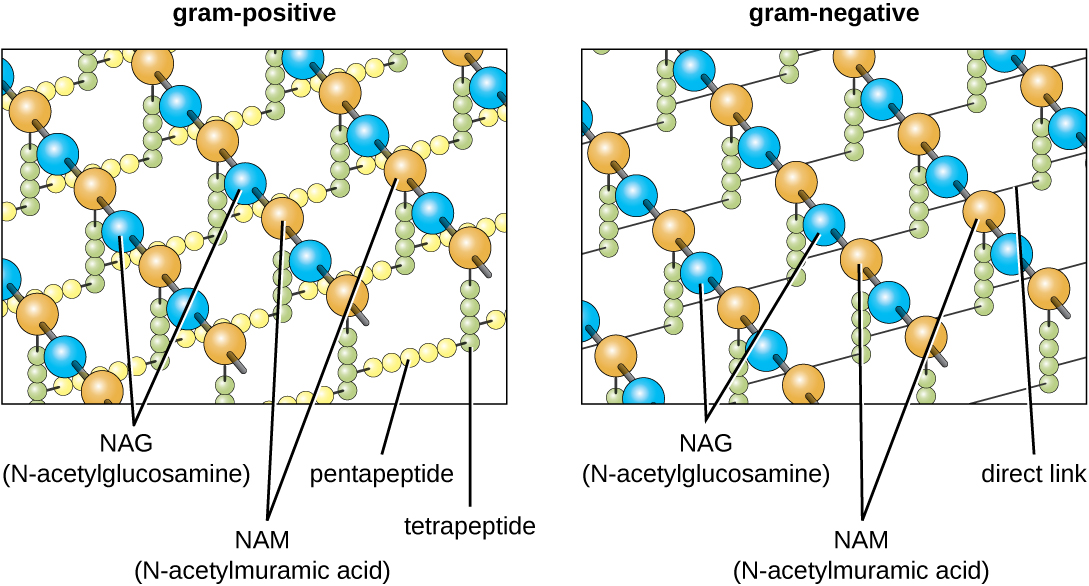
המרכיב העיקרי בדפנות התא החיידקי נקרא פפטידוגליקן (או מורין); הוא נמצא רק בחיידקים. מבחינה מבנית, פפטידוגליקן דומה לשכבה של רשת או בד (איור). \(\PageIndex{14}\) כל שכבה מורכבת משרשראות ארוכות של מולקולות מתחלפות של N-acetylglucosamine (NAG) וחומצה N-acetylmuramic (NAM). למבנה השרשראות הארוכות יש חוזק מתיחה דו-ממדי משמעותי עקב היווצרות גשרי פפטיד המחברים NAG ו-NAM בתוך כל שכבת פפטידוגליקן. בחיידקים גרם שליליים, שרשראות טטרפפטיד המשתרעות מכל יחידת NAM מקושרות ישירות, בעוד שבחיידקים גרם חיוביים, שרשראות טטרפפטיד אלו מקושרות על ידי גשרים צולבים מחומשים. יחידות משנה של פפטידוגליקן מיוצרות בתוך תא החיידק ולאחר מכן מיוצאות ומורכבות בשכבות, ומעניקות לתא את צורתו.
מכיוון שפפטידוגליקן ייחודי לחיידקים, תרופות אנטיביוטיות רבות נועדו להפריע לסינתזה של פפטידוגליקן, להחליש את דופן התא ולהפוך את תאי החיידקים לרגישים יותר להשפעות הלחץ האוסמוטי (ראה מנגנונים של תרופות אנטיבקטריאליות). בנוסף, תאים מסוימים במערכת החיסון האנושית מסוגלים "לזהות" פתוגנים חיידקיים על ידי גילוי פפטידוגליקן על פני תא חיידקי; תאים אלה בולעים ומשמידים את התא החיידקי, באמצעות אנזימים כמו ליזוזים, המפרקים ומעכלים את הפפטידוגליקן בדפנות התא שלהם (ראה זיהוי פתוגן ופגוציטוזיס).
