
10.4: A estrutura e função dos genomas celulares
- Page ID
- 181278

Objetivos de
- Defina o gene e o genótipo e diferencie o genótipo do fenótipo
- Descreva a estrutura e a embalagem dos cromossomos
- Compare os cromossomos procarióticos e eucarióticos
- Explique por que o DNA extracromossômico é importante em uma célula
Até agora, discutimos a estrutura e a função de partes individuais de DNA e RNA. Nesta seção, discutiremos como todo o material genético de um organismo — coletivamente chamado de genoma — é organizado dentro da célula. Como a genética de um organismo, em grande parte, dita suas características, não deve surpreender que os organismos difiram na disposição de seu DNA e RNA.
Genótipo versus fenótipo
Todas as atividades celulares são codificadas no DNA de uma célula. A sequência de bases dentro de uma molécula de DNA representa a informação genética da célula. Segmentos de moléculas de DNA são chamados de genes, e genes individuais contêm o código instrucional necessário para sintetizar várias proteínas, enzimas ou moléculas de RNA estáveis.
A coleção completa de genes que uma célula contém em seu genoma é chamada de genótipo. No entanto, uma célula não expressa todos os seus genes simultaneamente. Em vez disso, ele ativa (expressa) ou desliga certos genes quando necessário. O conjunto de genes que está sendo expresso em um determinado momento determina as atividades da célula e suas características observáveis, conhecidas como seu fenótipo. Genes que são sempre expressos são conhecidos como genes constitutivos; alguns genes constitutivos são conhecidos como genes domésticos porque são necessários para as funções básicas da célula.
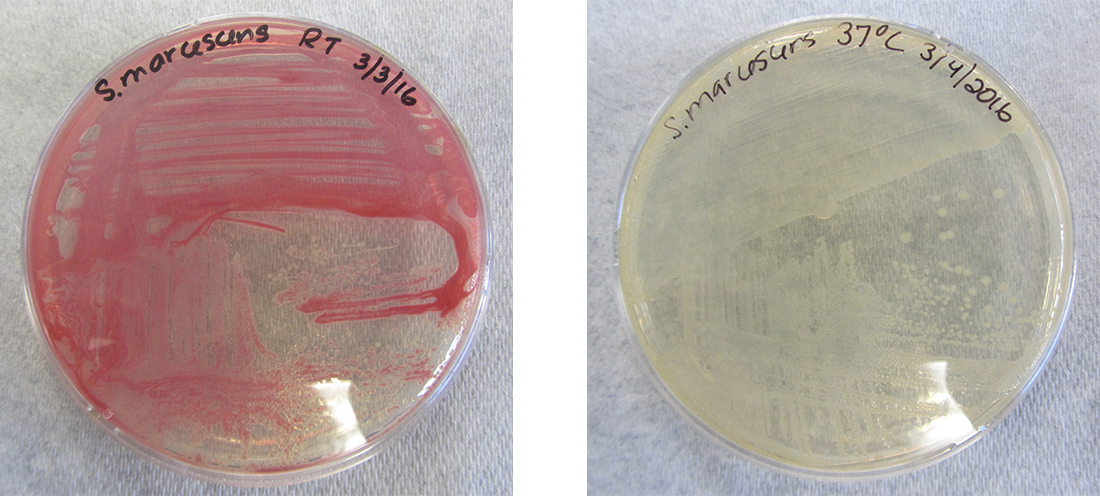
Embora o genótipo de uma célula permaneça constante, o fenótipo pode mudar em resposta a sinais ambientais (por exemplo, mudanças na temperatura ou na disponibilidade de nutrientes) que afetam quais genes não constitutivos são expressos. Por exemplo, a bactéria oral Streptococcus mutans produz uma camada de limo pegajoso que permite que ela adira aos dentes, formando placa dentária; no entanto, os genes que controlam a produção da camada de lodo só são expressos na presença de sacarose (açúcar de mesa). Assim, enquanto o genótipo de S. mutans é constante, seu fenótipo muda dependendo da presença e ausência de açúcar em seu ambiente. A temperatura também pode regular a expressão gênica. Por exemplo, a bactéria gram-negativa Serratia marcescens, um patógeno frequentemente associado a infecções adquiridas em hospitais, produz um pigmento vermelho a 28° C, mas não a 37° C, a temperatura interna normal do corpo humano (Figura\(\PageIndex{1}\)).
