
3.4: מאפיינים ייחודיים של תאים אוקריוטיים
- Page ID
- 208903

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)מטרות למידה
- הסבר את המאפיינים המבדילים של תאים אוקריוטיים
- תאר מבנים פנימיים וחיצוניים של תאים פרוקריוטים במונחים של המבנה הפיזי שלהם, המבנה הכימי והתפקוד שלהם
- לזהות ולתאר מבנים ואברונים ייחודיים לתאים אוקריוטיים
- השווה והשוואה בין מבנים דומים המצויים בתאים פרוקריוטיים ואיקריוטים
אורגניזמים אוקריוטיים כוללים פרוטוזואנים, אצות, פטריות, צמחים ובעלי חיים. חלק מהתאים האיקריוטים הם מיקרואורגניזמים חד תאיים עצמאיים, בעוד שאחרים הם חלק מאורגניזמים רב-תאיים. לתאים של אורגניזמים אוקריוטיים יש כמה מאפיינים מובחנים. מעל לכל, תאים אוקריוטיים מוגדרים על ידי נוכחות של גרעין המוקף בקרום גרעיני מורכב. כמו כן, תאים אוקריוטיים מאופיינים בנוכחות אברונים הקשורים לקרום בציטופלזמה. אברונים כגון מיטוכונדריה, הרשת האנדופלזמית (ER), מנגנון גולגי, ליזוזומים ופרוקסיזומים מוחזקים במקומם על ידי הציטוס שלד, רשת פנימית התומכת בהובלה של רכיבים תוך תאיים ומסייעת בשמירה על צורת התא (איור). \(\PageIndex{1}\) הגנום של תאים אוקריוטיים ארוז בכרומוזומים מרובים בצורת מוט בניגוד לכרומוזום היחיד בצורת מעגל המאפיין את רוב התאים הפרוקריוטים. הטבלה \(\PageIndex{1}\) משווה את המאפיינים של מבני תאים אוקריוטיים לאלה של חיידקים וארכיאה.

| מבנה התא | פרוקריוטים | אוקריוטים | |
|---|---|---|---|
| חיידקים | ארכאה | ||
| גודל | ~ 0.5-1 מיקרומטר | ~ 0.5-1 מיקרומטר | ~ 5—20 מיקרומטר |
| יחס שטח פנים לנפח | גבוה | גבוה | נמוך |
| גרעין | לא | לא | הן |
| מאפייני הגנום |
|
|
|
| חלוקת תאים | ביקוע בינארי | ביקוע בינארי | מיטוזיס, מיוזה |
| הרכב שומנים ממברנה |
|
|
|
| הרכב דופן התא |
|
|
|
| מבני תנועתיות | סמלונים ספירליים קשיחים המורכבים מפלגלין | דגלונים ספירליים קשיחים המורכבים מפלגלינים ארכאיים | דגלים גמישים וסיליה המורכבים ממיקרוטובולים |
| אברונים הקשורים לממברנה | לא | לא | הן |
| מערכת אנדוממברנה | לא | לא | כן (ER, גולגי, ליזוזומים) |
| ריבוזומים | שנות ה -70 | שנות ה -70 |
|
מורפולוגיות תאים
תאים אוקריוטיים מציגים מגוון רחב של מורפולוגיות תאים שונות. צורות אפשריות כוללות כדורית, ביצית, קובואידית, גלילית, שטוחה, עדשה, פיוזיפורמית, דיסקואידית, סהר, כוכב טבעת ומצולע (איור). \(\PageIndex{2}\) חלק מהתאים האיקריוטים אינם סדירים בצורתם, וחלקם מסוגלים לשנות צורה. צורתו של סוג מסוים של תא אוקריוטי עשויה להיות מושפעת מגורמים כמו תפקידו העיקרי, ארגון השלד שלו, צמיגות הציטופלזמה שלו, קשיחות קרום התא או דופן התא שלו (אם יש לו כזה), והלחץ הפיזי המופעל עליו על ידי הסביבה הסובבת ו/או התאים הסמוכים.
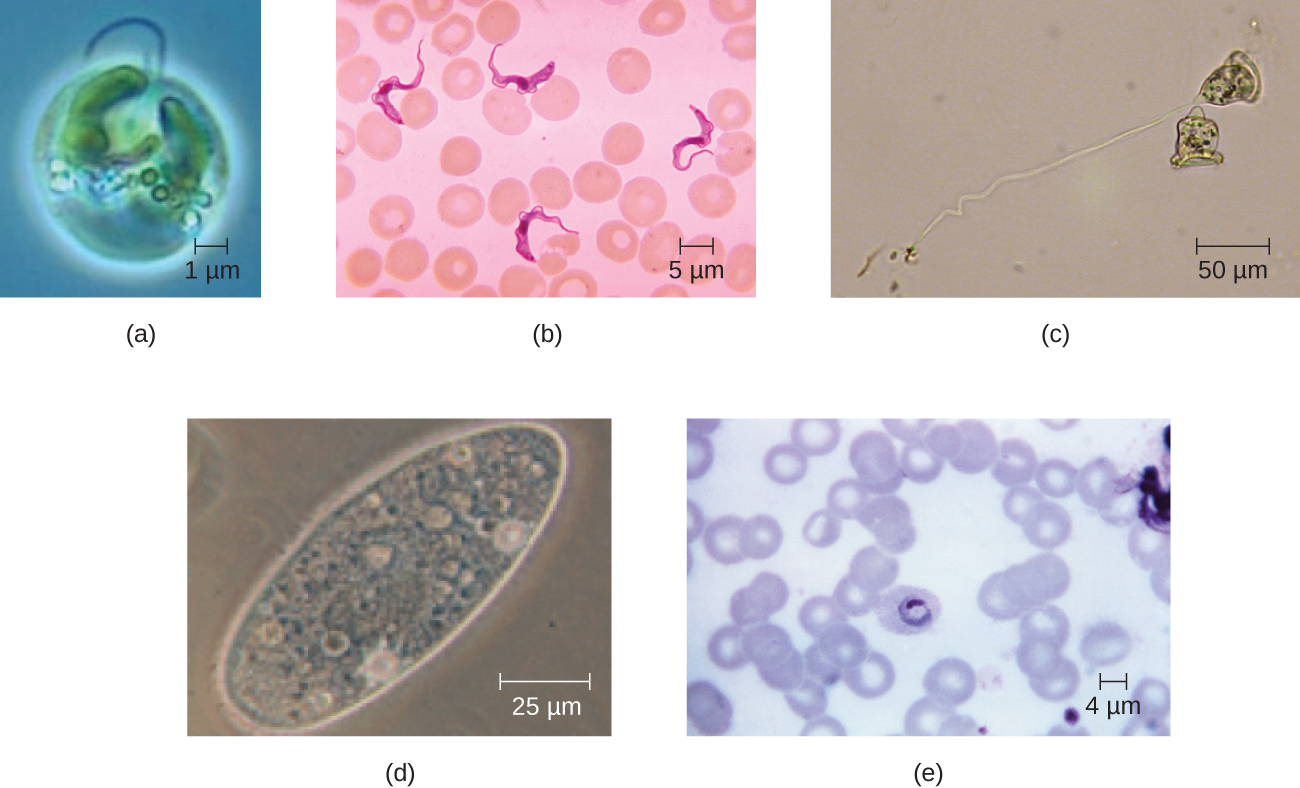
תרגיל \(\PageIndex{1}\)
זהה שני הבדלים בין תא אוקריוטי לפרוקריוטי.
גרעין
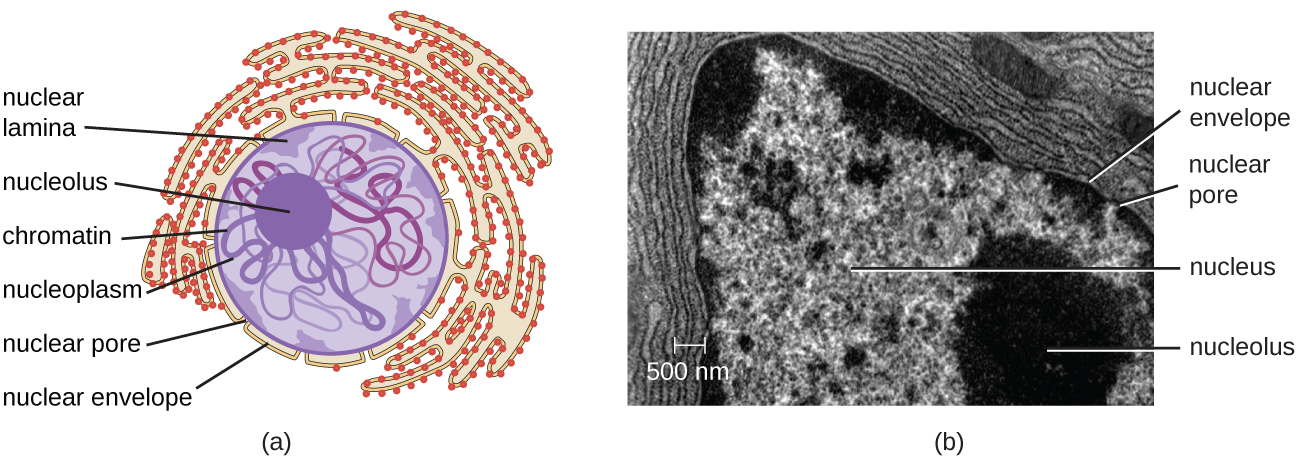
בניגוד לתאים פרוקריוטים, שבהם ה-DNA כלול באופן רופף באזור הנוקלאואיד, לתאים אוקריוטיים יש גרעין, המוקף בממברנה גרעינית מורכבת המאכלסת את גנום ה-DNA (איור\(\PageIndex{3}\)). על ידי מכיל את ה- DNA של התא, הגרעין שולט בסופו של דבר בכל פעילויות התא וגם ממלא תפקיד חיוני ברבייה ותורשה. לתאים אוקריוטיים בדרך כלל ה-DNA שלהם מאורגן לכרומוזומים ליניאריים מרובים. ה- DNA בתוך הגרעין מאורגן ומעובה מאוד כך שיתאים בתוך הגרעין, מה שמתבצע על ידי עטיפת ה- DNA סביב חלבונים הנקראים היסטונים.