פרוטוקול צביעת גראם (ראה צביעת דגימות מיקרוסקופיות) משמש להבדיל בין שני סוגים נפוצים של מבני דופן התא (איור\(\PageIndex{15}\)). לתאים גראם חיוביים יש דופן תא המורכבת משכבות רבות של פפטידוגליקן בעובי של 30-100 ננומטר. שכבות פפטידוגליקן אלו משובצות בדרך כלל בחומצות טיכואיות (TaS), שרשראות פחמימות המשתרעות דרך ומעבר לשכבת הפפטידוגליקן. 4 TA נחשב לייצב פפטידוגליקן על ידי הגברת קשיחותו. TA ממלא גם תפקיד ביכולתם של חיידקים גרם חיוביים פתוגניים כגון סטרפטוקוקוס להיקשר לחלבונים מסוימים על פני תאי המארח, ומשפר את יכולתם לגרום לזיהום. בנוסף לפפטידוגליקן ו- TaS, לחיידקים ממשפחת Mycobacteriaceae יש שכבה חיצונית של חומצות מיקוליות שעווה בדופן התא שלהם; כמתואר בצביעת דגימות מיקרוסקופיות, חיידקים אלה מכונים מהירים בחומצה, שכן יש להשתמש בכתמים מהירים בחומצה כדי לחדור לשכבת החומצה המיקולית למטרות מיקרוסקופיה (איור). \(\PageIndex{16}\)

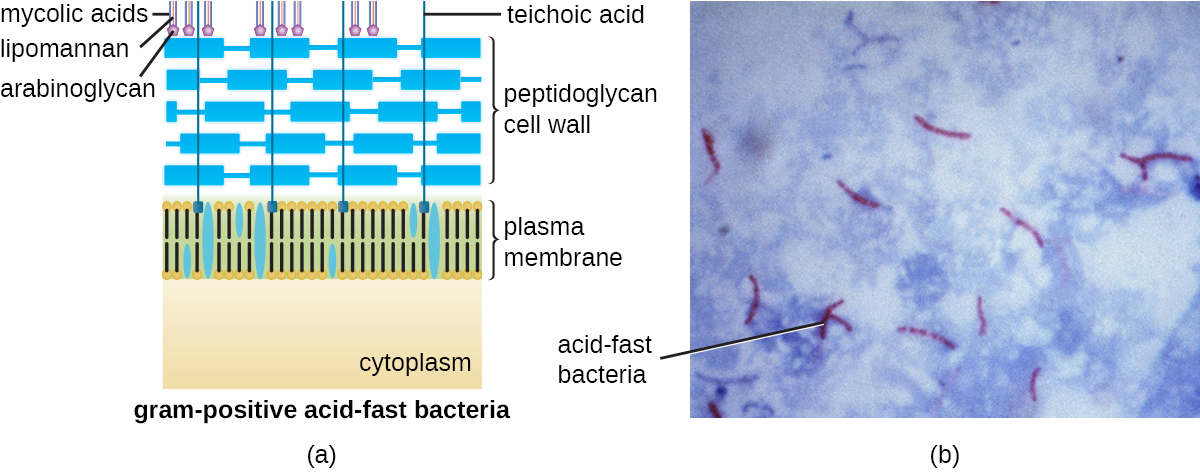
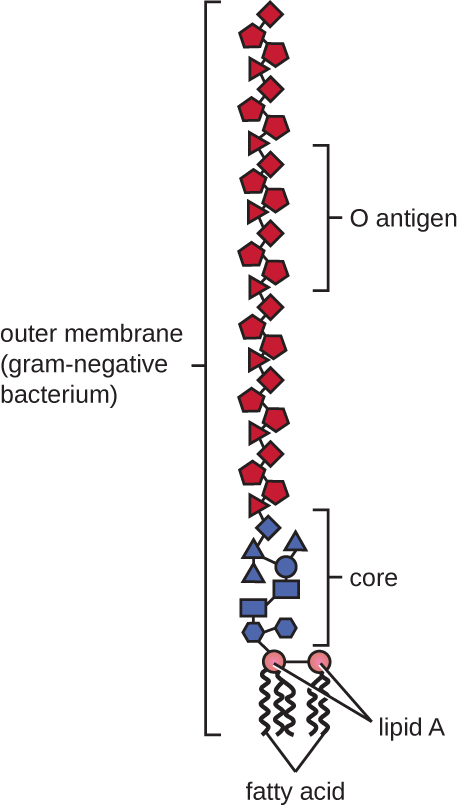
לתאים גראם שליליים יש שכבה דקה בהרבה של פפטידוגליקן (לא יותר מ-4 ננומטר בעובי 6) מאשר תאים חיוביים לגרם, והמבנה הכללי של מעטפת התא שלהם מורכב יותר. בתאים גרם שליליים, מטריצה דמוית ג'ל תופסת את החלל הפריפלזמי בין דופן התא לקרום הפלזמה, ויש דו שכבה שומנית שנייה הנקראת הממברנה החיצונית, שהיא חיצונית לשכבת הפפטידוגליקן (איור). \(\PageIndex{15}\) קרום חיצוני זה מחובר לפפטידוגליקן על ידי ליפופרוטאין מורין. העלון החיצוני של הממברנה החיצונית מכיל את המולקולה ליפופוליסכריד (LPS), המתפקדת כאנדוטוקסין בזיהומים הכוללים חיידקים גרם שליליים, התורמים לתסמינים כגון חום, דימום והלם ספיגה. כל מולקולת LPS מורכבת מליפיד A, פוליסכריד ליבה ושרשרת צד O המורכבת ממולקולות דמויות סוכר המרכיבות את הפנים החיצוניות של ה- LPS (איור). \(\PageIndex{17}\) הרכב שרשרת הצד O משתנה בין מינים שונים לזני חיידקים. ניתן לזהות חלקים משרשרת הצד O הנקראים אנטיגנים באמצעות בדיקות סרולוגיות או אימונולוגיות לזיהוי זנים פתוגניים ספציפיים כמו Escherichia coli O157:H7, זן קטלני של חיידקים הגורם לשלשול עקוב מדם ואי ספיקת כליות.