Organização do material genético
A grande maioria do genoma de um organismo está organizada nos cromossomos da célula, que são estruturas discretas de DNA dentro das células que controlam a atividade celular. Lembre-se de que, embora os cromossomos eucarióticos estejam alojados no núcleo ligado à membrana, a maioria dos procariontes contém um único cromossomo circular que é encontrado em uma área do citoplasma chamada nucleóide (consulte Características únicas das células procarióticas). Um cromossomo pode conter vários milhares de genes.
Organização do cromossomo eucariótico
A estrutura dos cromossomos difere um pouco entre as células eucarióticas e procarióticas. Os cromossomos eucarióticos são tipicamente lineares e as células eucarióticas contêm vários cromossomos distintos. Muitas células eucarióticas contêm duas cópias de cada cromossomo e, portanto, são diploides.
O comprimento de um cromossomo excede em muito o comprimento da célula, portanto, um cromossomo precisa ser empacotado em um espaço muito pequeno para caber dentro da célula. Por exemplo, o comprimento combinado de todos os 3 bilhões de pares de bases 1 do DNA do genoma humano mediria aproximadamente 2 metros se completamente estendido, e alguns genomas eucarióticos são muitas vezes maiores que o genoma humano. O superenrolamento de DNA se refere ao processo pelo qual o DNA é torcido para caber dentro da célula. O superenrolamento pode resultar em DNA que está subenrolado (menos de uma volta da hélice por 10 pares de bases) ou sobrecarregado (mais de uma volta por 10 pares de bases) de seu estado normal de relaxamento. As proteínas conhecidas por estarem envolvidas no superenrolamento incluem topoisomerases; essas enzimas ajudam a manter a estrutura dos cromossomos superenrolados, evitando o enrolamento excessivo do DNA durante certos processos celulares, como a replicação do DNA.
Durante o empacotamento do DNA, as proteínas de ligação ao DNA chamadas histonas realizam vários níveis de envolvimento e fixação do DNA às proteínas de sustentação. A combinação de DNA com essas proteínas ligadas é chamada de cromatina. Em eucariotos, o empacotamento do DNA pelas histonas pode ser influenciado por fatores ambientais que afetam a presença de grupos metil em certos nucleotídeos de citosina do DNA. A influência dos fatores ambientais na embalagem do DNA é chamada de epigenética. A epigenética é outro mecanismo para regular a expressão gênica sem alterar a sequência de nucleotídeos. As alterações epigenéticas podem ser mantidas por meio de várias rodadas de divisão celular e, portanto, podem ser hereditárias.
Veja esta animação do Centro de Aprendizagem de DNA para saber mais sobre embalagens de DNA em eucariotos.
Organização dos cromossomos procarióticos
Os cromossomos em bactérias e arquéias geralmente são circulares, e uma célula procariótica normalmente contém apenas um único cromossomo dentro do nucleóide. Como o cromossomo contém apenas uma cópia de cada gene, os procariontes são haplóides. Como nas células eucarióticas, o superenrolamento do DNA é necessário para que o genoma se encaixe na célula procariótica. O DNA no cromossomo bacteriano está organizado em vários domínios superenrolados. Assim como os eucariotos, as topoisomerases estão envolvidas no superenrolamento do DNA. A DNA girase é um tipo de topoisomerase, encontrada em bactérias e em algumas arquéias, que ajuda a evitar o enrolamento excessivo do DNA. (Alguns antibióticos matam bactérias atacando a DNA girase.) Além disso, proteínas semelhantes à histona se ligam ao DNA e auxiliam no empacotamento do DNA. Outras proteínas se ligam à origem da replicação, a localização no cromossomo onde a replicação do DNA se inicia. Como diferentes regiões do DNA são empacotadas de forma diferente, algumas regiões do DNA cromossômico são mais acessíveis às enzimas e, portanto, podem ser usadas mais facilmente como modelos para a expressão gênica. Curiosamente, foi demonstrado que várias bactérias, incluindo Helicobacter pylori e Shigella flexneri, induzem alterações epigenéticas em seus hospedeiros após a infecção, levando à remodelação da cromatina que pode causar efeitos a longo prazo na imunidade do hospedeiro. 2
Exercício\(\PageIndex{1}\)
- Qual é a diferença entre o genótipo de uma célula e seu fenótipo?