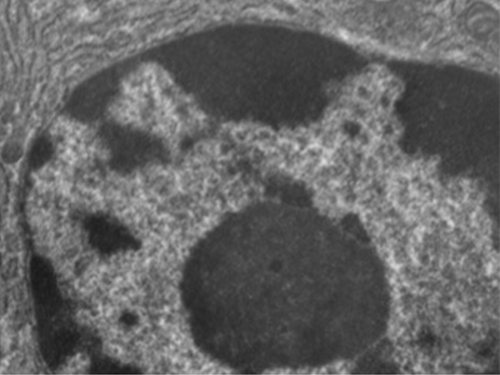
למרות שלרוב התאים האיקריוטים יש גרעין אחד בלבד, קיימים חריגים. לדוגמה, לפרוטוזואנים מהסוג Paramecium יש בדרך כלל שני גרעינים שלמים: גרעין קטן המשמש להתרבות (מיקרו-גרעין) וגרעין גדול המכוון את חילוף החומרים התאי (מקרו -גרעין). בנוסף, חלק מהפטריות יוצרות באופן זמני תאים עם שני גרעינים, הנקראים תאים הטרוקריוטיים, במהלך רבייה מינית. תאים שהגרעינים שלהם מתחלקים, אך הציטופלזמה שלהם לא, נקראים קואנוציטים.
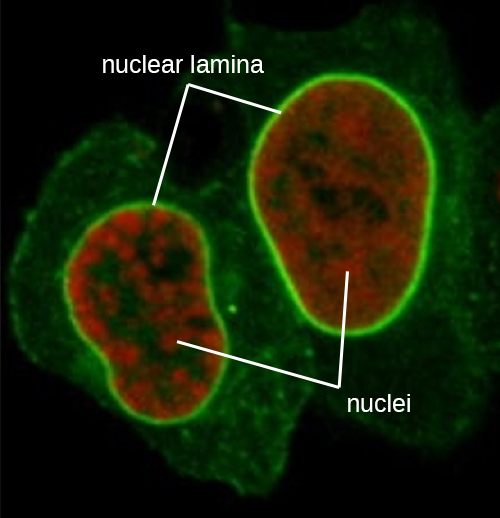
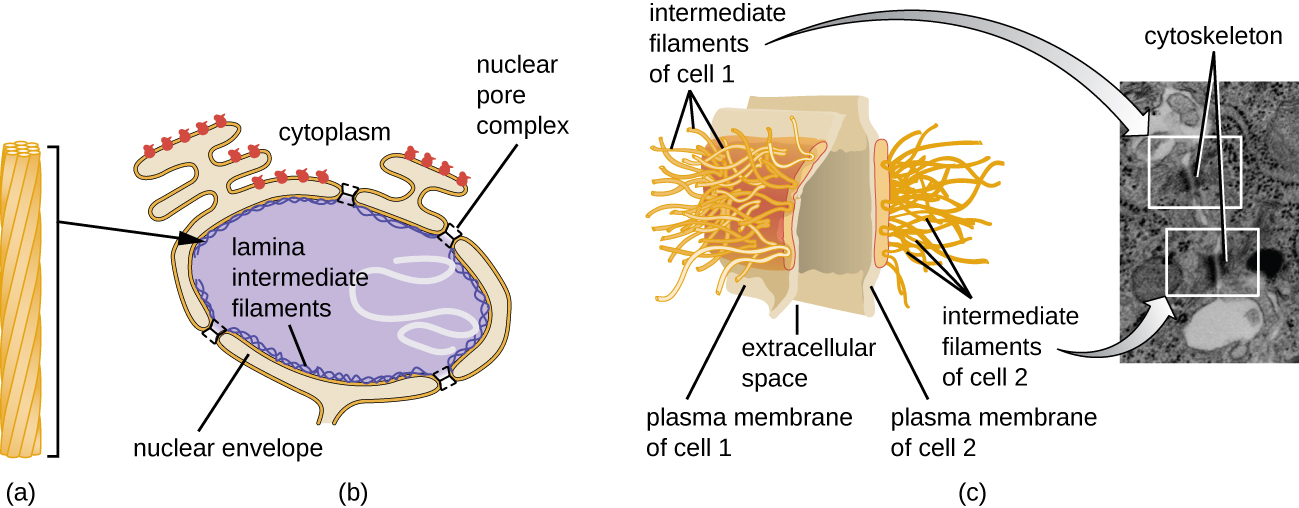
הגרעין קשור בממברנה גרעינית מורכבת, הנקראת לעתים קרובות המעטפת הגרעינית, המורכבת משתי שכבות שומנים נפרדות הצמודות זו לזו (איור). \(\PageIndex{4}\) למרות הקשרים הללו בין הממברנות הפנימיות והחיצוניות, כל קרום מכיל שומנים וחלבונים ייחודיים על פני השטח הפנימיים והחיצוניים שלו. המעטפת הגרעינית מכילה נקבוביות גרעיניות, שהן מתחמי חלבון גדולים בצורת שושנה השולטים בתנועת החומרים אל תוך הגרעין ומחוצה לו. הצורה הכוללת של הגרעין נקבעת על ידי הלמינה הגרעינית, רשת של חוטי ביניים שנמצאים ממש בתוך ממברנות המעטפת הגרעינית. מחוץ לגרעין, חוטי ביניים נוספים יוצרים רשת רופפת יותר ומשמשים לעיגון הגרעין במיקום בתוך התא.
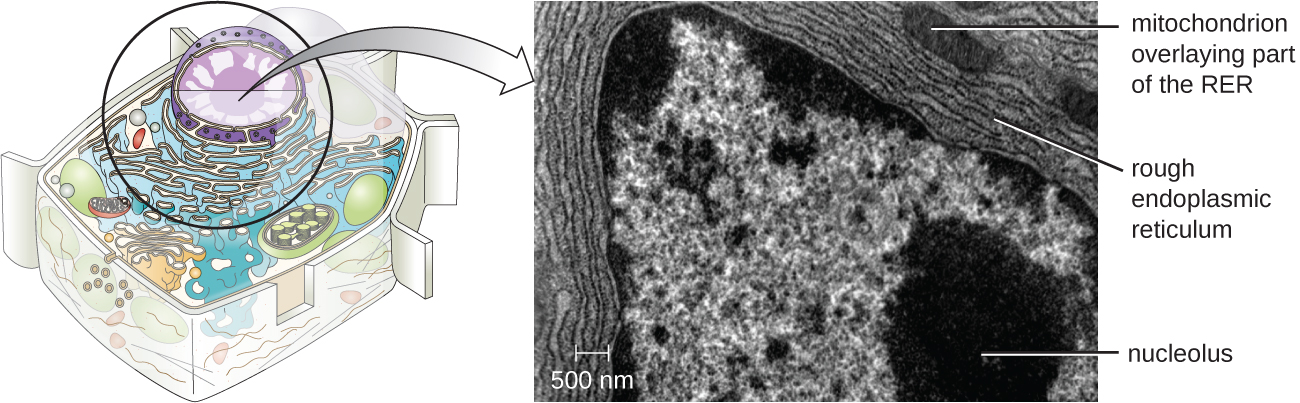
נוקלאולוס
הגרעין הוא אזור צפוף בתוך הגרעין בו מתרחשת ביוסינתזה של RNA ריבוזומלי (rRNA). בנוסף, הגרעין הוא גם האתר בו מתחילה הרכבה של ריבוזומים. קומפלקסים פרה-ריבוזומליים מורכבים מ-rRNA וחלבונים בגרעין; לאחר מכן הם מועברים החוצה לציטופלזמה, שם הושלמה הרכבת הריבוזומים (איור). \(\PageIndex{5}\)
ריבוזומים
לריבוזומים המצויים באברונים אוקריוטיים כגון מיטוכונדריה או כלורופלסטים יש ריבוזומים 70S - באותו גודל כמו ריבוזומים פרוקריוטיים. עם זאת, ריבוזומים שאינם קשורים לאברונים בתאים אוקריוטיים הם ריבוזומים 80S, המורכבים מיחידת משנה קטנה של 40S ויחידת משנה גדולה של 60S. מבחינת גודל והרכב, זה מבדיל אותם מהריבוזומים של תאים פרוקריוטים.
שני סוגי הריבוזומים האוקריוטיים שאינם קשורים לאיברים מוגדרים על פי מיקומם בתא: ריבוזומים חופשיים וריבוזומים הקשורים לממברנה. ריבוזומים חופשיים נמצאים בציטופלזמה ומשמשים לסינתזה של חלבונים מסיסים במים; ריבוזומים הקשורים לממברנה נמצאים מחוברים לרטיקולום האנדופלזמי המחוספס ויוצרים חלבונים להחדרה לקרום התא או חלבונים המיועדים לייצוא מהתא.
ההבדלים בין ריבוזומים אוקריוטיים לפרוקריוטים רלוונטיים מבחינה קלינית מכיוון שתרופות אנטיביוטיות מסוימות נועדו למקד לאחת או השנייה. לדוגמה, ציקלוהקסימיד מכוון לפעולה אוקריוטית, ואילו כלורמפניקול מכוון לריבוזומים פרוקריוטיים. 1 מכיוון שתאים אנושיים הם אוקריוטים, הם בדרך כלל אינם נפגעים מאנטיביוטיקה ההורסת את הריבוזומים הפרוקריוטיים בחיידקים. עם זאת, לפעמים תופעות לוואי שליליות עלולות להתרחש מכיוון שהמיטוכונדריה בתאים אנושיים מכילה ריבוזומים פרוקריוטיים.
מערכת אנדוממברנה
מערכת האנדוממברנה, הייחודית לתאים אוקריוטיים, היא סדרה של צינוריות ממברנות, שקיות ודיסקים שטוחים המסנתזים רכיבי תאים רבים ומניעים חומרים בתוך התא (איור). \(\PageIndex{6}\) בגלל גודל התא הגדול יותר שלהם, תאים אוקריוטיים דורשים ממערכת זו להעביר חומרים שלא ניתן לפזר על ידי דיפוזיה בלבד. מערכת האנדוממברנה כוללת מספר אברונים וקשרים ביניהם, כולל הרשת האנדופלזמית, מנגנון גולגי, ליזוזומים ושלפוחיות.
