
10.1: Kutumia Microbiology kugundua Siri za Maisha
- Page ID
- 174628

\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\( \newcommand{\dsum}{\displaystyle\sum\limits} \)
\( \newcommand{\dint}{\displaystyle\int\limits} \)
\( \newcommand{\dlim}{\displaystyle\lim\limits} \)
\( \newcommand{\id}{\mathrm{id}}\) \( \newcommand{\Span}{\mathrm{span}}\)
( \newcommand{\kernel}{\mathrm{null}\,}\) \( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\) \( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\) \( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\id}{\mathrm{id}}\)
\( \newcommand{\Span}{\mathrm{span}}\)
\( \newcommand{\kernel}{\mathrm{null}\,}\)
\( \newcommand{\range}{\mathrm{range}\,}\)
\( \newcommand{\RealPart}{\mathrm{Re}}\)
\( \newcommand{\ImaginaryPart}{\mathrm{Im}}\)
\( \newcommand{\Argument}{\mathrm{Arg}}\)
\( \newcommand{\norm}[1]{\| #1 \|}\)
\( \newcommand{\inner}[2]{\langle #1, #2 \rangle}\)
\( \newcommand{\Span}{\mathrm{span}}\) \( \newcommand{\AA}{\unicode[.8,0]{x212B}}\)
\( \newcommand{\vectorA}[1]{\vec{#1}} % arrow\)
\( \newcommand{\vectorAt}[1]{\vec{\text{#1}}} % arrow\)
\( \newcommand{\vectorB}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\( \newcommand{\vectorC}[1]{\textbf{#1}} \)
\( \newcommand{\vectorD}[1]{\overrightarrow{#1}} \)
\( \newcommand{\vectorDt}[1]{\overrightarrow{\text{#1}}} \)
\( \newcommand{\vectE}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash{\mathbf {#1}}}} \)
\( \newcommand{\vecs}[1]{\overset { \scriptstyle \rightharpoonup} {\mathbf{#1}} } \)
\(\newcommand{\longvect}{\overrightarrow}\)
\( \newcommand{\vecd}[1]{\overset{-\!-\!\rightharpoonup}{\vphantom{a}\smash {#1}}} \)
\(\newcommand{\avec}{\mathbf a}\) \(\newcommand{\bvec}{\mathbf b}\) \(\newcommand{\cvec}{\mathbf c}\) \(\newcommand{\dvec}{\mathbf d}\) \(\newcommand{\dtil}{\widetilde{\mathbf d}}\) \(\newcommand{\evec}{\mathbf e}\) \(\newcommand{\fvec}{\mathbf f}\) \(\newcommand{\nvec}{\mathbf n}\) \(\newcommand{\pvec}{\mathbf p}\) \(\newcommand{\qvec}{\mathbf q}\) \(\newcommand{\svec}{\mathbf s}\) \(\newcommand{\tvec}{\mathbf t}\) \(\newcommand{\uvec}{\mathbf u}\) \(\newcommand{\vvec}{\mathbf v}\) \(\newcommand{\wvec}{\mathbf w}\) \(\newcommand{\xvec}{\mathbf x}\) \(\newcommand{\yvec}{\mathbf y}\) \(\newcommand{\zvec}{\mathbf z}\) \(\newcommand{\rvec}{\mathbf r}\) \(\newcommand{\mvec}{\mathbf m}\) \(\newcommand{\zerovec}{\mathbf 0}\) \(\newcommand{\onevec}{\mathbf 1}\) \(\newcommand{\real}{\mathbb R}\) \(\newcommand{\twovec}[2]{\left[\begin{array}{r}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\ctwovec}[2]{\left[\begin{array}{c}#1 \\ #2 \end{array}\right]}\) \(\newcommand{\threevec}[3]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\cthreevec}[3]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \end{array}\right]}\) \(\newcommand{\fourvec}[4]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\cfourvec}[4]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \end{array}\right]}\) \(\newcommand{\fivevec}[5]{\left[\begin{array}{r}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\cfivevec}[5]{\left[\begin{array}{c}#1 \\ #2 \\ #3 \\ #4 \\ #5 \\ \end{array}\right]}\) \(\newcommand{\mattwo}[4]{\left[\begin{array}{rr}#1 \amp #2 \\ #3 \amp #4 \\ \end{array}\right]}\) \(\newcommand{\laspan}[1]{\text{Span}\{#1\}}\) \(\newcommand{\bcal}{\cal B}\) \(\newcommand{\ccal}{\cal C}\) \(\newcommand{\scal}{\cal S}\) \(\newcommand{\wcal}{\cal W}\) \(\newcommand{\ecal}{\cal E}\) \(\newcommand{\coords}[2]{\left\{#1\right\}_{#2}}\) \(\newcommand{\gray}[1]{\color{gray}{#1}}\) \(\newcommand{\lgray}[1]{\color{lightgray}{#1}}\) \(\newcommand{\rank}{\operatorname{rank}}\) \(\newcommand{\row}{\text{Row}}\) \(\newcommand{\col}{\text{Col}}\) \(\renewcommand{\row}{\text{Row}}\) \(\newcommand{\nul}{\text{Nul}}\) \(\newcommand{\var}{\text{Var}}\) \(\newcommand{\corr}{\text{corr}}\) \(\newcommand{\len}[1]{\left|#1\right|}\) \(\newcommand{\bbar}{\overline{\bvec}}\) \(\newcommand{\bhat}{\widehat{\bvec}}\) \(\newcommand{\bperp}{\bvec^\perp}\) \(\newcommand{\xhat}{\widehat{\xvec}}\) \(\newcommand{\vhat}{\widehat{\vvec}}\) \(\newcommand{\uhat}{\widehat{\uvec}}\) \(\newcommand{\what}{\widehat{\wvec}}\) \(\newcommand{\Sighat}{\widehat{\Sigma}}\) \(\newcommand{\lt}{<}\) \(\newcommand{\gt}{>}\) \(\newcommand{\amp}{&}\) \(\definecolor{fillinmathshade}{gray}{0.9}\)Malengo ya kujifunza
- Eleza ugunduzi wa asidi nucleic na nucleotides
- Eleza majaribio ya kihistoria ambayo imesababisha Tabia ya DNA
- Eleza jinsi microbiolojia na microorganisms zimetumika kugundua biokemia ya jeni
- Eleza jinsi wanasayansi walivyoanzisha uhusiano kati ya DNA na urithi
Mwelekeo wa kliniki: Sehemu ya 1
Alex ni mwanafunzi wa chuo mwenye umri wa miaka 22 ambaye alikwenda likizo huko Puerta Vallarta, Mexico, kwa mapumziko ya spring. Kwa bahati mbaya, siku mbili baada ya kuruka nyumbani kwa Ohio, alianza kupata tumbo la tumbo na kuhara kwa kina maji. Kwa sababu ya usumbufu wake, alitafuta matibabu katika hospitali kubwa ya Cincinnati karibu.
Zoezi\(\PageIndex{1}\)
Ni aina gani za maambukizi au hali nyingine zinaweza kuwajibika?