מבנה דופן התא הארכאי שונה מזה של חיידקים בכמה דרכים משמעותיות. ראשית, קירות תאים ארכאיים אינם מכילים פפטידוגליקן; במקום זאת, הם מכילים פולימר דומה הנקרא pseudopeptidoglycan (pseudomurein) שבו NAM מוחלף ביחידת משנה אחרת. לארכאות אחרות עשויה להיות שכבה של גליקופרוטאינים או פוליסכרידים המשמשת כדופן התא במקום פסאודופפטידוגליקן. אחרון, כמו במקרה של כמה מיני חיידקים, ישנם כמה ארכאים שנראים חסרי דפנות תאים לחלוטין.
גליקוקליסים ושכבות S
למרות שלרוב התאים הפרוקריוטים יש דפנות תאים, לחלקם עשויים להיות מבני מעטפת תאים נוספים מחוץ לדופן התא, כגון גליקוקליסים ושכבות S. גליקוקליקס הוא מעיל סוכר, שיש בו שני סוגים חשובים: כמוסות ושכבות רפש. קפסולה היא שכבה מאורגנת הממוקמת מחוץ לדופן התא ובדרך כלל מורכבת מפוליסכרידים או חלבונים (איור\(\PageIndex{18}\)). שכבת רפש היא שכבה פחות מאורגנת היטב המחוברת רק באופן רופף לדופן התא וניתן לשטוף אותה ביתר קלות. שכבות רפש עשויות להיות מורכבות מפוליסכרידים, גליקופרוטאינים או גליקוליפידים.
Glycocalyces מאפשר לתאים להיצמד למשטחים, ומסייע ביצירת ביופילמים (מושבות של חיידקים הנוצרים בשכבות על משטחים). בטבע, רוב החיידקים חיים בקהילות מעורבות בתוך ביופילמים, בין היתר מכיוון שהביופילם מעניק להם רמה מסוימת של הגנה. ביופילמים בדרך כלל מחזיקים מים כמו ספוג, ומונעים התייבשות. הם גם מגנים על התאים מפני טריפה ומעכבים את פעולתם של אנטיביוטיקה וחומרי חיטוי. כל המאפיינים הללו מועילים לחיידקים החיים בביופילם, אך הם מציבים אתגרים במסגרת קלינית, שבה המטרה היא לעתים קרובות לחסל חיידקים.

היכולת לייצר כמוסה יכולה לתרום לפתוגניות של החיידק (היכולת לגרום למחלות) מכיוון שהקפסולה יכולה להקשות על תאים פגוציטים (כגון תאי דם לבנים) לבלוע ולהרוג את המיקרואורגניזם. Streptococcus pneumoniae, למשל, מייצר כמוסה הידועה כמסייעת בפתוגניות של חיידק זה. כפי שהוסבר בצביעת דגימות מיקרוסקופיות, כמוסות קשות להכתים למיקרוסקופיה; בדרך כלל משתמשים בטכניקות צביעה שליליות.