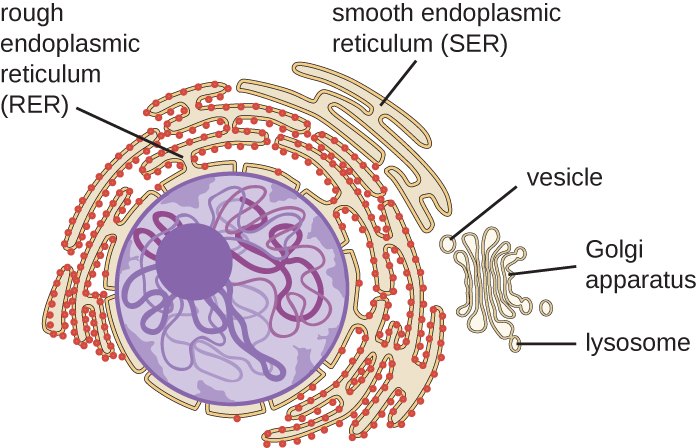
רטיקולום אנדופלזמי
הרטיקולום האנדופלזמי (ER) הוא מערך מחובר זה לזה של צינוריות ובורות מים (שקיות שטוחות) עם דו שכבה שומנית אחת (איור). \(\PageIndex{7}\) החללים שבתוך בורות המים נקראים לומן של המיון. ישנם שני סוגים של ER, רטיקולום אנדופלזמי מחוספס (RER) ורטיקולום אנדופלזמי חלק (SER). שני סוגים שונים אלה של ER הם אתרים לסינתזה של סוגים שונים של מולקולות. RER משובץ בריבוזומים הקשורים בצד הציטופלזמי של הממברנה. ריבוזומים אלה מייצרים חלבונים המיועדים לקרום הפלזמה (איור\(\PageIndex{}\)). לאחר סינתזה, חלבונים אלה מוכנסים לתוך הממברנה של RER. שקים קטנים של ה-RER המכילים את החלבונים החדשים שסונתזו לאחר מכן יוצאים כשלפוחיות הובלה ועוברים למנגנון גולגי לעיבוד נוסף, ישירות לקרום הפלזמה, לקרום של אברון אחר, או אל מחוץ לתא. שלפוחיות הובלה הן כדורים חד-שכבתיים, דו-שכבתיים, קרומיים עם פנים חלולים הנושאים מולקולות. ל- SER אין ריבוזומים ולכן נראה "חלק". הוא מעורב בביוסינתזה של שומנים, מטבוליזם של פחמימות וניקוי רעלים של תרכובות רעילות בתוך התא.
מכשיר גולגי
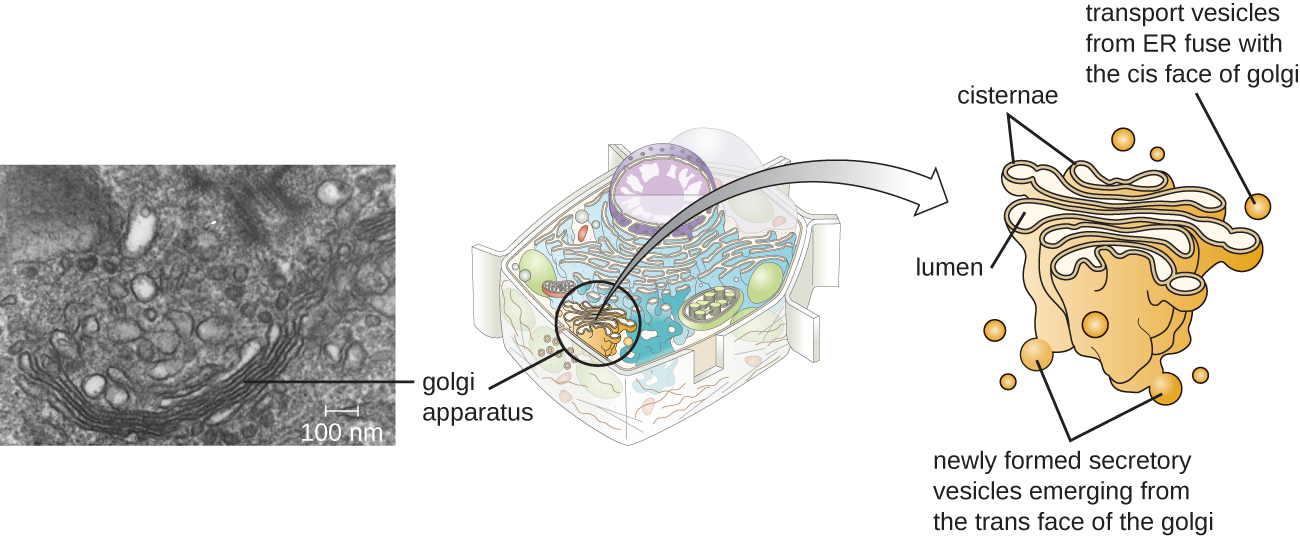
מנגנון גולגי התגלה בתוך מערכת האנדוממברנה בשנת 1898 על ידי המדען האיטלקי קמילו גולגי (1843—1926), שפיתח טכניקת צביעה חדשה שהראתה מבני קרום מוערמים בתוך תאי פלסמודיום, הגורם הסיבתי למלריה. מנגנון גולגי מורכב מסדרה של דיסקים קרומיים הנקראים דיקטוזומים, שלכל אחד מהם יש דו שכבה שומנית אחת, הנערמים יחד (איור). \(\PageIndex{8}\)
אנזימים במנגנון גולגי משנים שומנים וחלבונים המועברים מה- ER לגולגי, ולעתים קרובות מוסיפים להם רכיבי פחמימות, מייצרים גליקוליפידים, גליקופרוטאינים או פרוטאוגליקנים. גליקוליפידים וגליקופרוטאינים מוכנסים לעיתים קרובות לקרום הפלזמה וחשובים לזיהוי אותות על ידי תאים אחרים או חלקיקים זיהומיים. ניתן להבחין בין סוגים שונים של תאים זה מזה על ידי המבנה והסידור של הגליקוליפידים והגליקופרוטאינים הכלולים בקרומי הפלזמה שלהם. גליקוליפידים וגליקופרוטאינים אלה משמשים בדרך כלל גם כקולטני פני התא.
שלפוחיות הובלה עוזבות את נתיך ה- ER עם מכשיר גולגי על פניו המקבלים, או cis. החלבונים מעובדים בתוך מנגנון גולגי, ולאחר מכן שלפוחיות הובלה נוספות המכילות את החלבונים והשומנים שהשתנו צובטים ממנגנון גולגי על פניו היוצאות, או הטרנס. שלפוחיות יוצאות אלה עוברות ומתמזגות עם קרום הפלזמה או הממברנה של אברונים אחרים.
אקסוציטוזיס הוא התהליך שבו שלפוחיות הפרשה (שקיות קרומיות כדוריות) משחררות את תוכנן לחלק החיצוני של התא (איור). \(\PageIndex{8}\) לכל התאים יש מסלולי הפרשה מכוננים בהם שלפוחיות הפרשה מעבירות חלבונים מסיסים המשתחררים מהתא ללא הרף (מכוננת). לתאים מיוחדים מסוימים יש גם מסלולי הפרשה מווסתים, המשמשים לאחסון חלבונים מסיסים בשלפוחיות הפרשה. הפרשה מוסדרת כוללת חומרים שמשתחררים רק בתגובה לאירועים או אותות מסוימים. לדוגמה, תאים מסוימים של מערכת החיסון האנושית (למשל, תאי פיטום) מפרישים היסטמין בתגובה לנוכחות של חפצים זרים או פתוגנים בגוף. היסטמין הוא תרכובת המפעילה מנגנונים שונים המשמשים את המערכת החיסונית לחיסול פתוגנים.
ליזוזומים
בשנות השישים גילה המדען הבלגי כריסטיאן דה דוב (1917—2013) ליזוזומים, אברונים הקשורים לממברנה של מערכת האנדוממברנה המכילים אנזימי עיכול. סוגים מסוימים של תאים אוקריוטיים משתמשים בליזוזומים כדי לפרק חלקיקים שונים, כגון מזון, אברונים פגומים או פסולת תאית, מיקרואורגניזמים או קומפלקסים חיסוניים. מידור אנזימי העיכול בתוך הליזוזום מאפשר לתא לעכל חומר ביעילות מבלי לפגוע ברכיבים הציטופלזמיים של התא.
תרגיל \(\PageIndex{2}\)
תן שם למרכיבי מערכת האנדוממברנה ותאר את הפונקציה של כל רכיב.
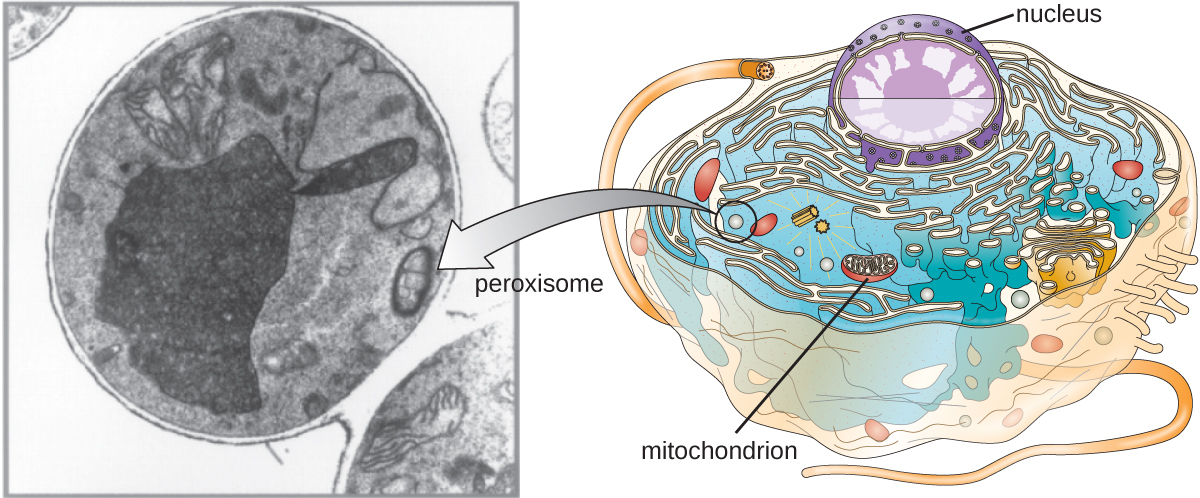
פרוקסיזומים
כריסטיאן דה דוב זוכה גם לגילוי פרוקסיזומים, אברונים הקשורים לממברנה שאינם חלק ממערכת האנדוממברנה (איור). \(\PageIndex{9}\) פרוקסיזומים נוצרים באופן עצמאי בציטופלזמה מסינתזה של חלבוני פרוקסין על ידי ריבוזומים חופשיים ושילוב חלבוני פרוקסין אלה בפרוקסיזומים קיימים. גידול פרוקסיזומים מתחלק ואז בתהליך הדומה לביקוע בינארי.