- Como o DNA se encaixa dentro das células?
DNA não codificante
Além dos genes, um genoma também contém muitas regiões de DNA não codificador que não codificam proteínas ou produtos de RNA estáveis. O DNA não codificante é comumente encontrado em áreas antes do início das sequências codificadoras de genes, bem como em regiões intergênicas (ou seja, sequências de DNA localizadas entre genes) (Figura\(\PageIndex{2}\)).
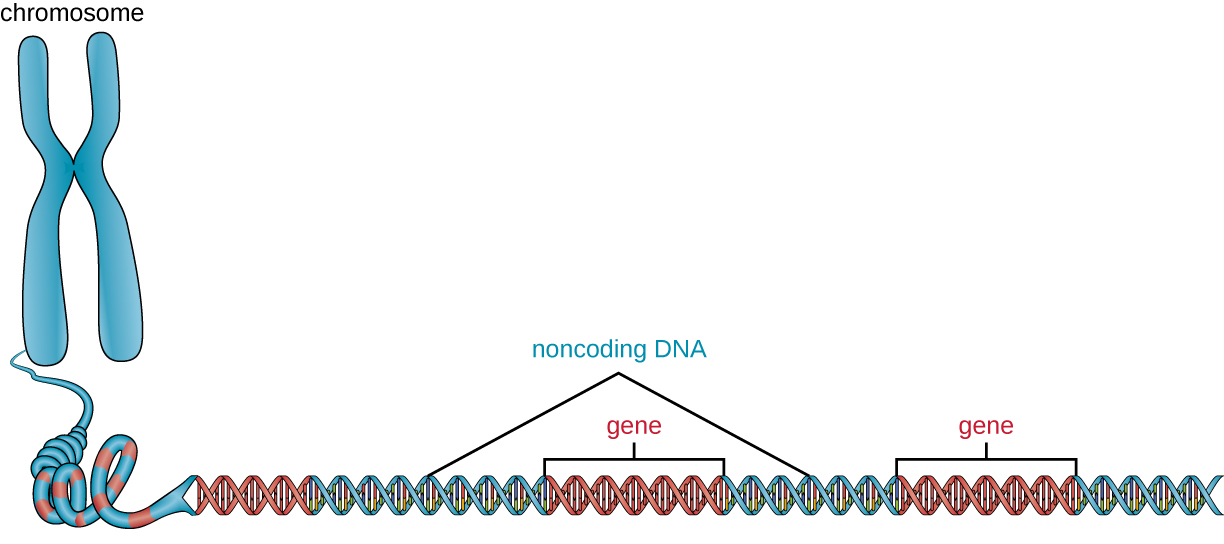
Os procariontes parecem usar seus genomas de forma muito eficiente, com apenas uma média de 12% do genoma sendo absorvido por sequências não codificantes. Em contraste, o DNA não codificante pode representar cerca de 98% do genoma em eucariotos, como visto em humanos, mas a porcentagem de DNA não codificante varia entre as espécies. 3 Essas regiões de DNA não codificantes já foram chamadas de “DNA lixo”; no entanto, essa terminologia não é mais amplamente aceita porque os cientistas, desde então, encontraram papéis para algumas dessas regiões, muitas das quais contribuem para a regulação da transcrição ou tradução por meio da produção de pequenas moléculas de RNA não codificantes, embalagem de DNA e estabilidade cromossômica. Embora os cientistas possam não compreender completamente os papéis de todas as regiões não codificadoras do DNA, geralmente acredita-se que elas tenham propósitos dentro da célula.