שכבת S היא סוג אחר של מבנה מעטפת התא; הוא מורכב מתערובת של חלבונים מבניים וגליקופרוטאינים. בחיידקים, שכבות S נמצאות מחוץ לדופן התא, אך בחלק מהארכאות, שכבת ה- S משמשת כדופן התא. הפונקציה המדויקת של שכבות S אינה מובנת לחלוטין, וקשה לחקור אותן; אך עדויות זמינות מצביעות על כך שהם עשויים למלא מגוון פונקציות בתאים פרוקריוטים שונים, כגון עזרה לתא לעמוד בלחץ אוסמוטי, ועבור פתוגנים מסוימים, אינטראקציה עם מערכת החיסון המארחת.
מיקוד קליני: חלק 3
לאחר אבחון ברברה עם דלקת ריאות, הרשות הפלסטינית כותבת לה מרשם לאמוקסיצילין, סוג נפוץ של נגזרת פניצילין. יותר משבוע לאחר מכן, למרות שלקחה את הקורס המלא לפי ההוראות, ברברה עדיין מרגישה חלשה ואינה מתאוששת לחלוטין, אם כי היא עדיין מסוגלת לעבור את פעילותה היומיומית. היא חוזרת למרכז הבריאות לביקור המשך.
סוגים רבים של חיידקים, פטריות ווירוסים עלולים לגרום לדלקת ריאות. אמוקסיצילין מכוון לפפטידוגליקן של דפנות תאי חיידקים. מכיוון שהאמוקסיצילין לא פתר את הסימפטומים של ברברה, הרשות מסכמת כי הסוכן הסיבתי כנראה חסר פפטידוגליקן, כלומר הפתוגן יכול להיות וירוס, פטרייה או חיידק חסר פפטידוגליקן. אפשרות נוספת היא שהפתוגן הוא חיידק המכיל פפטידוגליקן אך פיתח עמידות לאמוקסיצילין.
תרגיל \(\PageIndex{3}\)
- כיצד יכולה הרשות לזהות באופן סופי את הגורם לדלקת ריאות של ברברה?
- איזו צורת טיפול צריכה הרשות לרשום, בהתחשב בכך שהאמוקסיצילין לא היה יעיל?
נספחים חוטיים
לתאים חיידקיים רבים יש תוספות חלבון המוטמעות בתוך מעטפות התא שלהם המשתרעות כלפי חוץ, ומאפשרות אינטראקציה עם הסביבה. נספחים אלה יכולים להיצמד למשטחים אחרים, להעביר DNA או לספק תנועה. נספחים חוטיים כוללים פימבריה, פילי ודגלים.
פימבריה ופילי
Fimbriae ו- pili דומים מבחינה מבנית ומכיוון שההבחנה בין השניים בעייתית, מונחים אלה משמשים לעתים קרובות לסירוגין. 7 8 המונח fimbriae מתייחס בדרך כלל לחלבונים דמויי זיפים קצרים המוקרנים מעל פני התא במאות. Fimbriae מאפשרים לתא להיצמד למשטחים ולתאים אחרים. עבור חיידקים פתוגניים, היצמדות לתאי מארח חשובה להתיישבות, זיהומיות וארסיות. דבקות במשטחים חשובה גם ביצירת ביופילם.
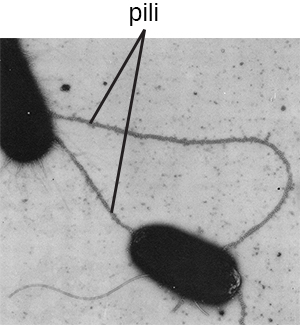
המונח פילי (יחיד: פילוס) מתייחס בדרך כלל לתוספות חלבון ארוכות יותר ופחות רבות המסייעות בהתקשרות למשטחים (איור\(\PageIndex{19}\)). סוג מסוים של פילוס, הנקרא F pilus או sex pilus, חשוב בהעברת ה- DNA בין תאי חיידקים, המתרחש בין בני אותו דור כאשר שני תאים מעבירים או מחליפים פיזית חלקים מהגנום שלהם (ראה כיצד פרוקריוטים א-מיניים משיגים גיוון גנטי).
לפני שהמבנה והתפקוד של המרכיבים השונים במעטפת התא החיידקי הובנו היטב, מדענים כבר השתמשו במאפייני מעטפת התא כדי לסווג חיידקים. בשנת 1933, רבקה לנספילד הציעה שיטה לסרוטיפ של זנים β-המוליטיים שונים של מיני סטרפטוקוקוס באמצעות בדיקת צבירה, טכניקה המשתמשת בגוש חיידקים לאיתור אנטיגנים ספציפיים על פני התא. בכך גילה לנספילד שקבוצה אחת של S. pyogenes, שנמצא בקבוצה A, היה קשור למגוון מחלות אנושיות. היא קבעה שניתן להבחין בין זנים שונים של דלקת קבוצה A זה מזה בהתבסס על וריאציות בחלבונים ספציפיים על פני התא שהיא כינתה חלבוני M.