פרוקסיזומים נקראו לראשונה בזכות יכולתם לייצר מי חמצן, מולקולה תגובתית מאוד המסייעת בפירוק מולקולות כגון חומצת שתן, חומצות אמינו וחומצות שומן. לפרוקסיזומים יש גם את האנזים קטלאז, שעלול לפרק מי חמצן. יחד עם ה- SER, לפרוקסיזומים יש תפקיד גם בביוסינתזה של שומנים. כמו ליזוזומים, המידור של מולקולות השפלה אלה בתוך אברון מסייע בהגנה על התוכן הציטופלזמי מפני נזק לא רצוי.
הפרוקסיזומים של אורגניזמים מסוימים מתמחים בכדי לענות על צרכיהם התפקודיים הספציפיים. לדוגמה, גליוקסיזומים הם פרוקסיזומים משתנים של שמרים ותאי צמחים המבצעים מספר פונקציות מטבוליות, כולל ייצור מולקולות סוכר. באופן דומה, גליקוזומים הם פרוקסיזומים משתנים המיוצרים על ידי טריפנוזומים מסוימים, הפרוטוזואנים הפתוגניים הגורמים למחלת צ'אגס ולמחלת שינה אפריקאית.
שלד
לתאים אוקריוטיים יש שלד פנימי העשוי ממיקרופילמנטים, חוטי ביניים ומיקרוטובולים. מטריצה זו של סיבים וצינורות מספקת תמיכה מבנית וכן רשת שעליה ניתן להעביר חומרים בתוך התא ועליה ניתן לעגן אברונים (איור\(\PageIndex{10}\)). לדוגמה, תהליך האקסוציטוזיס כרוך בתנועה של שלפוחית דרך הרשת הציטו-שלד אל קרום הפלזמה, שם היא יכולה לשחרר את תוכנה.
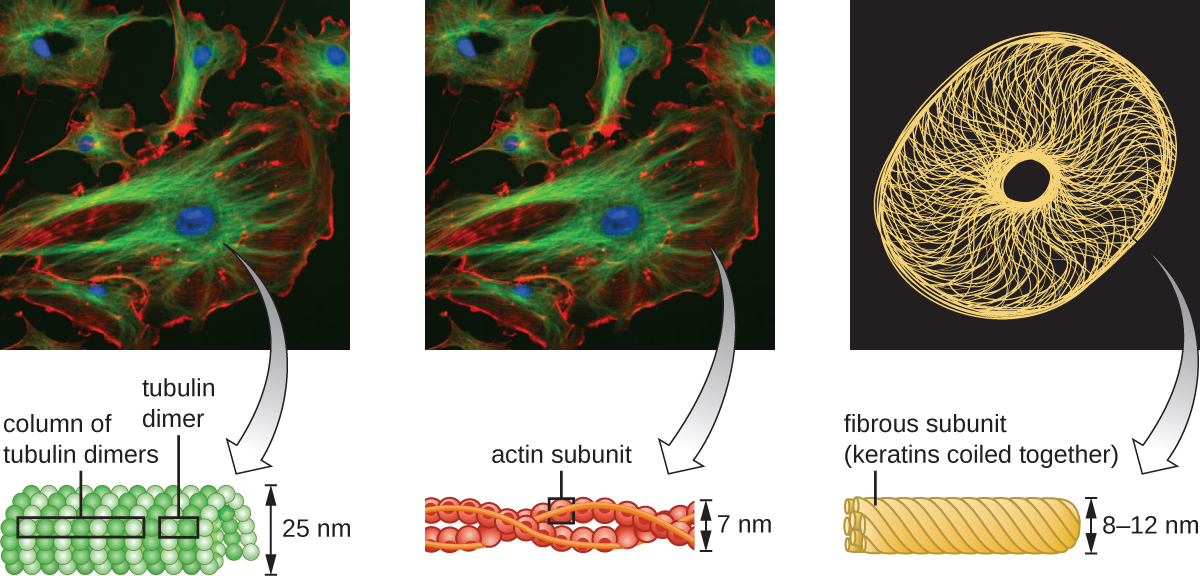
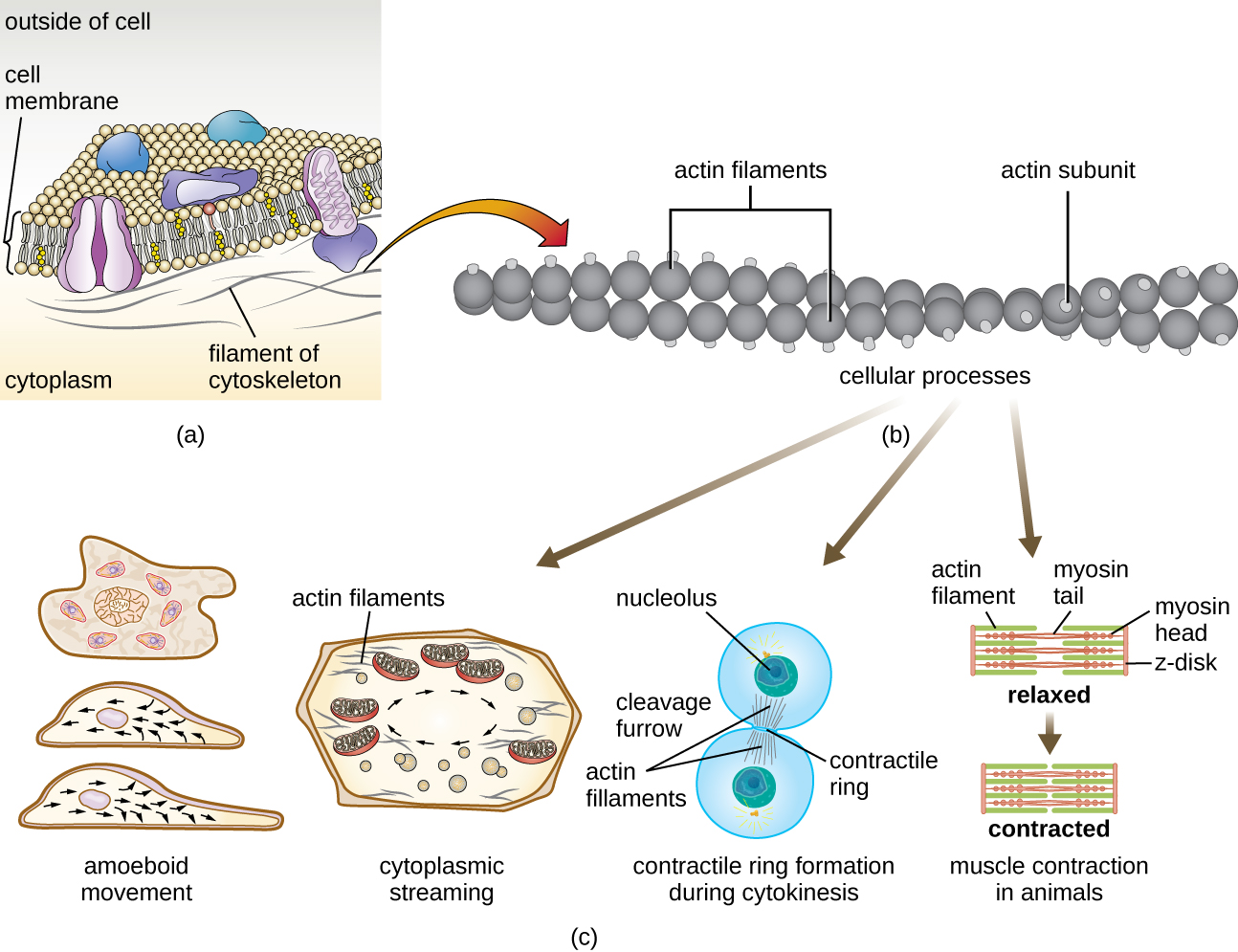
מיקרופילמנטים מורכבים משני גדילי אקטין שזורים זה בזה, שכל אחד מהם מורכב ממונומרים אקטין היוצרים כבלים חוטים בקוטר 6 ננומטר 2 (איור). \(\PageIndex{11}\) חוטי האקטין פועלים יחד עם חלבונים מוטוריים, כמו מיוסין, כדי להשפיע על התכווצות שרירים בבעלי חיים או על התנועה האמבואידית של כמה חיידקים אוקריוטיים. באורגניזמים אמבואידים ניתן למצוא אקטין בשתי צורות: צורת ג'ל נוקשה יותר, מפולמרית, וצורה מסיסה נוזלית יותר ולא פולימרית. אקטין בצורת הג'ל יוצר יציבות באקטופלזמה, האזור דמוי הג'ל של הציטופלזמה ממש בתוך קרום הפלזמה של פרוטוזואנים אמבואידים.