Exercício\(\PageIndex{2}\)
Qual é o papel do DNA não codificador?
DNA extracromossômico
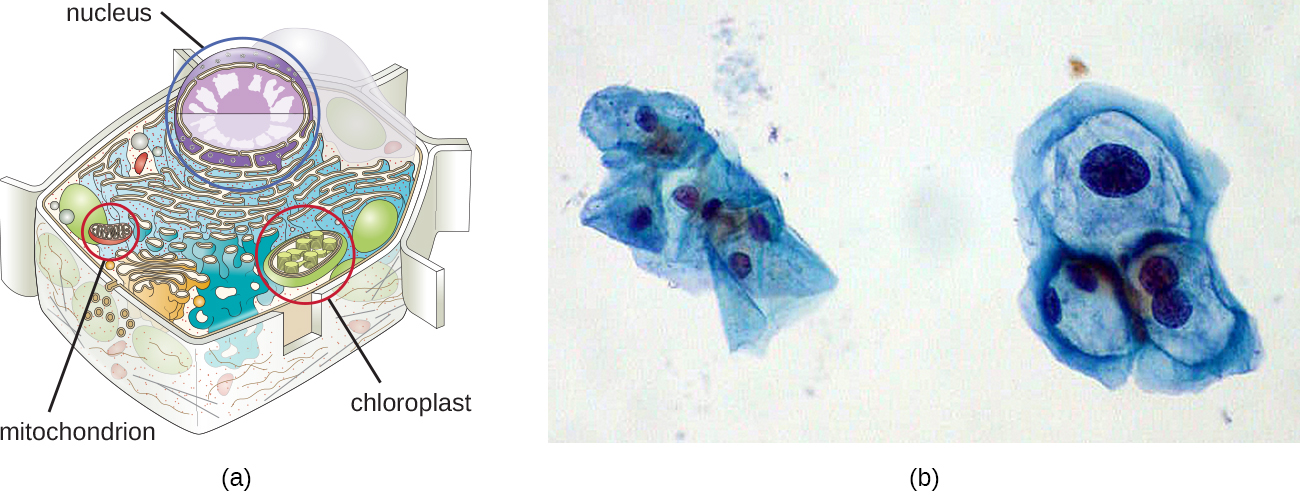
Embora a maior parte do DNA esteja contida nos cromossomos de uma célula, muitas células têm moléculas adicionais de DNA fora dos cromossomos, chamadas de DNA extracromossômico, que também fazem parte de seu genoma. Os genomas das células eucarióticas também incluiriam os cromossomos de quaisquer organelas, como mitocôndrias e/ou cloroplastos, que essas células mantêm (Figura\(\PageIndex{3}\)). A manutenção de cromossomos circulares nessas organelas é um vestígio de suas origens procarióticas e apóia a teoria endossimbiótica (veja Fundamentos da Teoria Celular Moderna). Em alguns casos, os genomas de certos vírus de DNA também podem ser mantidos de forma independente nas células hospedeiras durante a infecção viral latente. Nesses casos, esses vírus são outra forma de DNA extracromossômico. Por exemplo, o papilomavírus humano (HPV) pode ser mantido em células infectadas dessa forma.
Além dos cromossomos, alguns procariontes também têm loops menores de DNA chamados plasmídeos que podem conter um ou alguns genes não essenciais para o crescimento normal (Figura 3.3.1). As bactérias podem trocar esses plasmídeos com outras bactérias em um processo conhecido como transferência horizontal de genes (HGT). A troca de material genético em plasmídeos às vezes fornece aos micróbios novos genes benéficos para o crescimento e a sobrevivência em condições especiais. Em alguns casos, genes obtidos de plasmídeos podem ter implicações clínicas, codificando fatores de virulência que dão a um micróbio a capacidade de causar doenças ou torná-lo resistente a certos antibióticos. Os plasmídeos também são muito usados em engenharia genética e biotecnologia como forma de mover genes de uma célula para outra. O papel dos plasmídeos na transferência horizontal de genes e na biotecnologia será discutido mais adiante em Mecanismos de Genética Microbiana e Aplicações Modernas da Genética Microbiana.