כיום זוהו יותר מ -80 זנים שונים של דלקת קבוצה A על בסיס חלבוני M. זנים שונים של דלקת קבוצה A קשורים למגוון רחב של זיהומים בבני אדם, כולל דלקת הלוע סטרפטוקוקלית (דלקת גרון), אימפטיגו, תסמונת הלם רעיל, קדחת ארגמנית, קדחת שגרונית ודלקת נמק. חלבון M הוא גורם ארסיות חשוב לדלקת קבוצה A, המסייע לזנים אלה להתחמק ממערכת החיסון. נראה כי שינויים בחלבוני M משנים את ההדבקה של זן מסוים של דלקת קבוצה A.
פלאגלה
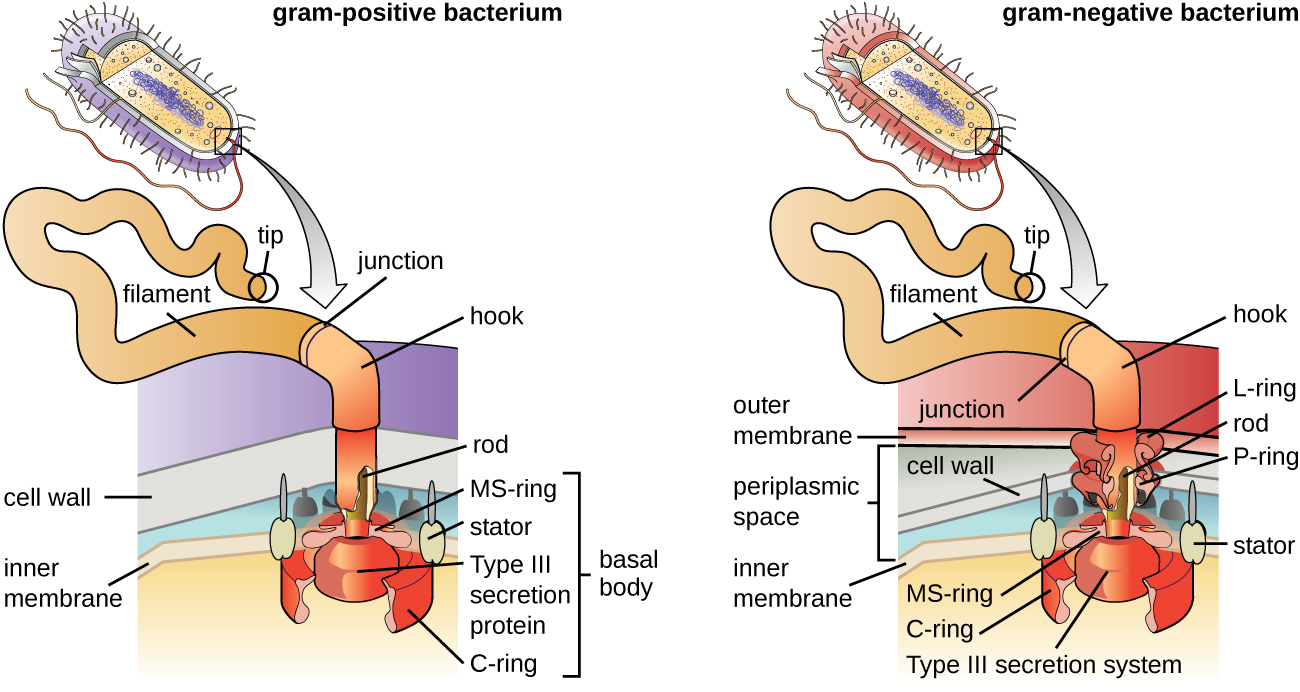
Flagella הם מבנים המשמשים תאים לנוע בסביבות מימיות. דגלים חיידקיים מתנהגים כמו מדחפים. הם חוטים ספירליים נוקשים המורכבים מיחידות משנה של חלבון פלגלין המשתרעות החוצה מהתא ומסתובבות בתמיסה. הגוף הבסיסי הוא המנוע של הדגל והוא מוטבע בקרום הפלזמה (איור\(\PageIndex{20}\)). אזור וו מחבר את הגוף הבסיסי לנימה. לחיידקים גראם חיוביים וגרם שליליים יש תצורות גוף בסיסיות שונות עקב הבדלים במבנה דופן התא.