הרחבות זמניות של הממברנה הציטופלזמית הנקראת פסאודופודיה (כלומר "רגליים כוזבות") מיוצרות באמצעות זרימה קדימה של חוטי אקטין מסיסים לתוך הפסאודופודיה, ואחריה מחזור הג'ל-סול של חוטי האקטין, וכתוצאה מכך תנועתיות התא. ברגע שהציטופלזמה משתרעת החוצה, ויוצרת פסאודופודיום, הציטופלזמה הנותרת זורמת למעלה כדי להצטרף לקצה המוביל, ובכך יוצרת תנועה קדימה. מעבר לתנועה אמבואידית, מיקרופילמנטים מעורבים גם במגוון תהליכים אחרים בתאים אוקריוטיים, כולל זרימה ציטופלזמית (תנועה או זרימת ציטופלזמה בתוך התא), היווצרות תלם מחשוף במהלך חלוקת התא ותנועת שרירים בבעלי חיים (איור). \(\PageIndex{11}\) פונקציות אלו הן תוצאה של האופי הדינמי של מיקרופילמנטים, שיכולים לפלמר ולדה-פולימריזציה בקלות יחסית בתגובה לאותות סלולריים, והאינטראקציות שלהם עם מנועים מולקולריים בסוגים שונים של תאים אוקריוטיים.
חוטי ביניים (איור\(\PageIndex{12}\)) הם קבוצה מגוונת של חוטים ציטוסקלטליים הפועלים ככבלים בתוך התא. הם מכונים "ביניים" מכיוון שקוטר 10 ננומטר שלהם עבה יותר מזה של אקטין אך דק יותר מזה של המיקרוטובולים. 3 הם מורכבים מכמה גדילים של יחידות משנה פולימריות, אשר בתורן מורכבות ממגוון רחב של מונומרים. חוטי ביניים נוטים להיות קבועים יותר בתא ולשמור על מיקום הגרעין. הם גם יוצרים את הלמינה הגרעינית (בטנה או שכבה) ממש בתוך המעטפת הגרעינית. בנוסף, חוטי ביניים ממלאים תפקיד בעיגון תאים יחד ברקמות בעלי חיים. חלבון נימה הביניים דסמין נמצא בדסמוזומים, מבני החלבון המחברים יחד תאי שריר ועוזרים להם להתנגד לכוחות פיזיים חיצוניים. חלבון נימה הביניים קרטין הוא חלבון מבני המצוי בשיער, בעור ובציפורניים.
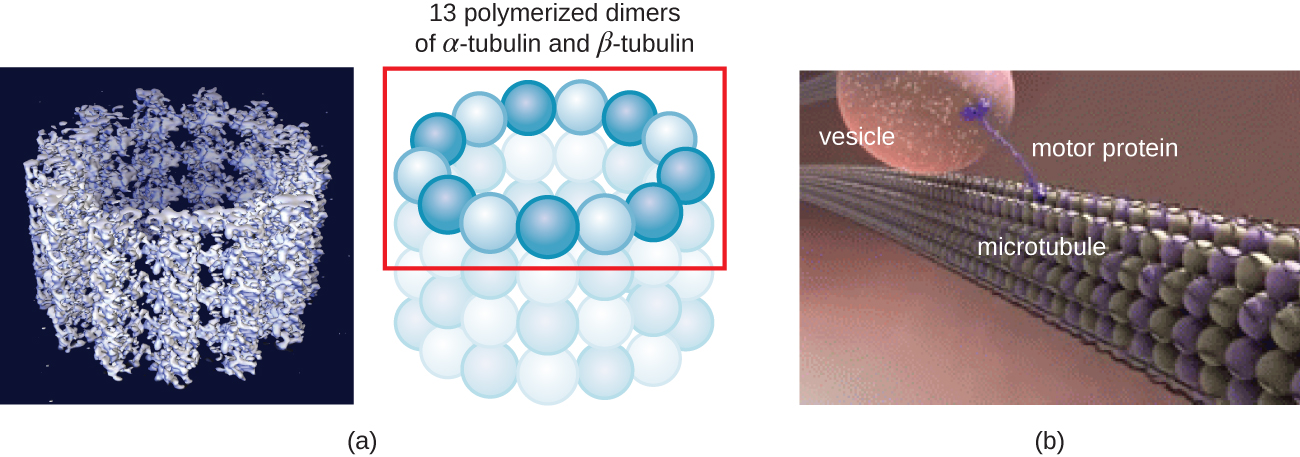
מיקרוטובולים (איור\(\PageIndex{13}\)) הם סוג שלישי של סיבים ציטו-שלד המורכבים מדימרים טובולין (α טובולין ו β טובולין). אלה יוצרים צינורות חלולים בקוטר 23 ננומטר המשמשים כקורות בתוך השלד. 4 בדומה למיקרופילמנטים, המיקרוטובולים הם דינמיים ובעלי יכולת להרכיב ולהתפרק במהירות. מיקרוטובולים עובדים גם עם חלבונים מוטוריים (כגון דינין וקינסין) כדי להזיז אברונים ושלפוחיות בתוך הציטופלזמה. בנוסף, מיקרוטובולים הם המרכיבים העיקריים של דגלים אוקריוטיים וסיליה, המרכיבים הן את החוט והן את מרכיבי הגוף הבסיסיים (איור). \(\PageIndex{20}\)
בנוסף, מיקרו-צינורות מעורבים בחלוקת תאים ויוצרים את הציר המיטוטי המשמש להפרדת כרומוזומים במהלך מיטוזה ומיוזה. הציר המיטוטי מיוצר על ידי שני צנטרוזומים, שהם בעצם מרכזים מארגנים מיקרו-צינוריות, בקצוות מנוגדים של התא. כל צנטרוזום מורכב מזוג צנטריולים הממוקמים בזווית ישרה זה לזה, וכל צנטריול הוא מערך של תשעה מיקרוטובולים מקבילים המסודרים בשלישיות (איור). \(\PageIndex{14}\)
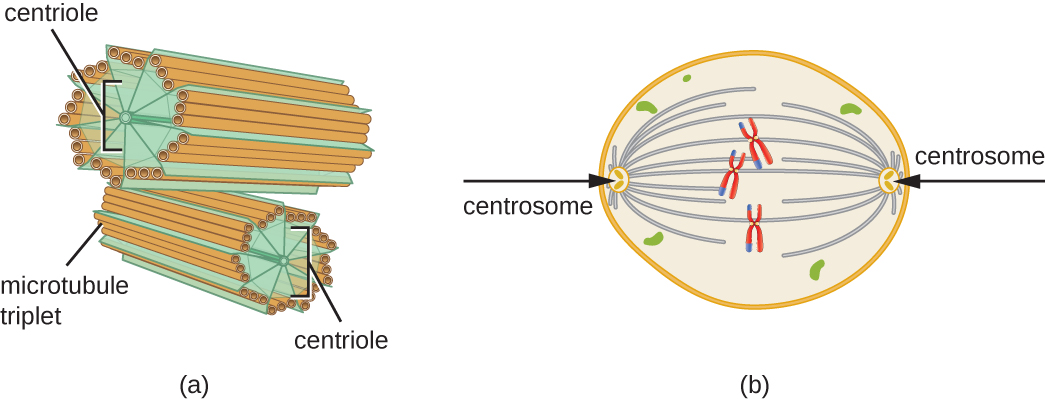
תרגיל \(\PageIndex{3}\)
השווה והשוואה בין שלושת סוגי המבנים הציטו-שלד המתוארים בסעיף זה.
מיטוכונדריה
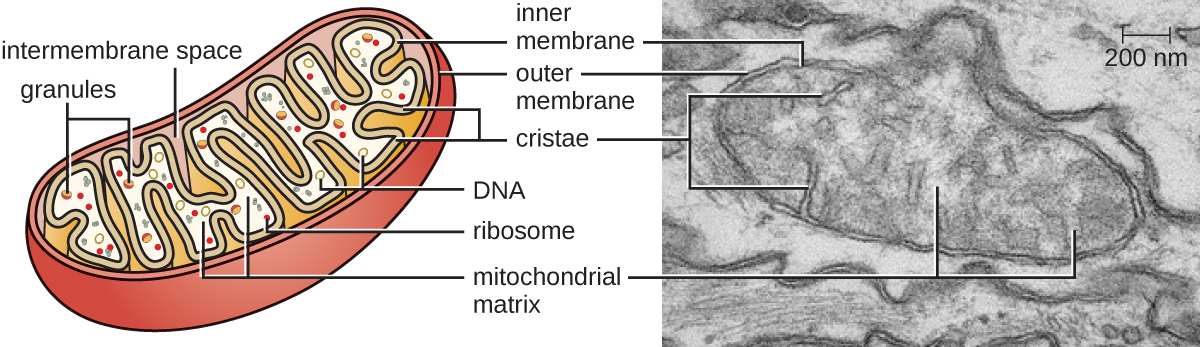
האברונים הגדולים והמורכבים שבהם מתרחשת נשימה תאית אירובית בתאים אוקריוטיים נקראים מיטוכונדריה (איור\(\PageIndex{15}\)). המונח "מיטוכונדריון" נטבע לראשונה על ידי המיקרוביולוג הגרמני קרל בנדה בשנת 1898 ומאוחר יותר נקשר לתהליך הנשימה על ידי אוטו ורבורג בשנת 1913. מדענים במהלך שנות השישים גילו שלמיטוכונדריה יש גנום משלהם וריבוזומים 70S. הגנום המיטוכונדריאלי נמצא כחיידקי, כאשר הוא רצף בשנת 1976. ממצאים אלה תמכו בסופו של דבר בתיאוריה האנדוסימביוטית שהציעה לין מרגוליס, הקובעת כי המיטוכונדריה נוצרה במקור באמצעות אירוע אנדוסימביוטי שבו חיידק המסוגל לנשימה תאית אירובית נקלט על ידי פגוציטוזיס לתא מארח ונשאר כמרכיב תוך תאי בר-קיימא.