Exercício\(\PageIndex{3}\)
Como os plasmídeos estão envolvidos na resistência aos antibióticos?
Plasmídeos letais
Maria, uma estudante de antropologia de 20 anos do Texas, adoeceu recentemente na nação africana de Botswana, onde estava conduzindo pesquisas como parte de um programa de estudos no exterior. A pesquisa de Maria foi focada nos métodos tradicionais africanos de curtimento de peles para a produção de couro. Durante um período de três semanas, ela visitou um curtume diariamente por várias horas para observar e participar do processo de bronzeamento. Um dia, depois de voltar do curtume, Maria desenvolveu febre, calafrios e dor de cabeça, além de dores no peito, dores musculares, náuseas e outros sintomas semelhantes aos da gripe. Inicialmente, ela não estava preocupada, mas quando a febre aumentou e ela começou a tossir sangue, sua família anfitriã africana ficou alarmada e a levou às pressas para o hospital, onde sua condição continuava piorando.
Depois de saber sobre seu trabalho recente no curtume, a médica suspeitou que Maria havia sido exposta ao antraz. Ele pediu uma radiografia de tórax, uma amostra de sangue e uma punção lombar e imediatamente iniciou um curso de penicilina intravenosa. Infelizmente, os exames laboratoriais confirmaram o diagnóstico presuntivo do médico. A radiografia de tórax de Maria exibiu derrame pleural, o acúmulo de líquido no espaço entre as membranas pleurais e uma coloração de Gram de seu sangue revelou a presença de bactérias gram-positivas em forma de bastonete em cadeias curtas, consistentes com Bacillus anthracis. Também foi demonstrado que sangue e bactérias estavam presentes em seu líquido cefalorraquidiano, indicando que a infecção havia evoluído para meningite. Apesar do tratamento de apoio e da antibioticoterapia agressiva, Maria entrou em um estado de inatividade e morreu três dias depois.
O antraz é uma doença causada pela introdução de endosporos da bactéria gram-positiva B. anthracis no corpo. Uma vez infectados, os pacientes geralmente desenvolvem meningite, geralmente com resultados fatais. No caso de Maria, ela inalou os endosporos enquanto manuseava as peles de animais que haviam sido infectados.
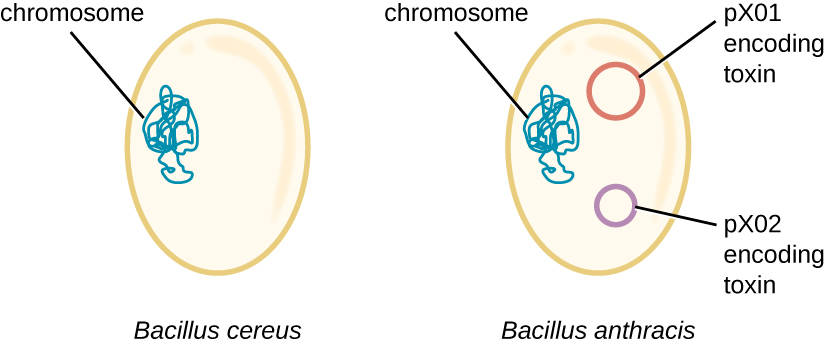
O genoma de B. anthracis ilustra como pequenas diferenças estruturais podem levar a grandes diferenças na virulência. Em 2003, os genomas de B. anthracis e Bacillus cereus, uma bactéria similar, mas menos patogênica, do mesmo gênero, foram sequenciados e comparados. 4 Os pesquisadores descobriram que as sequências do gene 16S rRNA dessas bactérias são mais de 99% idênticas, o que significa que elas são na verdade membros da mesma espécie, apesar de sua classificação tradicional como espécies separadas. Embora suas sequências cromossômicas também tenham revelado muita semelhança, descobriu-se que vários fatores de virulência de B. anthracis estão codificados em dois grandes plasmídeos não encontrados em B. cereus. O plasmídeo pX01 codifica uma toxina de três partes que suprime o sistema imunológico do hospedeiro, enquanto o plasmídeo pX02 codifica um polissacarídeo capsular que protege ainda mais a bactéria do sistema imunológico do hospedeiro (Figura\(\PageIndex{4}\)). Como o B. cereus não possui esses plasmídeos, ele não produz esses fatores de virulência e, embora ainda seja patogênico, geralmente está associado a casos leves de diarreia, dos quais o corpo pode se recuperar rapidamente. Infelizmente para Maria, a presença desses plasmídeos codificadores de toxinas em B. anthracis lhe confere sua virulência letal.