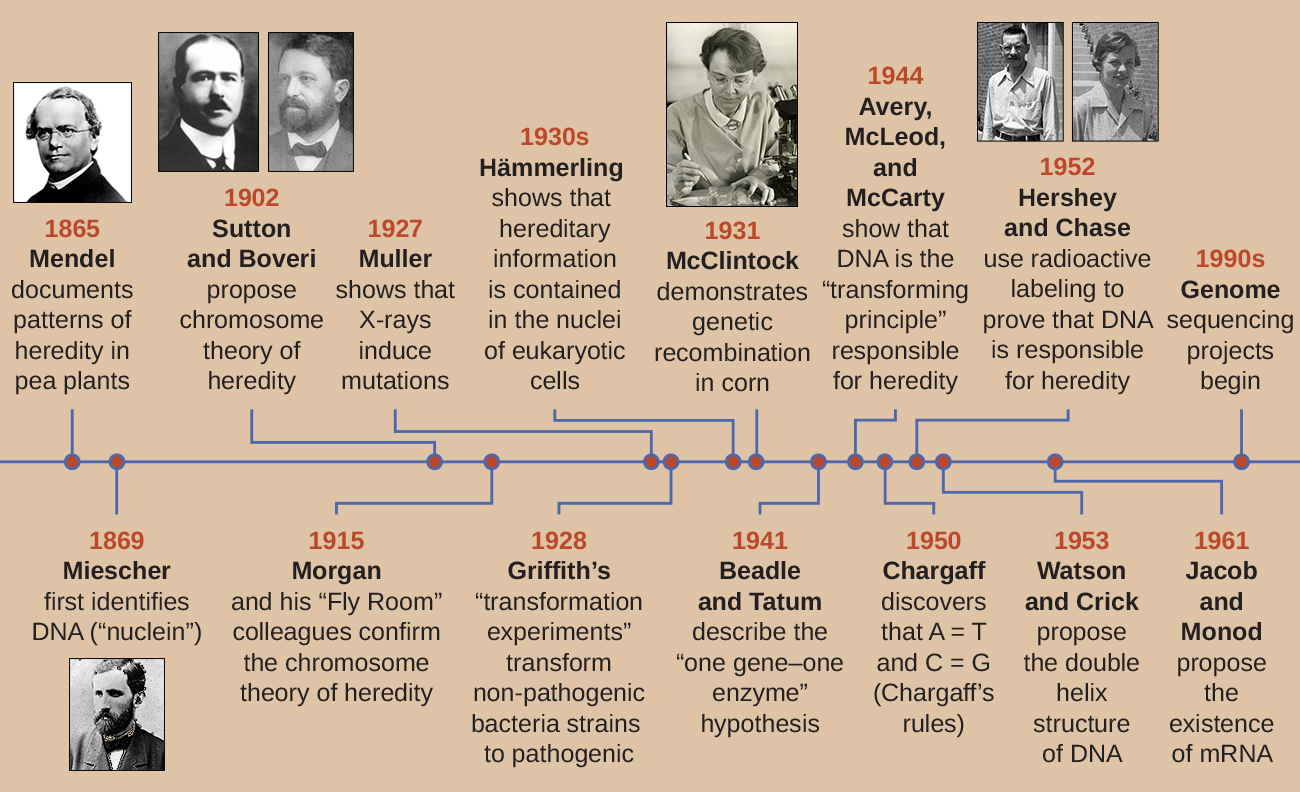
Kupitia mapema karne ya 20, DNA haijawahi kutambuliwa kama nyenzo za maumbile zinazohusika na urithi, kifungu cha sifa kutoka kizazi kimoja hadi kijacho. Kwa kweli, utafiti mwingi ulifukuzwa hadi katikati ya karne ya 20. Jumuiya ya kisayansi iliamini, kwa usahihi, kwamba mchakato wa urithi ulihusisha kuchanganya sifa za wazazi ambazo zilizalisha kuonekana kwa kimwili kwa watoto; mchakato huu wa nadharia ulionekana kuwa sahihi kwa sababu ya kile tunachokijua sasa kama tofauti inayoendelea, ambayo inatokana na hatua ya jeni nyingi kuamua tabia fulani, kama urefu wa binadamu. Watoto wanaonekana kuwa “mchanganyiko” wa sifa za wazazi wao tunapoangalia sifa zinazoonyesha tofauti inayoendelea. Nadharia ya kuchanganya ya urithi ilisema kuwa sifa za wazazi wa awali zilipotea au kufyonzwa na kuchanganya katika watoto, lakini sasa tunajua kwamba hii sio kesi.
Mistari miwili tofauti ya utafiti, iliyoanza katikati hadi miaka ya 1800 marehemu, hatimaye imesababisha ugunduzi na Tabia ya DNA na misingi ya jenetiki, sayansi ya urithi. Mistari hii ya utafiti ilianza kuungana katika miaka ya 1920, na utafiti kwa kutumia mifumo microbial hatimaye ilisababisha michango muhimu ya elucidating msingi Masi ya jenetiki.
Ugunduzi na Tabia ya DNA
Uelewa wa kisasa wa DNA umebadilika kutokana na ugunduzi wa asidi ya nucleic hadi maendeleo ya mfano wa mbili-helix. Katika miaka ya 1860, Friedrich Miescher (1844—1895), daktari kwa taaluma, alikuwa mtu wa kwanza kutenganisha kemikali tajiri ya fosforasi kutoka leukocytes (seli nyeupe za damu) kutoka usaha kwenye bandeji zilizotumika kutoka kliniki ya upasuaji wa ndani. Alitaja kemikali hizi (ambazo hatimaye zingejulikana kama RNA na DNA) “nuclein” kwa sababu zilitengwa na viini vya seli. Mwanafunzi wake Richard Altmann (1852—1900) baadaye aliiita “asidi nucleic” miaka 20 baadaye alipogundua asili ya tindikali ya nuclein. Katika miongo miwili iliyopita ya karne ya 19, mwanabiolojia wa Ujerumani Albrecht Kossel (1853—1927) alitenga na sifa ya misingi mitano tofauti ya nucleotide inayojumuisha asidi ya nucleic. Hizi ni adenine, guanine, cytosine, thymine (katika DNA), na uracil (katika RNA). Kossell alipokea Tuzo ya Nobel katika Physiolojia au Tiba mwaka wa 1910 kwa kazi yake juu ya asidi ya nucleic na kwa kazi yake kubwa juu ya protini, ikiwa ni pamoja na ugunduzi wa histidine.
Misingi ya Genetics
Licha ya ugunduzi wa DNA mwishoni mwa miaka ya 1800, wanasayansi hawakufanya ushirikiano na urithi kwa miongo mingi zaidi. Ili kufanya uhusiano huu, wanasayansi, ikiwa ni pamoja na idadi ya microbiologists, walifanya majaribio mengi juu ya mimea, wanyama, na bakteria.
Mimea ya Pea ya Mendel
Wakati Miescher alikuwa akitenga na kugundua DNA katika miaka ya 1860, mtawa na mtaalamu wa mimea wa Austria Johann Gregor Mendel (1822—1884) alikuwa akijaribu na mbaazi za bustani, akionyesha na kuandika mifumo ya msingi ya urithi, ambayo sasa inajulikana kama sheria za Mendel.
Mwaka 1856, Mendel alianza utafiti wake wa miaka kumi katika mifumo ya urithi. Alitumia pea ya bustani ya diploid, Pisum sativum, kama mfumo wake wa msingi wa mfano kwa sababu kwa kawaida hujitengeneza mbolea na ni inbred sana, huzalisha “kweli-kuzaliana” mistari ya mimea ya mbaa-mimea ambayo daima huzalisha watoto ambao huonekana kama mzazi. Kwa kujaribu mimea ya kweli ya kuzaliana, Mendel aliepuka kuonekana kwa sifa zisizotarajiwa katika watoto ambazo zinaweza kutokea ikiwa alitumia mimea ambayo haikuwa ya kuzaliana kweli. Mendel alifanya mahuluti, ambayo inahusisha kuunganisha watu wawili wa kweli wa kuzaliana (P kizazi) ambao wana sifa tofauti, na kuchunguza sifa za watoto wao (kizazi cha kwanza cha filial, F 1) pamoja na watoto wa kujitegemea wa kizazi cha F 1 (pili kizazi cha watoto, F 2) (Kielelezo\(\PageIndex{1}\)).