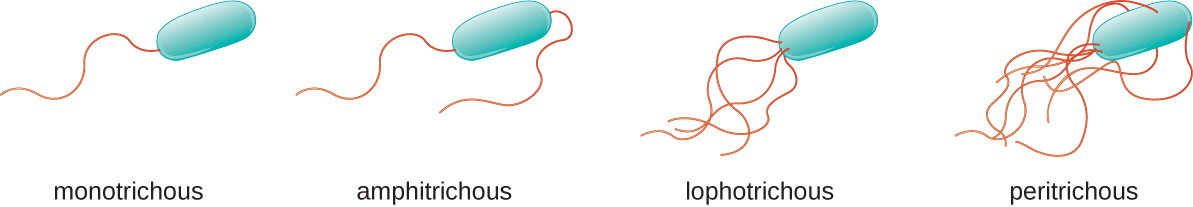
סוגים שונים של חיידקים תנועתיים מציגים סידורים שונים של דגלים (איור\(\PageIndex{21}\)). לחיידק בעל דגל יחיד, הממוקם בדרך כלל בקצה אחד של התא (קוטבי), אומרים שיש לו דגל חד-פעמי. דוגמה לפתוגן חיידקי בעל דגל חד-פעמי הוא Vibrio cholerae, החיידק הגראם-שלילי הגורם לכולרה. לתאים עם דגלים אמפיטריכיים יש דגל או ציציות של דגלים בכל קצה. דוגמה לכך היא ספירילום מינור, הגורם לחום עקיצת עכברוש ספירילרי (אסייתי) או סודוקו. לתאים עם דגלונים לופוטריכיים יש ציצית בקצה אחד של התא. לבצילוס הגראם-שלילי Pseudomonas aeruginosa, פתוגן אופורטוניסטי הידוע בגרימת זיהומים רבים, כולל "אוזן השחיין" ודלקות בפצעי כוויות, יש דגלים לופוטריכיים. דגלים המכסים את כל פני השטח של תא חיידקי נקראים דגלים פריטריכיים. החיידק הגראם-שלילי E. coli מראה סידור פריטריצ'י של דגלים.
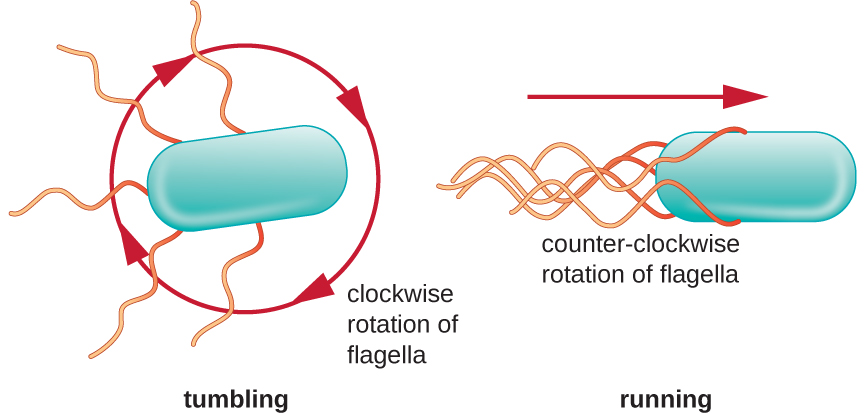
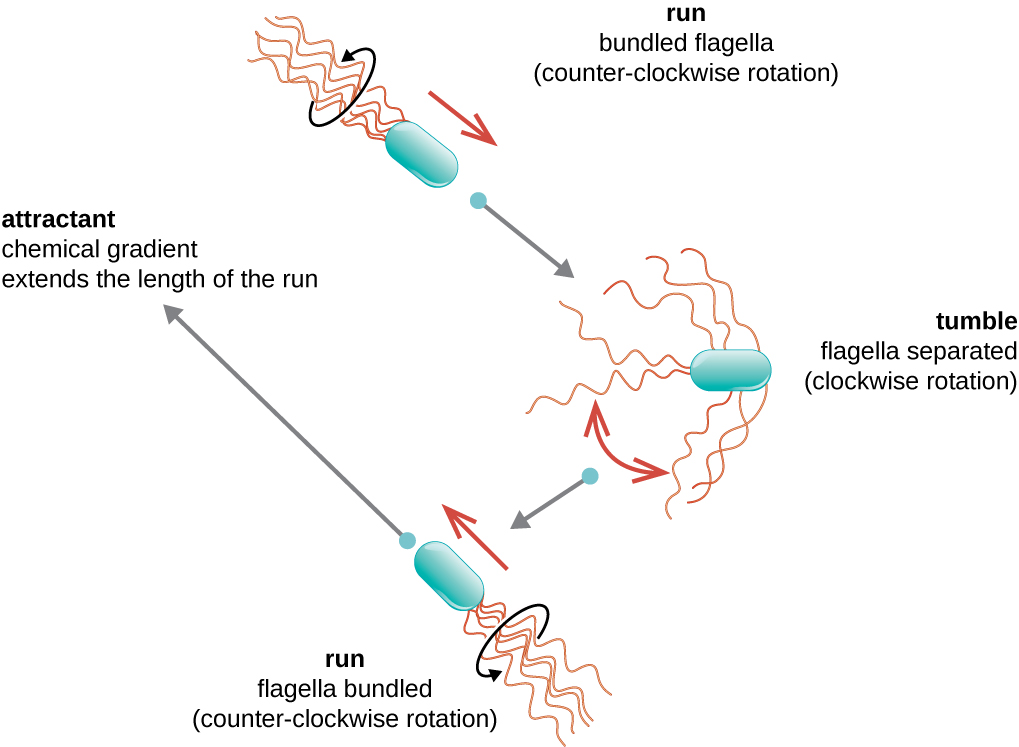
תנועה כיוונית תלויה בתצורה של flagella. חיידקים יכולים לנוע בתגובה למגוון אותות סביבתיים, כולל אור (פוטוטקסיס), שדות מגנטיים (מגנטוטקסיס) באמצעות מגנטוזומים, ולרוב, שיפועים כימיים (כימוטקסיס). תנועה תכליתית לעבר חומר משיכה כימי, כמו מקור מזון, או הרחק מחומר דוחה, כמו כימיקל רעיל, מושגת על ידי הגדלת אורך הריצות והקטנת אורך הנפילות. בעת ריצה, דגלים מסתובבים בכיוון נגד כיוון השעון, ומאפשרים לתא החיידק לנוע קדימה. בחיידק פריטריצ'י, הדגלים כולם מקובצים יחד בצורה יעילה מאוד (איור\(\PageIndex{22}\)), ומאפשרים תנועה יעילה. בעת