Exercício\(\PageIndex{4}\)
O que você acha que aconteceria com a patogenicidade do B. anthracis se ele perdesse um ou ambos os plasmídeos?
Foco clínico: Resolução
Em 24 horas, os resultados da análise do teste diagnóstico da amostra de fezes de Alex revelaram que ela foi positiva para enterotoxina termo-lábil (LT), enterotoxina termo-estável (ST) e fator de colonização (CF), confirmando a suspeita do médico do hospital sobre ETEC. Durante um acompanhamento com o médico de família de Alex, esse médico observou que os sintomas de Alex não estavam se resolvendo rapidamente e ele estava sentindo um desconforto que o impedia de voltar às aulas. O médico de família prescreveu a Alex um curso de ciprofloxacina para resolver seus sintomas. Felizmente, a ciprofloxacina resolveu os sintomas de Alex em poucos dias.
Alex provavelmente contraiu a infecção ao ingerir água ou comida contaminada. Países industrializados emergentes, como o México, ainda estão desenvolvendo práticas de saneamento que evitam a contaminação da água com material fecal. Os viajantes nesses países devem evitar a ingestão de alimentos mal cozidos, especialmente carnes, frutos do mar, vegetais e laticínios não pasteurizados. Eles também devem evitar o uso de água que não tenha sido tratada; isso inclui água potável, cubos de gelo e até mesmo água usada para escovar os dentes. Usar água engarrafada para esses propósitos é uma boa alternativa. Uma boa higiene (lavar as mãos) também pode ajudar na prevenção de uma infecção por ETEC. Alex não tinha sido cuidadoso com o consumo de comida ou água, o que o levou à doença.
Os sintomas de Alex eram muito semelhantes aos da cólera, causada pela bactéria gram-negativa Vibrio cholerae, que também produz uma toxina semelhante à ST e LT. Em algum momento da história evolutiva da ETEC, uma cepa não patogênica de E. coli semelhante às normalmente encontradas no intestino pode ter adquirido os genes que codificam as toxinas ST e LT de V. cholerae. O fato de os genes que codificam essas toxinas serem codificados em plasmídeos extracromossômicos na ETEC apóia a ideia de que esses genes foram adquiridos por E. coli e provavelmente são mantidos em populações bacterianas por meio da transferência horizontal de genes.
Genomas virais
Os genomas virais apresentam uma diversidade significativa na estrutura. Alguns vírus têm genomas que consistem em DNA como material genético. Esse DNA pode ser de fita simples, como exemplificado por parvovírus humanos, ou fita dupla, como visto nos herpesvírus e poxvírus. Além disso, embora toda a vida celular use DNA como material genético, alguns genomas virais são feitos de moléculas de RNA de fita simples ou dupla, como discutimos. Os genomas virais são normalmente menores do que a maioria dos genomas bacterianos, codificando apenas alguns genes, porque eles dependem de seus hospedeiros para realizar muitas das funções necessárias para sua replicação. A diversidade das estruturas do genoma viral e suas implicações para os ciclos de vida da replicação viral são discutidas com mais detalhes em The Viral Life Cycle.