לכל מיטוכונדריון יש שתי ממברנות שומנים. הממברנה החיצונית היא שריד למבני הממברנה של התא המארח המקורי. הממברנה הפנימית נגזרה מממברנת הפלזמה החיידקית. שרשרת הובלת האלקטרונים לנשימה אירובית משתמשת בחלבונים אינטגרליים המוטבעים בקרום הפנימי. המטריצה המיטוכונדריאלית, המתאימה למיקום הציטופלזמה של החיידק המקורי, היא המיקום הנוכחי של אנזימים מטבוליים רבים. הוא מכיל גם DNA מיטוכונדריאלי וריבוזומים 70S. פלישות של הממברנה הפנימית, הנקראות cristae, התפתחו כדי להגדיל את שטח הפנים למיקום התגובות הביוכימיות. דפוסי הקיפול של הקריסטות שונים בין סוגים שונים של תאים אוקריוטיים ומשמשים להבחנה בין אורגניזמים אוקריוטיים שונים זה מזה.
כלורופלסטים
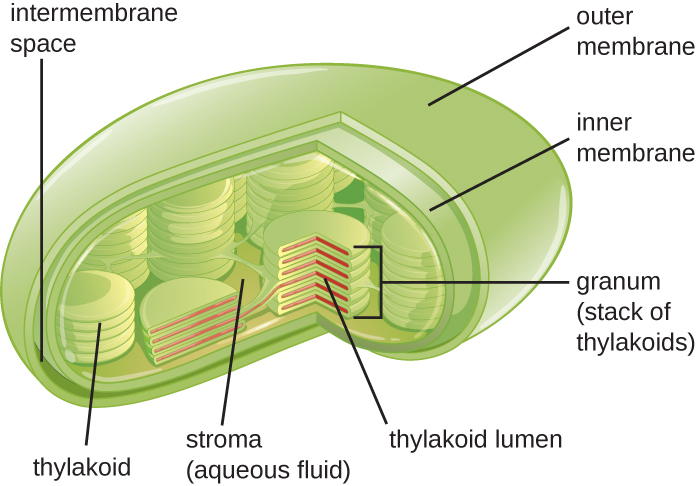
תאי צמחים ותאי אצות מכילים כלורופלסטים, האברונים שבהם מתרחשת פוטוסינתזה (איור). \(\PageIndex{16}\) לכל הכלורופלסטים יש לפחות שלוש מערכות ממברנה: הממברנה החיצונית, הממברנה הפנימית ומערכת הממברנה התילקואידית. בתוך הממברנות החיצוניות והפנימיות נמצאת סטרומה של כלורופלסט, נוזל דמוי ג'ל המרכיב חלק ניכר מנפח הכלורופלסט, ובו צפה מערכת התילקואיד. מערכת התילקואיד היא אוסף דינמי ביותר של שקי קרום מקופלים. זה המקום בו נמצא הפיגמנט הפוטוסינתטי הירוק כלורופיל ותגובות האור של הפוטוסינתזה מתרחשות. ברוב הכלורופלסטים הצמחיים, התילקואידים מסודרים בערימות הנקראות גרנה (ביחיד: גרנום), בעוד שבחלק מהכלורופלסטים של אצות, התילקואידים צפים חופשיים.
אברונים אחרים הדומים למיטוכונדריה התעוררו בסוגים אחרים של אוקריוטים, אך תפקידיהם שונים. הידרוגנוזומים נמצאים בכמה אוקריוטים אנאירוביים ומשמשים כמיקום של ייצור מימן אנאירובי. הידרוגנוזומים בדרך כלל חסרים DNA וריבוזומים משלהם. קינטופלסטים הם וריאציה של המיטוכונדריה המצויה בכמה פתוגנים אוקריוטיים. באורגניזמים אלה, לכל תא יש מיטוכונדריון יחיד, ארוך ומסועף שבו נמצא DNA קינטופלסט, המאורגן כחלקים עגולים מרובים של DNA, מרוכז בקוטב אחד של התא.
אברונים הקשורים למיטוכונדריה בטפילים פרוטוזואנים
פרוטוזואנים רבים, כולל מספר טפילים פרוטוזואנים הגורמים לזיהומים בבני אדם, ניתנים לזיהוי על ידי הופעתם יוצאת הדופן. מאפייני הבחנה עשויים לכלול מורפולוגיות תאים מורכבות, נוכחות של אברונים ייחודיים או היעדר אברונים נפוצים. הטפילים הפרוטוזואנים ג'יארדיה למבליה ו Trichomonas vaginalis הן שתי דוגמאות.
G. lamblia, גורם שכיח לשלשול בבני אדם ובעלי חיים רבים אחרים, הוא טפיל אנאירובי בעל שני גרעינים וכמה דגלים. מנגנון הגולגי והרטיקולום האנדופלזמי שלו מצטמצמים מאוד, והוא חסר מיטוכונדריה לחלוטין. עם זאת, יש לו אברונים המכונים מיטוזומים, אברונים הקשורים לממברנה כפולה שנראים כמיטוכונדריה מופחתים מאוד. זה גרם למדענים להאמין שאבות אבותיו של ג 'למבליה החזיקו פעם במיטוכונדריה שהתפתחו והפכו למיטוזומים. T. vaginalis, הגורם לזיהום המועבר במגע מיני דלקת הנרתיק, הוא טפיל פרוטוזואני נוסף חסר מיטוכונדריה קונבנציונאלית. במקום זאת, יש לו הידרוגנוזומים, אברונים הקשורים למיטוכונדריה, כפול-ממברנה, המייצרים מימן מולקולרי המשמש בחילוף החומרים התאי. מדענים מאמינים כי הידרוגנוזומים, כמו מיטוזומים, התפתחו גם הם מהמיטוכונדריה. 5
ממברנה פלזמה
קרום הפלזמה של תאים אוקריוטיים דומה במבנהו לקרום הפלזמה הפרוקריוטית בכך שהוא מורכב בעיקר מפוספוליפידים היוצרים דו שכבה עם חלבונים היקפיים ואינטגרליים מוטבעים (איור). \(\PageIndex{17}\) רכיבי הממברנה הללו נעים בתוך מישור הממברנה על פי מודל הפסיפס הנוזלי. עם זאת, בניגוד לקרום הפרוקריוטי, ממברנות אוקריוטיות מכילות סטרולים, כולל כולסטרול, המשנים את נזילות הממברנה. בנוסף, תאים אוקריוטיים רבים מכילים כמה שומנים מיוחדים, כולל ספינגוליפידים, הנחשבים למלא תפקיד בשמירה על יציבות הממברנה וכן מעורבים במסלולי העברת אותות ותקשורת בין תא לתא.

מנגנוני הובלת ממברנה
תהליכי דיפוזיה פשוטה, דיפוזיה קלה והובלה פעילה משמשים הן בתאים אוקריוטיים והן בתאים פרוקריוטים. עם זאת, לתאים אוקריוטיים יש גם את היכולת הייחודית לבצע סוגים שונים של אנדוציטוזיס, ספיגת חומר באמצעות פלישת קרום פלזמה ויצירת ואקוול/שלפוחית (איור). \(\PageIndex{18}\) סוג של אנדוציטוזיס הכולל בליעת חלקיקים גדולים באמצעות פלישת קרום נקרא פגוציטוזיס, שפירושו "אכילת תאים". בפגוציטוזיס, חלקיקים (או תאים אחרים) סגורים בכיס בתוך הממברנה, אשר לאחר מכן צובט מהקרום ליצירת ואקום המקיף לחלוטין את החלקיק. סוג אחר של אנדוציטוזיס נקרא פינוציטוזיס, שפירושו "שתיית תאים". בפינוציטוזיס, חומרים ונוזלים קטנים ומומסים נלקחים לתא דרך שלפוחיות קטנות. פטריות ספרופיטיות, למשל, משיגות את חומרי המזון שלהן מחומר מת ומתפורר בעיקר באמצעות פינוציטוזיס.
אנדוציטוזה בתיווך קולטן היא סוג של אנדוציטוזה שיוזמת על ידי מולקולות ספציפיות הנקראות ליגנדים כאשר הן נקשרות לקולטני פני התא על הממברנה. אנדוציטוזיס בתיווך קולטן הוא המנגנון בו משתמשים הורמונים שמקורם בפפטיד ובאמין כדי להיכנס לתאים ומשמש גם וירוסים וחיידקים שונים לכניסה לתאי המארח.
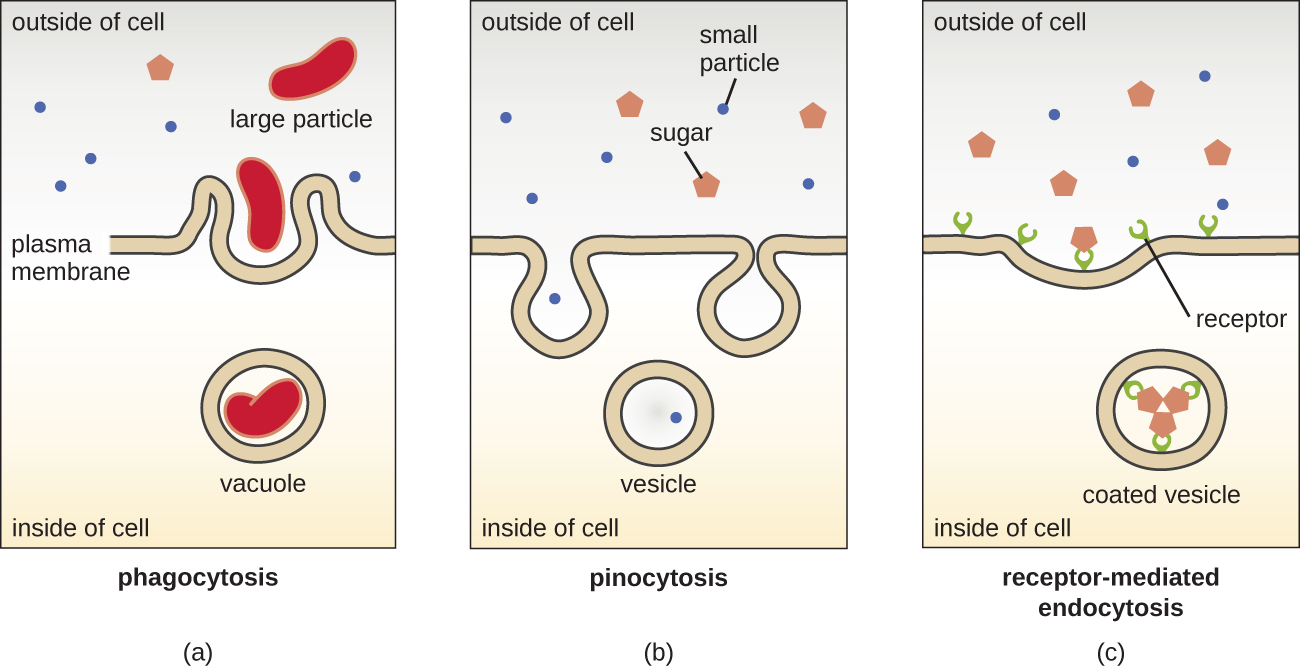
התהליך שבו שלפוחיות הפרשה משחררות את תוכנן לחלק החיצוני של התא נקרא אקסוציטוזיס. שלפוחיות נעות לכיוון קרום הפלזמה ואז מתמזגות עם הממברנה, ומוציאות את תוכנן מהתא. אקסוציטוזיס משמשת תאים להסרת מוצרי פסולת ועשויה לשמש גם לשחרור אותות כימיים שיכולים להיקלט על ידי תאים אחרים.