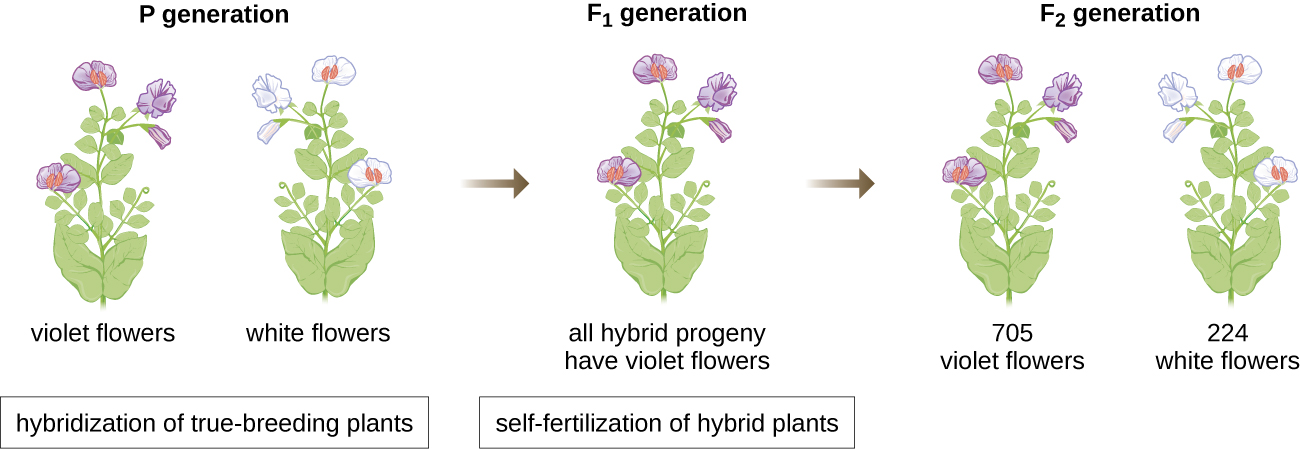
Mwaka 1865, Mendel aliwasilisha matokeo ya majaribio yake na karibu mimea ya pea 30,000 kwa jamii ya ndani ya historia ya asili. Alionyesha kuwa sifa zinaambukizwa kwa uaminifu kutoka kwa wazazi hadi watoto kwa kujitegemea sifa nyingine. Mwaka 1866, alichapisha kazi yake, “Majaribio katika Uchanganyiko wa Plant,” 1 katika Mahakama ya Shirika la Historia Asili la Brünn. Kazi ya Mendel ilikwenda karibu bila kutambuliwa na jumuiya ya kisayansi, ambayo iliamini, kwa usahihi, katika nadharia ya kuchanganya sifa katika tofauti inayoendelea.
Hakutambuliwa kwa michango yake ya ajabu ya kisayansi wakati wa maisha yake. Kwa kweli, haikuwa hadi 1900 kwamba kazi yake iligunduliwa tena, tena, na kuimarishwa na wanasayansi ukingoni mwa kugundua msingi wa chromosomal wa urithi.
Theory ya Chromosomal ya Urithi
Mendel alifanya majaribio yake muda mrefu kabla ya chromosomes kutazamwa chini ya darubini. Hata hivyo, pamoja na uboreshaji wa mbinu microscopic wakati wa miaka ya 1800 mwishoni mwa miaka ya 1800, wanabiolojia wa kiini wanaweza kudanganya na kutazama miundo ya subcellular na rangi na kuchunguza matendo yao wakati wa meiosis. Walikuwa na uwezo wa kuchunguza chromosomes replicating, condensing kutoka amorphous nyuklia molekuli katika tofauti X-umbo miili na kuhamia kwa tofauti miti ya mkononi. Uvumi kwamba kromosomu inaweza kuwa ufunguo wa kuelewa urithi ulisababisha wanasayansi kadhaa kuchunguza machapisho ya Mendel na kutathmini upya mfano wake katika suala la tabia ya kromosomu wakati wa mitosisi na meiosis.
Mwaka 1902, Theodor Boveri (1862—1915) aliona kuwa katika urchins za bahari, vipengele vya nyuklia (chromosomes) viliamua maendeleo sahihi ya kiinitete. Mwaka huo huo, Walter Sutton (1877—1916) aliona kujitenga kwa kromosomu katika seli za binti wakati wa meiosis. Kwa pamoja, uchunguzi huu ulisababisha maendeleo ya Nadharia ya Chromosomal ya Urithi, ambayo ilitambua chromosomes kama nyenzo za maumbile zinazohusika na urithi wa Mendelian.
Licha ya mahusiano ya kulazimisha kati ya tabia za kromosomu wakati wa meiosis na uchunguzi wa Mendel, Nadharia ya Chromosomal ya Urithi ilipendekezwa muda mrefu kabla ya kuwa na ushahidi wowote wa moja kwa moja kwamba sifa zilifanywa kwenye chromosomes. Thomas Hunt Morgan (1866—1945) na wenzake walitumia miaka kadhaa wakifanya misalaba na kuruka kwa matunda, Drosophila melanogaster. Walifanya uchunguzi wa kina wa microscopic ya chromosomes ya kuruka na kuunganisha uchunguzi huu na sifa za kuruka. Kazi yao ilitoa ushahidi wa kwanza wa majaribio ili kusaidia Nadharia ya Chromosomal ya Urithi katika miaka ya 1900 mapema. Mwaka wa 1915, Morgan na wenzake wa “Fly Room” walichapisha The Mechanism of Mendelian Heredity, ambayo ilitambua kromosomu kama miundo ya seli inayohusika na urithi. Kwa michango yake mingi muhimu katika jenetiki, Morgan alipokea Tuzo ya Nobel katika Fiziolojia au Tiba mwaka wa 1933.
Mwishoni mwa miaka ya 1920, Barbara McClintock (1902—1992) alitengeneza mbinu za kudanganya chromosomal ili kutazama na kutofautisha kati ya chromosomes tofauti za mahindi (mahindi). Katika miaka ya 1940 na 1950, alitambua tukio la kuvunjika kwenye kromosomu 9, ambalo aliita jina la dissociation locus (Ds). Ds inaweza kubadilisha msimamo ndani ya kromosomu. Pia alitambua locus activator (Ac). Ds kromosomu kuvunjika inaweza ulioamilishwa na kipengele Ac (enzyme transposase). Mwanzoni, uchunguzi wa McClintock wa jeni hizi za kuruka, ambazo sasa tunaziita transposons, hakukubaliwa na jumuiya ya kisayansi. Haikuwa mpaka miaka ya 1960 na baadaye kwamba transposons ziligunduliwa katika bacteriophages, bakteria, na Drosophila. Leo, tunajua kwamba transposons ni makundi ya simu ya DNA ambayo yanaweza kuhamia ndani ya genome ya kiumbe. Wanaweza kudhibiti usemi wa jeni, kujieleza kwa protini, na virulence (uwezo wa kusababisha ugonjwa).
Microbes na virusi katika Utafiti wa Maumbile
Wataalamu wa microbiologists pia wamekuwa na sehemu muhimu katika uelewa wetu wa maumbile. Viumbe vya majaribio kama vile mbaazi za bustani za Mendel, nzi za matunda ya Morgan, na mahindi ya McClintock yalikuwa tayari yametumiwa kwa mafanikio kusafisha njia kwa uelewa wa jenetiki. Hata hivyo, microbes na virusi zilikuwa (na bado ni) mifumo bora ya mfano kwa ajili ya utafiti wa jenetiki kwa sababu, tofauti na mbaazi, nzizi za matunda, na mahindi, zinaenea kwa urahisi zaidi katika maabara, huongezeka kwa msongamano mkubwa wa idadi ya watu kwa kiasi kidogo cha nafasi na kwa muda mfupi. Kwa kuongeza, kwa sababu ya unyenyekevu wao wa kimuundo, microbes na virusi vinatumiwa kwa urahisi zaidi.
Kwa bahati nzuri, licha ya tofauti kubwa katika ukubwa, muundo, mikakati ya uzazi, na sifa nyingine za kibaiolojia, kuna umoja wa biochemical kati ya viumbe vyote; wana kwa pamoja molekuli sawa za msingi zinazohusika na urithi na matumizi ya vifaa vya maumbile ili kuwapa seli tofauti zao sifa. Kwa maneno ya mwanasayansi wa Kifaransa Jacques Monod, “Nini kweli kwa E. koli pia ni kweli kwa tembo,” maana yake ni kwamba biokemia ya maisha imehifadhiwa katika mageuzi na inashirikiwa katika aina zote za maisha, kuanzia viumbe rahisi vya unicellular hadi viumbe vikubwa, tata. Uendelezaji huu wa biochemical hufanya viumbe vidogo bora vya kutumia kwa masomo ya maumbile.