Exercício\(\PageIndex{5}\)
Por que os genomas virais variam muito entre os vírus?
O tamanho do genoma importa
Há uma grande variação no tamanho dos genomas entre diferentes organismos. A maioria dos eucariotos mantém vários cromossomos; os humanos, por exemplo, têm 23 pares, o que lhes dá 46 cromossomos. Apesar de ser grande, com 3 bilhões de pares de bases, o genoma humano está longe de ser o maior genoma. As plantas geralmente mantêm genomas muito grandes, até 150 bilhões de pares de bases, e geralmente são poliplóides, com várias cópias de cada cromossomo.
O tamanho dos genomas bacterianos também varia consideravelmente, embora eles tendam a ser menores do que os genomas eucarióticos (Figura\(\PageIndex{5}\)). Alguns genomas bacterianos podem ser tão pequenos quanto apenas 112.000 pares de bases. Freqüentemente, o tamanho do genoma de uma bactéria está diretamente relacionado ao quanto a bactéria depende de seu hospedeiro para sobreviver. Quando uma bactéria depende da célula hospedeira para realizar determinadas funções, ela perde os genes que codificam as habilidades de realizar essas funções sozinha. Esses tipos de endossimbiontes bacterianos lembram as origens procarióticas das mitocôndrias e dos cloroplastos.
Do ponto de vista clínico, os patógenos intracelulares obrigatórios também tendem a ter genomas pequenos (cerca de 1 milhão de pares de bases). Como as células hospedeiras fornecem a maioria de seus nutrientes, elas tendem a ter um número reduzido de genes que codificam as funções metabólicas. Devido aos seus tamanhos pequenos, os genomas de organismos como Mycoplasma genitalium (580.000 pares de bases), Chlamydia trachomatis (1,0 milhão), Rickettsia prowazekii (1,1 milhão) e Treponema pallidum (1,1 milhão) foram alguns dos genomas bacterianos anteriores sequenciado. Respectivamente, esses patógenos causam uretrite e inflamação pélvica, clamídia, tifo e sífilis.
Enquanto os patógenos intracelulares obrigatórios têm genomas incomumente pequenos, outras bactérias com uma grande variedade de capacidades metabólicas e enzimáticas têm genomas bacterianos incomumente grandes. A Pseudomonas aeruginosa, por exemplo, é uma bactéria comumente encontrada no meio ambiente e é capaz de crescer em uma ampla variedade de substratos. Seu genoma contém 6,3 milhões de pares de bases, o que lhe confere uma alta capacidade metabólica e a capacidade de produzir fatores de virulência que causam vários tipos de infecções oportunistas.
Curiosamente, também houve uma variabilidade significativa no tamanho do genoma em vírus, variando de 3.500 pares de bases a 2,5 milhões de pares de bases, excedendo significativamente o tamanho de muitos genomas bacterianos. A grande variação observada nos tamanhos do genoma viral contribui ainda mais para a grande diversidade de características do genoma viral já discutidas.