קיר תא
בנוסף לקרום פלזמה, לחלק מהתאים האוקריוטיים יש דופן תא. לתאים של פטריות, אצות, צמחים ואפילו כמה פרוטיסטים יש קירות תאים. בהתאם לסוג התא האוקריוטי, קירות התא יכולים להיות עשויים ממגוון רחב של חומרים, כולל תאית (פטריות וצמחים); סיליקה ביוגנית, סידן פחמתי, אגר וקרגינן (פרוטיסטים ואצות); או כיטין (פטריות). באופן כללי, כל דפנות התא מספקות יציבות מבנית לתא והגנה מפני מתחים סביבתיים כגון התייבשות, שינויים בלחץ האוסמוטי ופגיעה טראומטית. 6
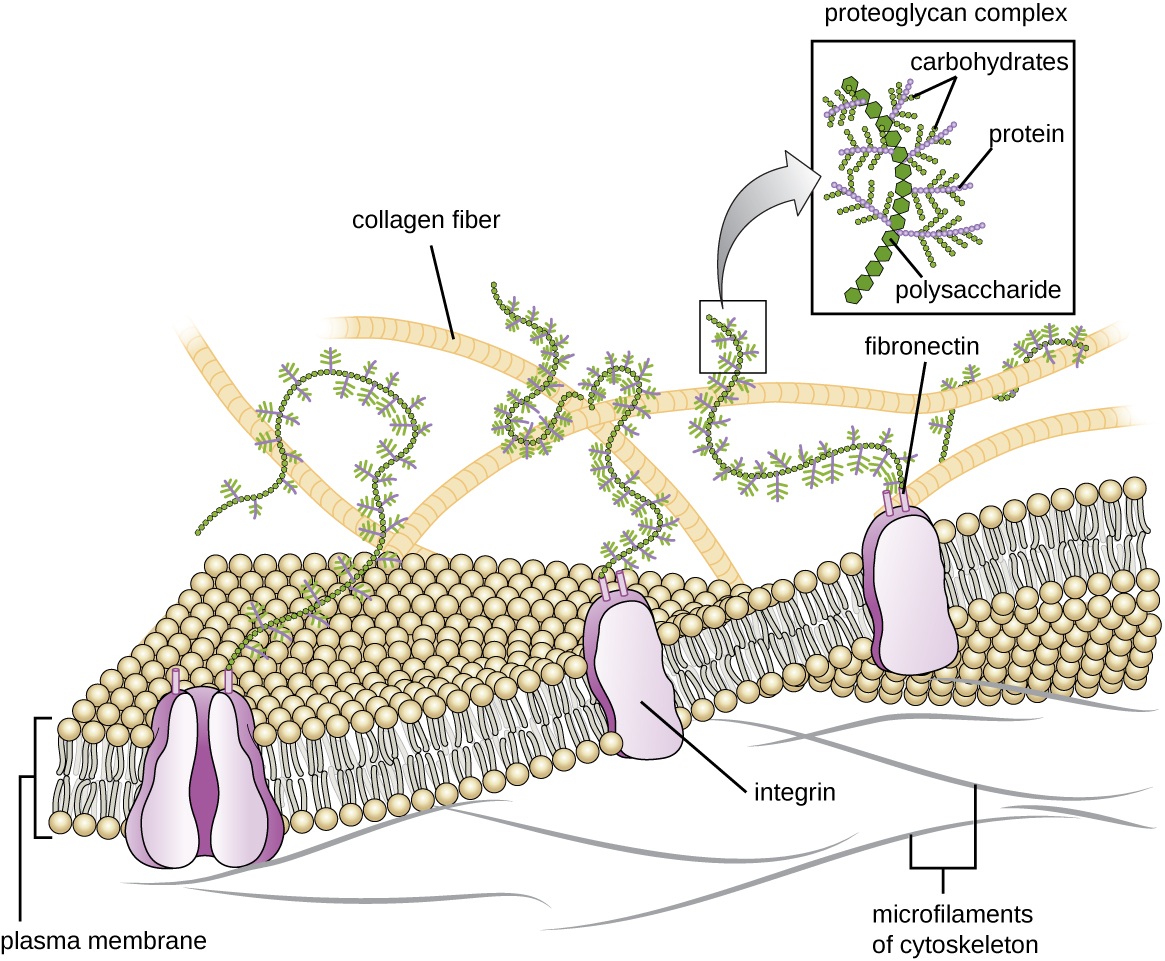
מטריקס חוץ תאי
לתאים של בעלי חיים ולחלק מהפרוטוזואנים אין קירות תאים שיעזרו לשמור על הצורה ולספק יציבות מבנית. במקום זאת, סוגים אלה של תאים אוקריוטיים מייצרים מטריצה חוץ -תאית למטרה זו. הם מפרישים מסה דביקה של פחמימות וחלבונים לחללים שבין תאים סמוכים (איור\(\PageIndex{19}\)). חלק ממרכיבי החלבון מתאספים לקרום מרתף שאליו נצמדים רכיבי המטריצה החוץ-תאית הנותרים. פרוטוגליקנים יוצרים בדרך כלל את המסה המגושמת של המטריצה החוץ -תאית בעוד שחלבונים סיביים, כמו קולגן, מספקים כוח. גם הפרוטוגליקנים וגם הקולגן מחוברים לחלבוני פיברונקטין, אשר בתורם מחוברים לחלבוני אינטגרין. חלבוני אינטגרין אלה מקיימים אינטראקציה עם חלבונים טרנסממברניים בממברנות הפלזמה של תאים אוקריוטיים חסרי דפנות תאים.
בתאי בעלי חיים, המטריצה החוץ -תאית מאפשרת לתאים בתוך הרקמות לעמוד בלחצים חיצוניים ומעבירה אותות מבחוץ התא פנימה. כמות המטריצה החוץ -תאית נרחבת למדי בסוגים שונים של רקמות חיבור, ושונות במטריצה החוץ -תאית יכולות לתת לסוגים שונים של רקמות את תכונותיהם המובהקות. בנוסף, המטריצה החוץ-תאית של תא מארח היא לעתים קרובות האתר שבו פתוגנים מיקרוביאליים מתחברים כדי לבסס זיהום. לדוגמה, Streptococcus pyogenes, החיידק הגורם לדלקת גרון וזיהומים שונים אחרים, נקשר לפיברונקטין במטריצה החוץ תאית של התאים המצפים את הלוע התחתון (האזור העליון של הגרון).
פלאגלה וסיליה
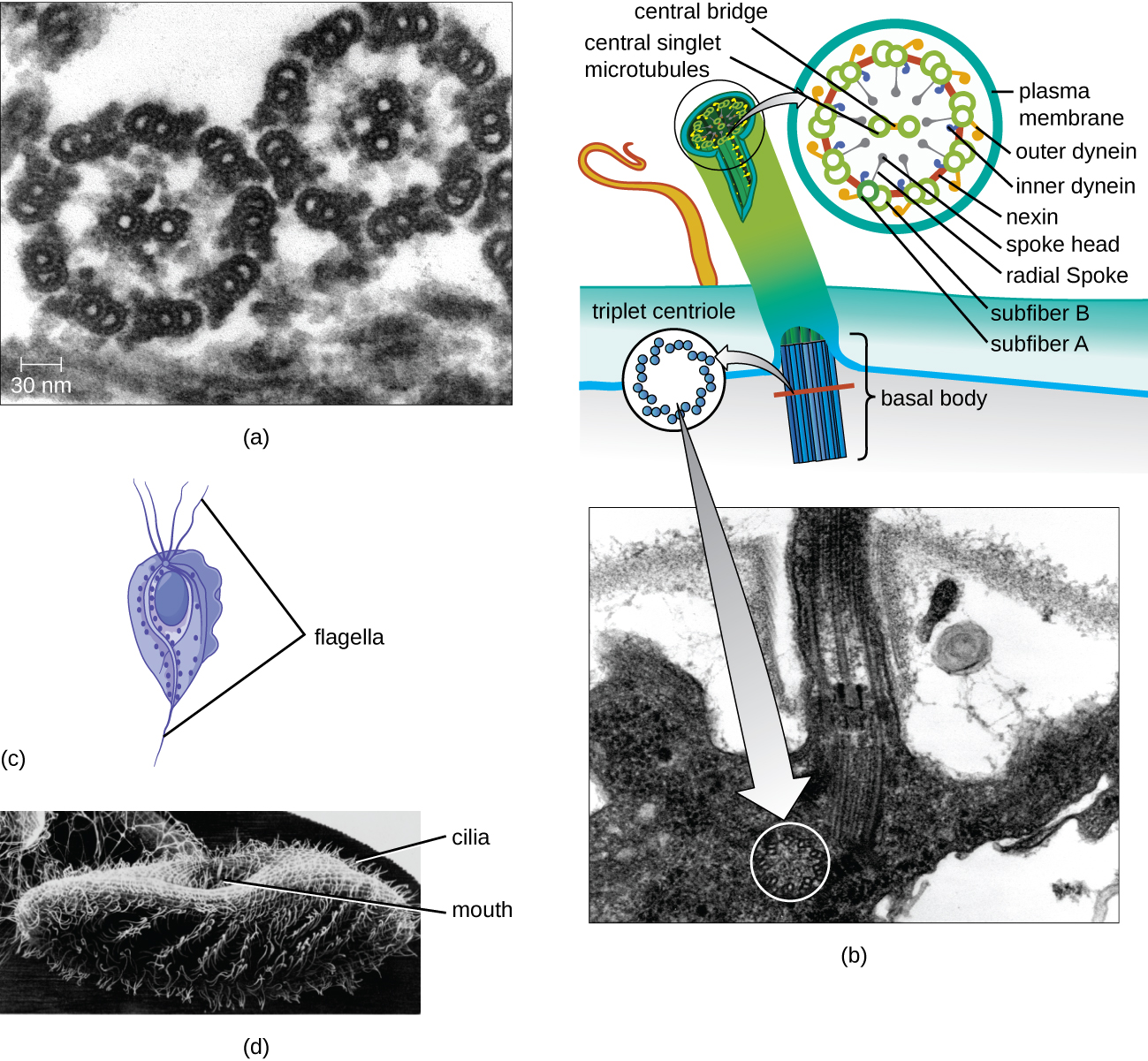
חלק מהתאים האיקריוטים משתמשים בדגלים לתנועה; עם זאת, דגלים אוקריוטיים נבדלים מבחינה מבנית מאלו המצויים בתאים פרוקריוטים. בעוד שהפלגלום הפרוקריוטי הוא מבנה נוקשה ומסתובב, דגל אוקריוטי דומה יותר לשוט גמיש המורכב מתשעה זוגות מקבילים של מיקרוטובולים המקיפים זוג מרכזי של מיקרו-צינורות. הסדר זה מכונה מערך 9+2 (איור\(\PageIndex{20}\)). המיקרוטובולים המקבילים משתמשים בחלבונים מוטוריים של דינין כדי לנוע יחסית זה לזה, מה שגורם לדגל להתכופף.