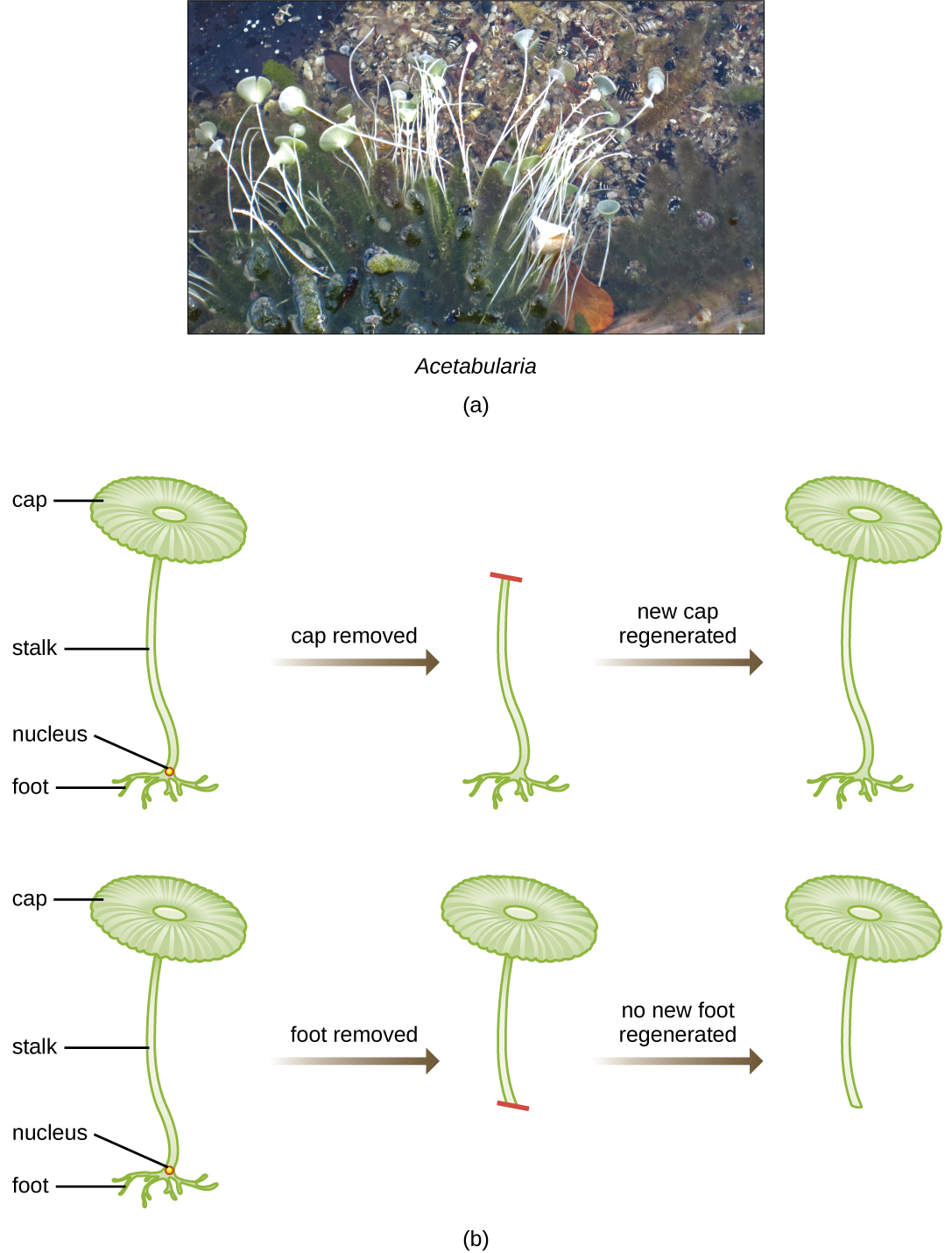
Katika seti ya wajanja ya majaribio katika miaka ya 1930 na 1940, mwanasayansi wa Ujerumani Joachim Hämmerling (1901—1980), akitumia Acetabularia ya alga moja ya seli kama mfano wa microbial, alibainisha kuwa habari za maumbile katika seli ya eukaryotic iko ndani ya kiini. Acetabularia spp. ni seli kubwa za algali ambazo zinakua kwa asymmetrically, na kutengeneza “mguu” iliyo na kiini, ambayo hutumiwa kwa attachment substrate; shina; na miundo kama cap-kama mwavuli ambayo inaweza kuonekana kwa urahisi kwa jicho la uchi. Katika seti ya awali ya majaribio, Hämmerling kuondolewa ama cap au mguu wa seli na aliona kama kofia mpya au miguu walikuwa upya (Kielelezo\(\PageIndex{2}\)). Aligundua kwamba wakati mguu wa seli hizi ulipoondolewa, miguu mpya haikukua; hata hivyo, wakati kofia ziliondolewa kwenye seli, kofia mpya zilirejeshwa. Hii ilipendekeza kuwa habari ya urithi ilikuwa iko katika mguu wa nucleus wa kila kiini.
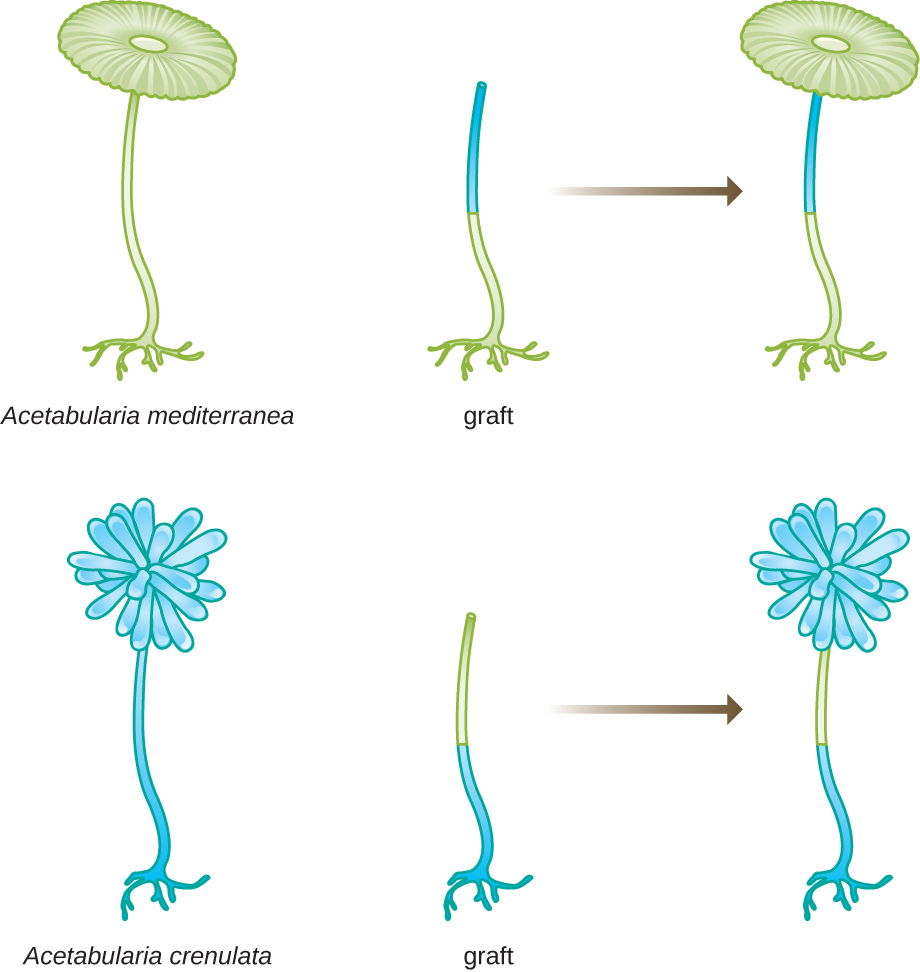
Katika seti nyingine ya majaribio, Hämmerling alitumia aina mbili za Acetabularia ambazo zina maumbile tofauti ya cap, A. crenulata na A. mediterranea (Kielelezo\(\PageIndex{3}\)). Alikata kofia kutoka kwa aina zote mbili za seli na kisha akapandikiza kilele kutoka kwa A. crenulata kwenye mguu wa A. mediterranea, na kinyume chake. Baada ya muda, aliona kwamba kiini kilichoshirikiwa na mguu wa A. crenulata na A. kilele cha mediterranea kilianzisha kofia na morphology ya A. crenulata. Kinyume chake, kiini kilichoshirikiwa na mguu wa A. mediterranea na A. crenulata kilele kilianzisha cap na morphology ya A. mediterranea. Yeye microscopically alithibitisha kuwepo kwa nuclei katika miguu ya seli hizi na kuhusishwa maendeleo ya maumbile haya cap kwa kiini cha kila kiini iliyoshirikiwa. Kwa hiyo, alionyesha majaribio kwamba kiini kilikuwa mahali pa vifaa vya maumbile ambavyo vinaelezea mali ya seli.