נפילה, דגלים מפוזרים החוצה תוך כדי סיבוב בכיוון השעון, יוצרים תנועת לולאה ומונעים תנועה קדימה משמעותית אך מכוונים מחדש את התא לכיוון המשיכה. כאשר קיים חומר משיכה, עדיין מתרחשות ריצות ונפילות; עם זאת, אורך הריצות ארוך יותר, בעוד שאורך הנפילות מצטמצם, מה שמאפשר תנועה כוללת לעבר הריכוז הגבוה יותר של חומר המשיכה. כאשר לא קיים שיפוע כימי, אורכי הריצות והנפילות שווים יותר, והתנועה הכוללת אקראית יותר (איור\(\PageIndex{23}\)).
תרגיל \(\PageIndex{4}\)
- מהי שכבת הפפטידוגליקן וכיצד היא שונה בין חיידקים גרם חיוביים לגרם שליליים?
- השווה והשוואה בין דגלים מונוטריכיים, אמפיטריכיים, לופוטריכיים ופריטריכיים.
סיכום
- תאים פרוקריוטיים נבדלים מתאים אוקריוטיים בכך שהחומר הגנטי שלהם כלול בגרעין נוקלאואיד ולא בגרעין הקשור לממברנה. בנוסף, לתאים פרוקריוטים חסרים בדרך כלל אברונים הקשורים לממברנה.
- תאים פרוקריוטיים מאותו מין חולקים בדרך כלל מורפולוגיה דומה של תאים וסידור תאי.
- לרוב התאים הפרוקריוטים יש דופן תא המסייעת לאורגניזם לשמור על מורפולוגיה תאית ומגן עליה מפני שינויים בלחץ האוסמוטי.
- מחוץ לנוקלאואיד, תאים פרוקריוטים עשויים להכיל DNA חוץ-כרומוזומלי בפלסמידים.
- לריבוזומים פרוקריוטיים שנמצאים בציטופלזמה יש גודל של 70S.
- לחלק מהתאים הפרוקריוטים יש תכלילים המאחסנים חומרים מזינים או כימיקלים לשימושים אחרים.
- חלק מהתאים הפרוקריוטים מסוגלים ליצור אנדוספורים באמצעות ספורולציה כדי לשרוד במצב רדומה כאשר התנאים אינם נוחים. אנדוספורים יכולים לנבוט, ולהפוך בחזרה לתאים וגטטיביים כאשר התנאים משתפרים.