Cilia (יחיד: cilium) הם מבנה חיצוני דומה המצוי בכמה תאים אוקריוטיים. ייחודיות לאאוקריוטים, הריסים קצרים יותר מהדגלים ולעתים קרובות מכסים את כל פני התא; עם זאת, הם דומים מבחינה מבנית ל- flagella (מערך 9+2 של מיקרו-צינורות) ומשתמשים באותו מנגנון לתנועה. מבנה הנקרא גוף בסיסי נמצא בבסיס כל ציליום ופלגלום. הגוף הבסיסי, המחבר את הציליום או הדגל לתא, מורכב ממערך של מיקרו-צינורות משולשים הדומים לזה של צנטריול אך מוטבעים בקרום הפלזמה. בגלל אורכם הקצר יותר, הריסים משתמשים בתנועה מהירה, גמישה ומנופפת. בנוסף לתנועתיות, לציליות עשויות להיות פונקציות אחרות כגון טאטוא חלקיקים מעבר לתאים או לתאים. לדוגמה, פרוטוזואים ריסים משתמשים בטאטוא של ריסים כדי להעביר חלקיקי מזון לחלקי הפה שלהם, ותאים ריסים בדרכי הנשימה של היונקים פועמים בסינכרון כדי לטאטא ריר ופסולת מעלה והחוצה מהריאות (איור). \(\PageIndex{20}\)
תרגיל \(\PageIndex{4}\)
- הסבר כיצד המעטפת התאית של תאים אוקריוטים משתווה לזו של תאים פרוקריוטים.
- הסבר את ההבדל בין דגלים אוקריוטיים לפרוקריוטים.
מיקוד קליני: רזולוציה
מכיוון שאמוקסיצילין לא פתר את המקרה של ברברה של דלקת ריאות, הרשות הפלסטינית רושמת אנטיביוטיקה נוספת, אזיתרומיצין, המכוונת לריבוזומים חיידקיים ולא לפפטידוגליקן. לאחר נטילת האזיתרומיצין לפי ההוראות, הסימפטומים של ברברה נפתרים והיא סוף סוף מתחילה להרגיש כמו עצמה שוב. בהנחה שלא הייתה מעורבת עמידות לתרופות לאמוקסיצילין, ובהתחשב ביעילות של אזיתרומיצין, הסוכן הסיבתי לדלקת ריאות של ברברה הוא ככל הנראה Mycoplasma pneumoniae. למרות שחיידק זה הוא תא פרוקריוטי, הוא אינו מעוכב על ידי אמוקסיצילין מכיוון שאין לו דופן תא ולכן אינו מייצר פפטידוגליקן.
מושגי מפתח וסיכום
- תאים אוקריוטיים מוגדרים על ידי נוכחות של גרעין המכיל את גנום ה- DNA וקשורים בממברנה גרעינית (או מעטפת גרעינית) המורכבת משתי שכבות שומנים המווסתות את הובלת החומרים אל הגרעין ומחוצה לו דרך נקבוביות גרעיניות.
- מורפולוגיות תאים אוקריוטיות משתנות מאוד ועשויות להישמר על ידי מבנים שונים, כולל שלד הציטוס, קרום התא ו/או דופן התא.
- הגרעין, הממוקם בגרעין התאים האוקריוטיים, הוא אתר הסינתזה הריבוזומלית והשלבים הראשונים של הרכבת הריבוזומים.
- תאים אוקריוטיים מכילים ריבוזומים של 80S ברטיקולום האנדופלזמי המחוספס (ריבוזומים מוגבלים בממברנה) ובציטופלזמה (ריבוזומים חופשיים). הם מכילים ריבוזומים משנות ה -70 במיטוכונדריה ובכלורופלסטים.
- תאים אוקריוטיים פיתחו מערכת אנדוממברנית, המכילה אברונים הקשורים לממברנה המעורבים בהובלה. אלה כוללים שלפוחיות, הרטיקולום האנדופלזמי ומנגנון גולגי.
- הרטיקולום האנדופלזמי החלק ממלא תפקיד בביוסינתזה של שומנים, מטבוליזם של פחמימות וניקוי רעלים של תרכובות רעילות. הרשת האנדופלזמית המחוספסת מכילה ריבוזומים 80S הקשורים לממברנה המסנתזים חלבונים המיועדים לקרום התא
- מנגנון גולגי מעבד חלבונים ושומנים, בדרך כלל באמצעות הוספת מולקולות סוכר, ייצור גליקופרוטאינים או גליקוליפידים, רכיבים של קרום הפלזמה המשמשים בתקשורת בין תא לתא.
- ליזוזומים מכילים אנזימי עיכול המפרקים חלקיקים קטנים הנבלעים על ידי אנדוציטוזיס, חלקיקים גדולים או תאים הנבלעים על ידי פגוציטוזיס ורכיבים תאיים פגומים.
- שלד הציטוס, המורכב ממיקרופילמנטים, חוטי ביניים ומיקרוטובולים, מספק תמיכה מבנית בתאים אוקריוטיים ומשמש כרשת להובלת חומרים תוך תאיים.
- צנטרוזומים הם מרכזים המארגנים מיקרו-צינורות החשובים ביצירת הציר המיטוטי במיטוזה.
- מיטוכונדריה הן אתר הנשימה התאית. יש להם שני ממברנות: קרום חיצוני וקרום פנימי עם cristae. המטריצה המיטוכונדריאלית, בתוך הממברנה הפנימית, מכילה את ה- DNA המיטוכונדריאלי, ריבוזומים 70S ואנזימים מטבוליים.
- קרום הפלזמה של תאים אוקריוטיים דומה מבחינה מבנית לזה שנמצא בתאים פרוקריוטים, ורכיבי הממברנה נעים על פי מודל הפסיפס הנוזלי. עם זאת, ממברנות אוקריוטיות מכילות סטרולים, המשנים את נזילות הממברנה, כמו גם גליקופרוטאינים וגליקוליפידים, המסייעים לתא לזהות תאים אחרים וחלקיקים זיהומיים.
- בנוסף להובלה פעילה והובלה פסיבית, ממברנות תאים אוקריוטיות יכולות לקחת חומר לתא באמצעות אנדוציטוזיס, או לגרש חומר מהתא באמצעות אקסוציטוזיס.
- לתאים של פטריות, אצות, צמחים וכמה פרוטיסטים יש דופן תא, בעוד שלתאים של בעלי חיים ולחלק מהפרוטוזואנים יש מטריצה חוץ-תאית דביקה המספקת תמיכה מבנית ומתווכת איתות תאי.
- דגלים אוקריוטיים נבדלים מבחינה מבנית מהדגלים הפרוקריוטים אך משרתים מטרה דומה (תנועה). Cilia דומים מבחינה מבנית לדגלים אוקריוטיים, אך קצרים יותר; הם עשויים לשמש לתנועה, האכלה או תנועה של חלקיקים חוץ-תאיים.
הערות שוליים
- א. א. ברנהיל, מ. טי. ברואר, ס.א. קרלסון. "השפעות שליליות של חומרים אנטי-מיקרוביאליים באמצעות עיכוב צפוי או אידיוסינקרטי של רכיבים מיטוכונדריאליים מארחים." תרופות אנטי-מיקרוביאליות וכימותרפיה 56 מס '8 (2012) :4046—4051.
- 2 פוקס E, קליבלנד DW. "פיגומים מבניים של חוטי ביניים בבריאות ומחלות." מדע 279 מס '5350 (1998) :514—519.
- 3 אי פוקס, DW קליבלנד. "פיגומים מבניים של חוטי ביניים בבריאות ומחלות." מדע 279 מס '5350 (1998) :514—519.
- 4 אי פוקס, DW קליבלנד. "פיגומים מבניים של חוטי ביניים בבריאות ומחלות." מדע 279 מס '5350 (1998) :514—519.
- 5 נ 'ירלט, JHP הקשטיין. "הידרוגנוזומים: אברון אחד, מקורות מרובים." מדע ביולוגי 55 מס '8 (2005) :657—658.
- 6 מ 'דודזיק. "פרוטיסטים". אופנסטקס CNX. 27 בנובמבר 2013. http://cnx.org/contents/f7048bb6-e46...ef291cf7049c@1