Mfano mwingine wa microbial, mold nyekundu ya mkate Neurospora crassa, ulitumiwa na George Beadle na Edward Tatum kuonyesha uhusiano kati ya jeni na protini wanazozifunga. Beadle alikuwa amefanya kazi na nzizi za matunda katika maabara ya Morgan lakini alizikuta ngumu sana kufanya aina fulani za majaribio. N. crassa, kwa upande mwingine, ni viumbe rahisi na ina uwezo wa kukua kwa kati ndogo kwa sababu ina njia enzymatic ambayo inaruhusu kutumia kati ya kuzalisha vitamini yake mwenyewe na amino asidi.
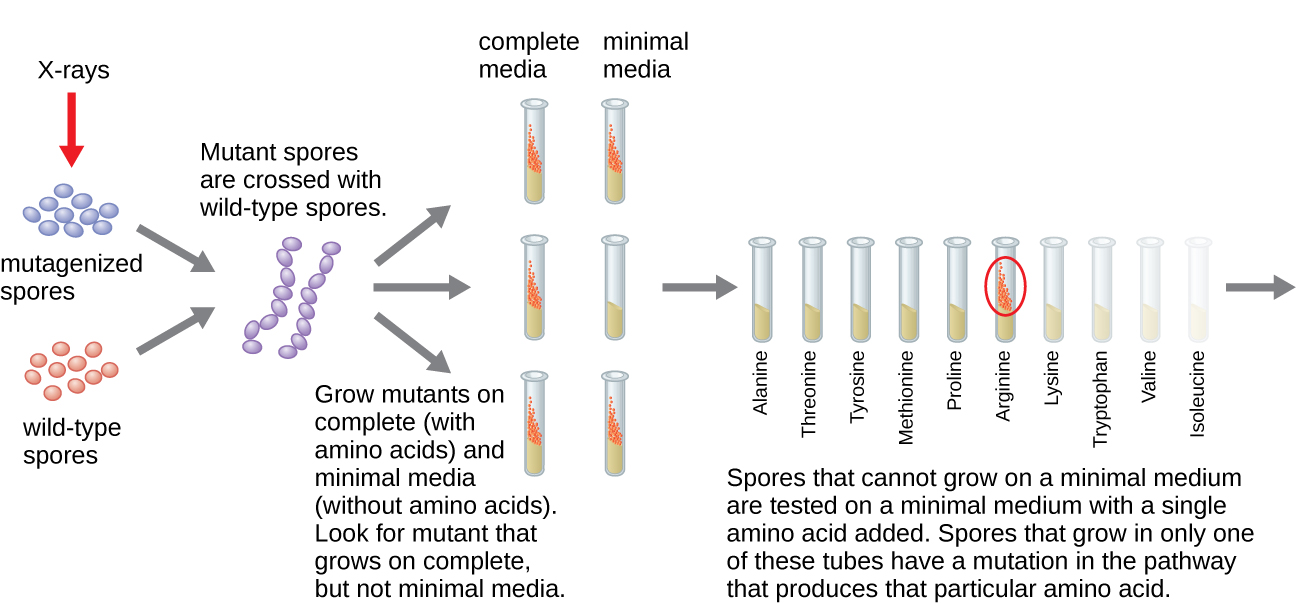
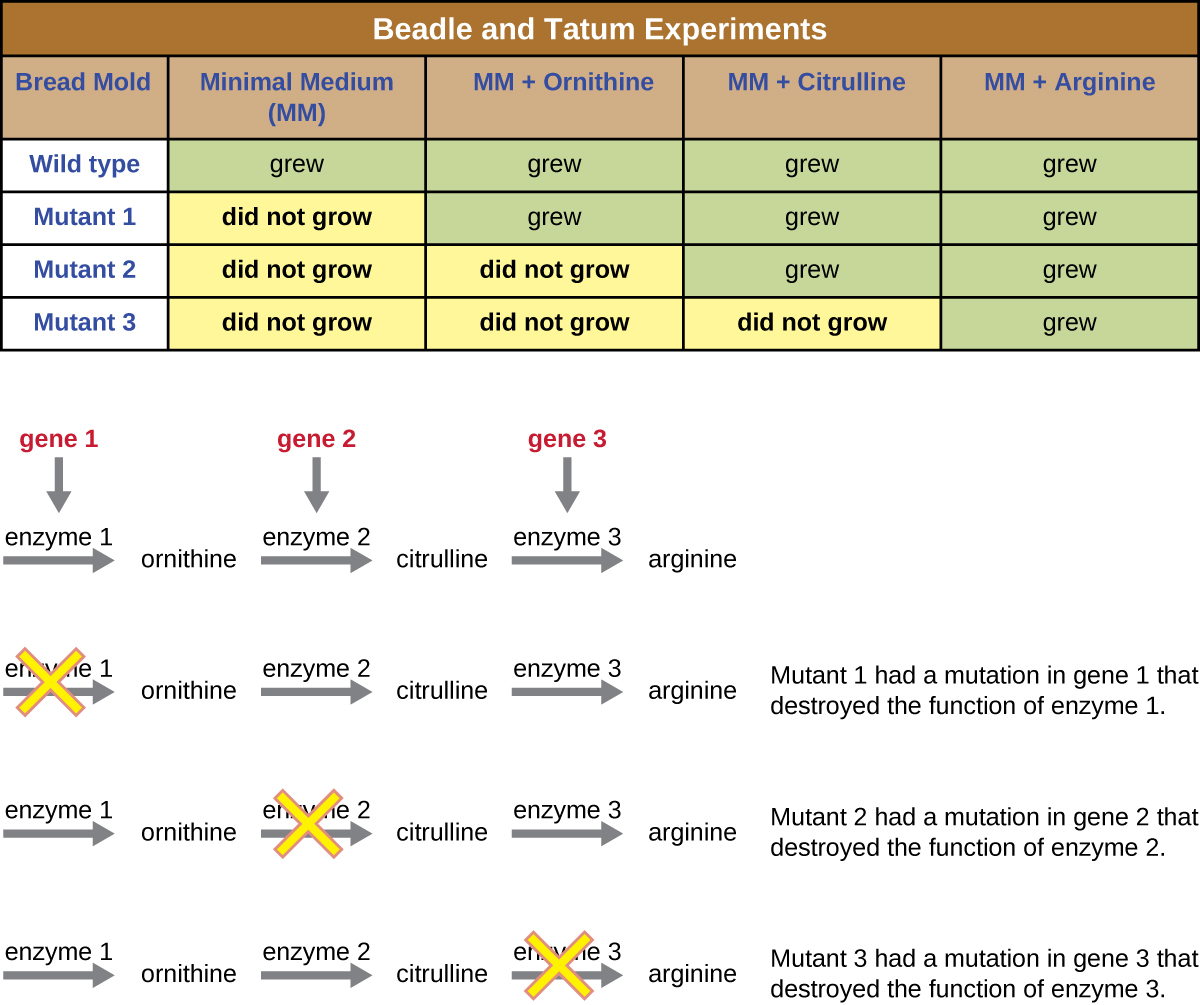
Beadle na Tatum irradiated mold na X-rays kushawishi mabadiliko kwa mlolongo wa asidi nucleic, aitwaye mutations. Wao mated irradiated mold spores na kujaribu kukua yao juu ya wote kati kamili na kati ndogo. Walitafuta mutants ambayo ilikua juu ya kati kamili, ikisaidiwa na vitamini na asidi za amino, lakini haikukua kwenye kati ndogo isiyo na virutubisho hivi. Molds vile kinadharia zilikuwa na mabadiliko katika jeni ambazo zimeandika njia za biosynthetic. Baada ya kupata mutants vile, wao utaratibu majaribio kila kuamua ambayo vitamini au amino asidi haikuweza kuzalisha (Kielelezo\(\PageIndex{4}\)) na kuchapishwa kazi hii katika 1941.