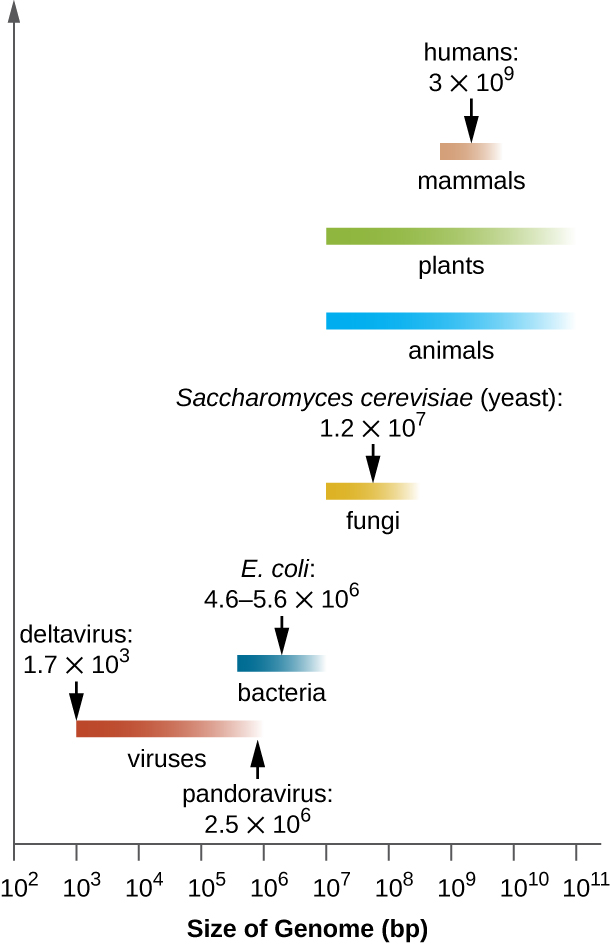
Visite o banco de dados de genomas do National Center for Biotechnology Information (NCBI) para ver os genomas que foram sequenciados e seus tamanhos.
Conceitos principais e resumo
- Todo o conteúdo genético de uma célula é seu genoma.
- Os genes codificam proteínas, ou moléculas de RNA estáveis, cada uma das quais desempenha uma função específica na célula.
- Embora o genótipo que uma célula possui permaneça constante, a expressão dos genes depende das condições ambientais.
- Um fenótipo é a característica observável de uma célula (ou organismo) em um determinado momento e resulta do complemento de genes atualmente em uso.
- A maioria do material genético é organizada em cromossomos que contêm o DNA que controla as atividades celulares.
- Os procariontes são tipicamente haplóides, geralmente com um único cromossomo circular encontrado no nucleóide. Os eucariotos são diplóides; o DNA é organizado em vários cromossomos lineares encontrados no núcleo.
- O superenrolamento e o empacotamento de DNA usando proteínas de ligação ao DNA permitem que moléculas longas se encaixem dentro de uma célula. Eucariotos e arquéias usam proteínas histonas, e as bactérias usam proteínas diferentes com função semelhante.
- Os genomas procarióticos e eucarióticos contêm DNA não codificante, cuja função não é bem compreendida. Alguns DNA não codificadores parecem participar da formação de pequenas moléculas de RNA não codificantes que influenciam a expressão gênica; alguns parecem desempenhar um papel na manutenção da estrutura cromossômica e no empacotamento do DNA.
- O DNA extracromossômico em eucariotos inclui os cromossomos encontrados nas organelas de origem procariótica (mitocôndrias e cloroplastos) que evoluíram por endossimbiose. Alguns vírus também podem se manter extracromossomicamente.
- O DNA extracromossômico em procariontes é comumente mantido como plasmídeos que codificam alguns genes não essenciais que podem ser úteis em condições específicas. Os plasmídeos podem se espalhar por meio de uma comunidade bacteriana por transferência horizontal de genes.
- Os genomas virais apresentam ampla variação e podem ser compostos de RNA ou DNA e podem ser de fita dupla ou simples.
Notas de pé
- 1 Instituto Nacional de Pesquisa do Genoma Humano. “A conclusão do projeto do genoma humano: perguntas frequentes.” https://www.genome.gov/11006943. Acessado em 10 de junho de 2016
- 2 H. Bierne et al. “Epigenética e infecções bacterianas”. Perspectivas de Cold Spring Harbor na Medicina 2 nº 12 (2012) :a010272.
- 3 R.J. Taft e cols.. “A relação entre DNA não codificador de proteínas e complexidade eucariótica.” Bioensaios 29 nº 3 (2007) :288—299.
- 4 N. Ivanova et al. “Sequência do genoma de Bacillus cereus e análise comparativa com Bacillus anthracis.” Nature 423 nº 6935 (2003) :87—91.