- בתאים פרוקריוטים מעטפת התא כוללת קרום פלזמה ובדרך כלל דופן תא.
- ממברנות חיידקיות מורכבות מפוספוליפידים עם חלבונים אינטגרליים או היקפיים. מרכיבי חומצות השומן של פוספוליפידים אלה קשורים לאסטר ומשמשים לעתים קרובות לזיהוי סוגים ספציפיים של חיידקים. החלבונים משרתים מגוון פונקציות, כולל הובלה, תקשורת בין תא לתא וחישת תנאים סביבתיים. ממברנות ארכאיות נבדלות בכך שהן מורכבות מחומצות שומן המקושרות לאתר לפוספוליפידים.
- חלק מהמולקולות יכולות לנוע על פני קרום החיידק על ידי דיפוזיה פשוטה, אך רוב המולקולות הגדולות חייבות להיות מועברות באופן פעיל דרך מבני ממברנה באמצעות אנרגיה תאית.
- קירות תאים פרוקריוטיים עשויים להיות מורכבים מפפטידוגליקן (חיידקים) או פסאודופפטידוגליקן (ארכאה).
- תאי חיידקים גראם חיוביים מאופיינים בשכבת פפטידוגליקן עבה, בעוד שתאי חיידקים גרם שליליים מאופיינים בשכבת פפטידוגליקן דקה המוקפת בקרום חיצוני.
- חלק מהתאים הפרוקריוטים מייצרים ציפויי גליקוקליקס, כגון כמוסות ושכבות רפש, המסייעים בהתקשרות למשטחים ו/או התחמקות ממערכת החיסון המארחת.
- בחלק מהתאים הפרוקריוטים יש פימבריה או פילי, תוספות חוטיות המסייעות בהתקשרות למשטחים. פילי משמשים גם בהעברת חומר גנטי בין תאים.
- חלק מהתאים הפרוקריוטים משתמשים בפללה אחת או יותר כדי לנוע במים. חיידקים פריטריכיים, שיש להם דגלים רבים, משתמשים בריצות ונפילות כדי לנוע בכוונה לכיוון של חומר משיכה כימי.
הערות שוליים
- 1 י.-ח. מ. צ'אן, ו.פ. מרשל. "מאפייני קנה מידה של גודל התא והאברון." אורגנוגנזה 6 מס '2 (2010) :88—96.
- 2 פ 'רוטפוס, מ' בנדר, ר קונרד. "הישרדות ופעילות של חיידקים במשקע אגם עמוק ומיושן (אגם קונסטנץ)." אקולוגיה מיקרוביאלית 33 מס '1 (1997) :69—77.
- 3 ר 'סינקלייר ואח '. "התמדה של סוכנים נבחרים בקטגוריה A בסביבה." מיקרוביולוגיה יישומית וסביבתית 74 מס '3 (2008) :555-563.
- 4 טי ג'יי סילהאבי, ד 'קאנה, ס 'ווקר. "מעטפת התא החיידקי." פרספקטיבות קולד ספרינג הארבור בביולוגיה 2 מס '5 (2010): a000414.
- 5 ב 'צובר ואח '. "שכבה גרגירית במרחב הפריפלזמי של חיידקים גראם חיוביים ומבנים עדינים של Enterococcus gallinarum ו- Streptococcus gordonii Septa נחשף על ידי מיקרוסקופיה קריו-אלקטרון של קטעי זגוגית." כתב העת לבקטריולוגיה 188 מס '18 (2006) :6652—6660
- 6 ל 'גאנה, ס 'צ'נה, ג'י ג'יי ג'נסנה. "ארגון מולקולרי של פפטידוגליקן גראם-שלילי." הליכי האקדמיה הלאומית למדעים של ארצות הברית של אמריקה 105 מס '48 (2008): 18953—18957.
- 7 ג'יי גרנטה ואח '. "תובנות מבניות לגבי הביוגנזה והיווצרות הביופילם על ידי הפילוס הנפוץ של Escherichia coli." הליכי האקדמיה הלאומית למדעים של ארצות הברית של אמריקה 109 מס '10 (2012): 3950—3955.
- 8 טי פרופט, א.נ. בייקר. "פילי בחיידקים גראם-שליליים וגרם-חיוביים - מבנה, הרכבה ותפקידם במחלות." מדעי החיים הסלולריים והמולקולריים 66 (2009) :613.