Kazi inayofuata na Beadle, Tatum, na wenzake ilionyesha kuwa wanaweza kutenganisha madarasa tofauti ya mutants ambayo yanahitaji kuongeza fulani, kama arginine ya amino asidi (Kielelezo\(\PageIndex{5}\)). Kwa ujuzi fulani wa njia ya biosynthesis ya arginine, walitambua madarasa matatu ya mutants ya arginine kwa kuongeza kati ndogo na intermediates (citrulline au ornithine) katika njia. Mutanti hizo tatu zilitofautiana katika uwezo wao wa kukua katika kila vyombo vya habari, jambo lililosababisha kundi la wanasayansi kupendekeza, mwaka wa 1945, kwamba kila aina ya mutant ilikuwa na kasoro katika jeni tofauti katika njia ya biosynthesis ya arginine. Hii ilisababisha kinachojulikana jeni moja—nadharia moja ya enzyme, ambayo ilipendekeza kwamba kila jeni husimbisha enzyme moja.
Maarifa ya baadaye kuhusu michakato ya transcription na tafsiri yalisababisha wanasayansi kurekebisha hili kwa nadharia ya “jeni-moja polipeptidi”. Ingawa kuna baadhi ya jeni ambazo hazizingatii polipeptidi (bali husimba kwa RNAs za uhamisho [trNAs] au RNAs za ribosomal [RRNAs], ambazo tutazungumzia baadaye), nadharia moja ya enzyme ya jeni—moja ni kweli katika matukio mengi, hasa katika vijidudu. Ugunduzi wa Beadle na Tatum wa uhusiano kati ya jeni na sifa sambamba uliwafanya tuzo ya Nobel ya 1958 katika Fiziolojia na Tiba na tangu hapo imekuwa msingi wa jenetiki za kisasa za Masi.
Ili kujifunza zaidi kuhusu majaribio ya Beadle na Tatum, tembelea tovuti hii kutoka Kituo cha Kujifunza DNA.
Zoezi\(\PageIndex{2}\)
- Ni viumbe gani ambavyo Morgan na wenzake walitumia kuendeleza Nadharia ya Chromosomal ya Urithi? Ni sifa gani walizofuatilia?
- Hämmerling alithibitisha nini na majaribio yake juu ya Acetabularia?
DNA kama Molekuli inayohusika na Heredity
Kufikia mwanzo wa karne ya 20, kazi kubwa ilikuwa imefanywa tayari juu ya kuashiria DNA na kuanzisha misingi ya jenetiki, ikiwa ni pamoja na kuhusisha urithi kwa kromosomu zilizopatikana ndani ya kiini. Pamoja na utafiti huu wote, haikuwa mpaka vizuri katika karne ya 20 kwamba mistari hii ya utafiti ilijiunga na wanasayansi wakaanza kufikiria kwamba DNA inaweza kuwa nyenzo za maumbile ambazo watoto walirithi kutoka kwa wazazi wao. DNA, iliyo na nucleotidi nne tu tofauti, ilidhaniwa kuwa kimuundo rahisi sana kuingiza habari hizo za maumbile tata. Badala yake, protini ilidhaniwa kuwa na utata unaohitajika kutumika kama habari za maumbile ya seli kwa sababu inaundwa na asidi amino 20 tofauti ambazo zinaweza kuunganishwa katika mchanganyiko mkubwa wa aina mbalimbali. Wataalamu wa mikrobiolojia walicheza jukumu muhimu katika utafiti ulioamua kwamba DNA ni molekuli inayohusika na urithi.
Griffith ya mabadiliko majaribio
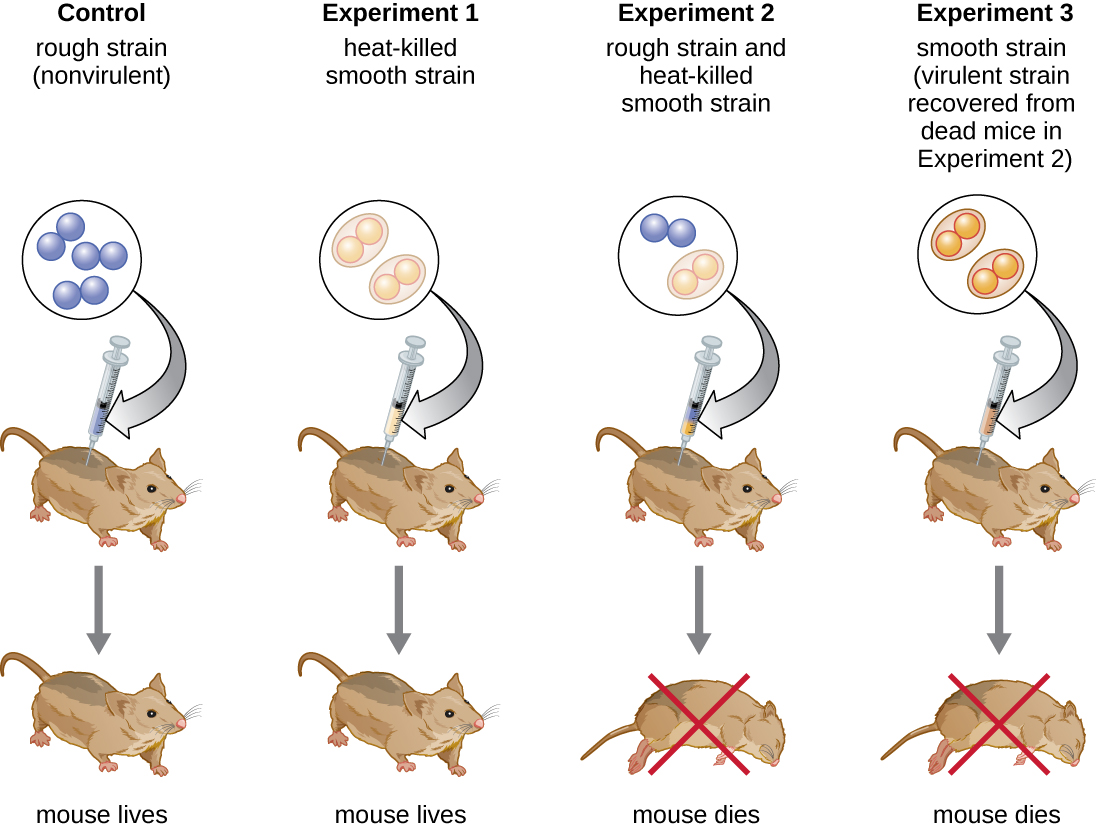
Mtaalamu wa bakteriolojia wa Uingereza Frederick Griffith (1879—1941) labda alikuwa mtu wa kwanza kuonyesha kwamba habari za urithi zinaweza kuhamishwa kutoka seli moja hadi nyingine “sambamba” (kati ya wanachama wa kizazi kimoja), badala ya “wima” (kutoka mzazi hadi uzao). Mwaka 1928, aliripoti maandamano ya kwanza ya mabadiliko ya bakteria, mchakato ambao DNA ya nje inachukuliwa na seli, na hivyo kubadilisha tabia zake. 3 Alikuwa akifanya kazi na aina mbili za Streptococcus pneumoniae, bakteria inayosababisha pneumonia: shida mbaya (R) na aina ya laini (S). Mzigo wa R hauna pathogenic na hauna capsule kwenye uso wake wa nje; Matokeo yake, makoloni kutoka kwa aina ya R yanaonekana kuwa mbaya wakati mzima kwenye sahani. Matatizo ya S ni pathogenic na ina capsule nje ya ukuta wake wa seli, kuruhusu kuepuka phagocytosis na mfumo wa kinga ya mwenyeji. Vidonge husababisha makoloni kutoka kwa aina ya S kuonekana laini wakati mzima kwenye sahani.
Katika mfululizo wa majaribio, Griffith alichambua madhara ya kuishi R, kuishi S, na joto kuuawa S Matatizo ya S. pneumoniae kwenye panya hai (Kielelezo\(\PageIndex{6}\)). Wakati panya walipoingizwa na aina ya S hai, panya walikufa. Alipoingiza panya na aina ya R hai au shida ya S iliyouawa na joto, panya zilinusurika. Lakini alipoingiza panya kwa mchanganyiko wa aina ya R hai na kuuawa na joto S strain, panya walikufa. Baada ya kutenganisha bakteria hai kutoka kwa panya iliyokufa, alipata tu aina ya S ya bakteria. Wakati yeye kisha injected hii pekee S aina katika panya safi, panya alikufa. Griffith alihitimisha kuwa kitu kilichopita kutoka kwenye aina ya S iliyouawa na joto ndani ya aina ya R hai na “kuibadilisha” kuwa aina ya pathogenic S; aliita hii “kanuni ya kubadilisha.” Majaribio haya sasa yanajulikana kama majaribio ya mabadiliko ya Griffith.
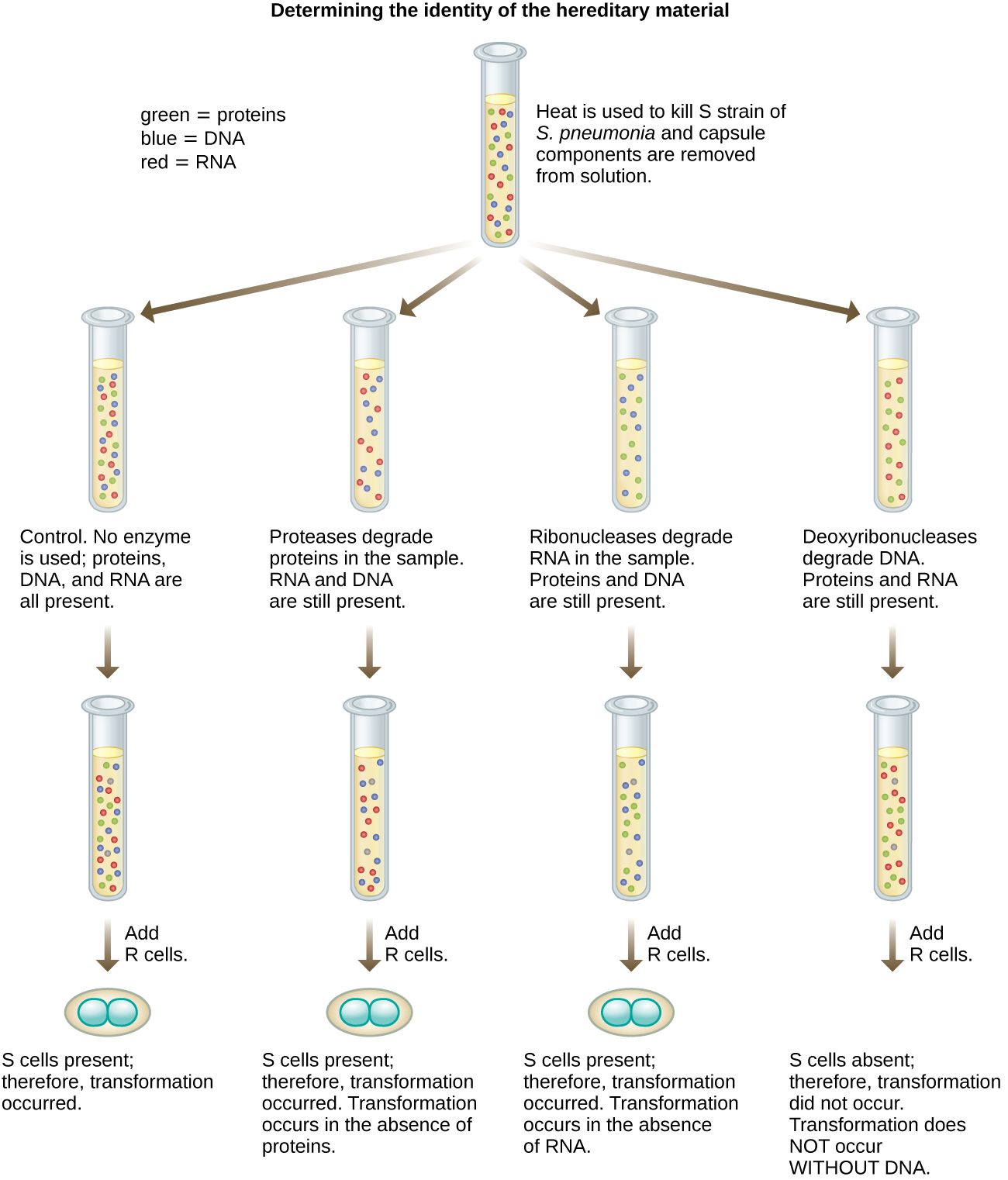
Mwaka wa 1944, Oswald Avery, Colin MacLeod, na Maclyn McCarty walikuwa na nia ya kuchunguza kanuni ya kubadilisha ya Griffith zaidi. Walitenga aina ya S kutoka kwa panya waliokufa walioambukizwa, waliuawa joto, na vipengele mbalimbali vya dondoo la S, kufanya utafiti wa utaratibu wa kuondoa (Kielelezo\(\PageIndex{7}\)). Walitumia enzymes ambazo hasa ziliharibu protini, RNA, na DNA na vikichanganya dondoo la S na kila moja ya enzymes hizi binafsi. Kisha, walijaribu kila dondo/enzyme mchanganyiko wa uwezo wa kubadilisha aina ya R, kama inavyoonekana na ukuaji wa aina ya S katika vyombo vya habari vya utamaduni na kuthibitishwa kuibua kwa ukuaji kwenye sahani. Waligundua kwamba wakati DNA ilipoharibika, mchanganyiko uliotokana haukuweza tena kubadilisha bakteria ya aina ya R, ilhali hakuna tiba nyingine ya enzymatic iliyoweza kuzuia mabadiliko. Hii iliwaongoza kuhitimisha kwamba DNA ilikuwa kanuni ya kubadilisha. Licha ya matokeo yao, wanasayansi wengi hawakukubali hitimisho lao, badala ya kuamini kuwa kulikuwa na uchafu wa protini ndani ya miche yao.
Zoezi\(\PageIndex{3}\)
Majaribio ya Avery, MacLeod, na McCarty yalionekanaje kwamba DNA ilikuwa kanuni ya kubadilisha iliyoelezwa kwanza na Griffith?
Ushahidi wa Hershey na Chase wa DNA kama Nyenzo za Maumbile
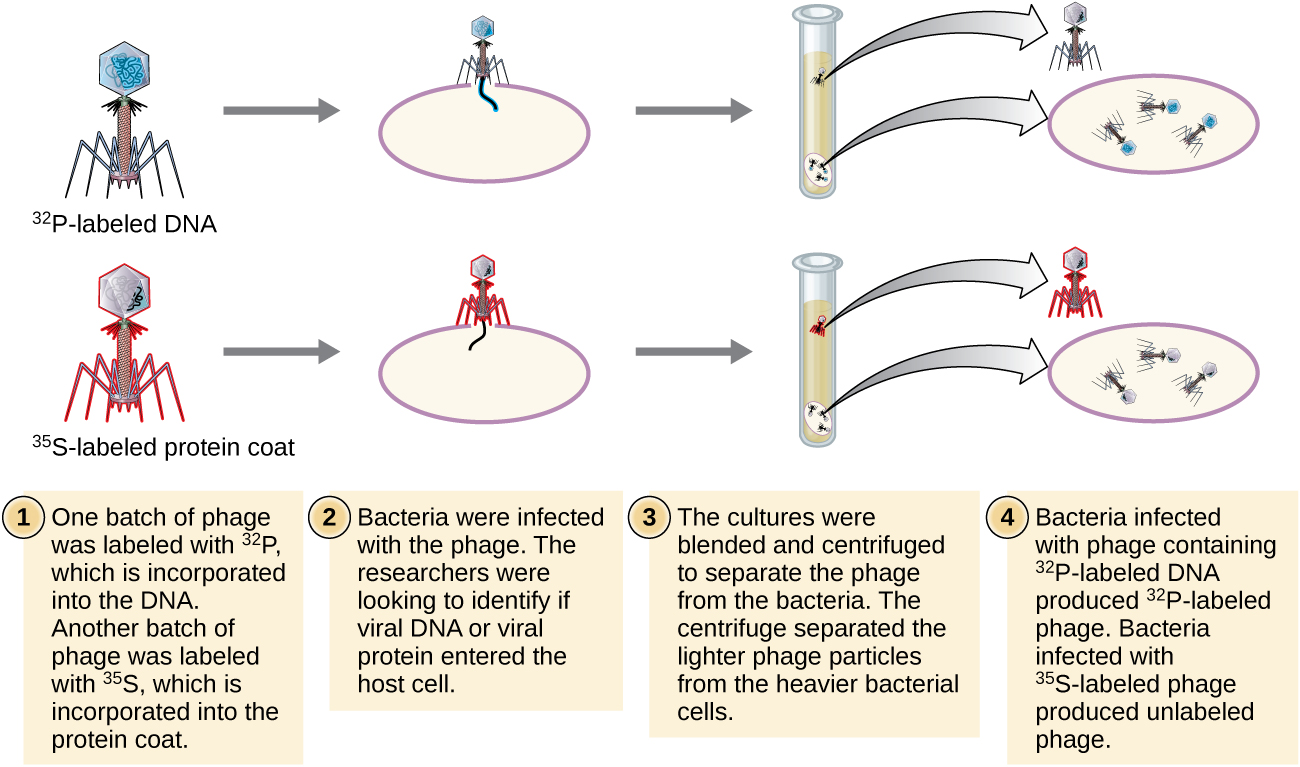
Alfred Hershey na Martha Chase walifanya majaribio yao wenyewe katika 1952 na waliweza kutoa ushahidi wa kuthibitisha kwamba DNA, si protini, ilikuwa nyenzo za maumbile (Kielelezo\(\PageIndex{8}\)). 4 Hershey na Chase walikuwa wanasoma bacteriophage, virusi vinavyoathiri bakteria. Virusi huwa na muundo rahisi: kanzu ya protini, inayoitwa capsid, na msingi wa asidi ya nucleic ambayo ina vifaa vya maumbile, ama DNA au RNA (tazama Virusi). Bacteriophage hasa waliyokuwa wakisoma ilikuwa bacteriophage ya T2, ambayo huambukiza seli za E. coli. Kama tunavyojua leo, T2 inaunganisha kwenye uso wa seli ya bakteria na kisha inaingiza asidi zake za nucleic ndani ya seli. DNA ya phage hufanya nakala nyingi yenyewe kwa kutumia mashine ya jeshi, na hatimaye kiini cha jeshi hupasuka, ikitoa idadi kubwa ya bacteriophages.
Hershey na Chase kinachoitwa kanzu ya protini katika kundi moja la phage kwa kutumia sulfuri ya mionzi, 35 S, kwa sababu sulfuri hupatikana katika methionine ya amino asidi na cysteine lakini si katika asidi ya nucleic. Wao labeled DNA katika kundi lingine kwa kutumia fosforasi mionzi, 32 P, kwa sababu fosforasi ni kupatikana katika DNA na RNA lakini si kawaida katika protini.
Kila kundi la phage liliruhusiwa kuambukiza seli tofauti. Baada ya kuambukizwa, Hershey na Chase waliweka kila kusimamishwa kwa bakteria ya phage katika blender, ambayo ilizuia nguo za phage kutoka kwenye kiini cha jeshi, na ikaacha kusimamishwa kwa centrifuge. Seli za bakteria nzito zilikaa chini na kuunda pellet, wakati chembe za phage nyepesi zilikaa katika supernatant. Katika tube na protini iliyoandikwa, radioactivity ilibakia tu katika supernatant. Katika tube na DNA iliyoandikwa, radioactivity iligunduliwa tu katika seli za bakteria. Hershey na Chase walihitimisha kuwa ilikuwa DNA ya phage iliyoingizwa ndani ya seli iliyobeba habari ili kuzalisha chembe za phage zaidi, hivyo kuthibitisha kwamba DNA, si protini, ilikuwa chanzo cha nyenzo za maumbile. Kutokana na kazi yao, jumuiya ya kisayansi ilikubali kwa upana zaidi DNA kama molekuli inayohusika na urithi.
Wakati Hershey na Chase walichapisha majaribio yao mwanzoni mwa miaka ya 1950, wanasayansi wa microbiolojia na wanasayansi wengine walikuwa wakitafiti urithi kwa zaidi ya miaka 80. Kujenga juu ya utafiti wa mtu mwingine wakati huo kilifikia kilele katika makubaliano ya jumla kwamba DNA ilikuwa nyenzo za maumbile zinazohusika na urithi (Kielelezo\(\PageIndex{9}\)). Maarifa haya yaliweka hatua kwa umri wa biolojia ya Masi ijayo na maendeleo makubwa katika bioteknolojia na mifumo ya biolojia ambayo tunapitia leo.
Ili kujifunza zaidi kuhusu majaribio yanayohusika katika historia ya jenetiki na ugunduzi wa DNA kama nyenzo za maumbile ya seli, tembelea tovuti hii kutoka Kituo cha Kujifunza DNA.
Zoezi\(\PageIndex{4}\)
Hershey na Chase walitumiaje microbes kuthibitisha kwamba DNA ni nyenzo za maumbile?
Dhana muhimu na Muhtasari
- DNA iligunduliwa na sifa muda mrefu kabla ya jukumu lake katika urithi kueleweka. Wataalamu wa mikrobiolojia walicheza majukumu muhimu katika kuonyesha kwamba DNA ni habari ya urithi inayopatikana ndani ya seli.
- Katika miaka ya 1850 na 1860, Gregor Mendel alijaribu na mbaazi za bustani za kuzaliana kweli ili kuonyesha urithi wa sifa maalum zinazoonekana.
- Mnamo mwaka wa 1869, Friedrich Miescher alitenga na kutakasa kiwanja kilicho tajiri katika fosforasi kutoka kwenye kiini cha seli nyeupe za damu; aliita jina la kiwanja. Mwanafunzi wa Miescher Richard Altmann aligundua asili yake ya tindikali, akiibadilisha tena asidi ya nucleic. Albrecht Kossell alifafanua misingi ya nucleotide iliyopatikana ndani ya asidi ya nucleic.
- Ingawa Walter Sutton na Theodor Boveri walipendekeza Nadharia ya Chromosomal ya Urithi mwaka 1902, haikuonyeshwa kisayansi hadi uchapishaji wa 1915 wa kazi ya Thomas Hunt Morgan na wenzake.
- Kwa kutumia Acetabularia, kiini kikubwa cha algali, kama mfumo wake wa mfano, Joachim Hämmerling alionyesha katika miaka ya 1930 na 1940 kwamba kiini kilikuwa mahali pa habari za urithi katika seli hizi.
- Katika miaka ya 1940, George Beadle na Edward Tatum walitumia mold Neurospora crassa kuonyesha kwamba kila uzalishaji wa protini ulikuwa chini ya udhibiti wa jeni moja, kuonyesha nadharia ya “enzyme moja—one”.
- Mnamo mwaka wa 1928, Frederick Griffith alionyesha kuwa bakteria zilizoharibika zilizokufa zinaweza kupitisha habari za maumbile ili kuishi bakteria zisizoingizwa na kuzibadilisha kuwa matatizo mabaya. Mwaka wa 1944, Oswald Avery, Colin McLeod, na Maclyn McCarty walitambua kiwanja kama DNA.
- Hali ya DNA kama molekuli inayohifadhi habari za maumbile ilionyeshwa bila usahihi katika jaribio la Alfred Hershey na Martha Chase iliyochapishwa mwaka wa 1952. Labeled DNA kutoka virusi vya bakteria aliingia na kuambukizwa seli za bakteria, na kusababisha kupanda kwa chembe zaidi ya virusi. Nguo za protini zilizoandikwa hazikushiriki katika maambukizi ya habari za maumbile.
maelezo ya chini
- 1 J.G. Mendel. “Versuche über Pflanzenhybriden.” Verhandlungen des naturforschenden Vereines katika Brünn, Bd. Abhandlungen 4 (1865) :3—7. (Kwa tafsiri ya Kiingereza, angalia http://www.mendelweb.org/Mendel.plain.html)
- 2 G.W Beadle, E.L. Tatum. “Udhibiti wa maumbile ya athari za Biochemical katika Neurospora.” Kesi za Chuo cha Taifa cha Sayansi 27 namba 11 (1941) :499—506.
- 3 F. Griffith. “Umuhimu wa Aina za Pneumococcal.” Jarida la Usafi 27 namba 2 (1928) :8—159.
- 4 AD Hershey, M. Chase. “Kazi za kujitegemea za Protini ya Virusi na Asidi ya Nucleic katika Ukuaji wa Bacteriophage.” Journal of General Physiolojia 36 namba 1 (1952) :39—56.