
15.2: Transcrição procariótica
- Page ID
- 182292

Habilidades para desenvolver
- Liste as diferentes etapas da transcrição procariótica
- Discuta o papel dos promotores na transcrição procariótica
- Descreva como e quando a transcrição é encerrada
Os procariontes, que incluem bactérias e arquéias, são em sua maioria organismos unicelulares que, por definição, não possuem núcleos ligados à membrana e outras organelas. Um cromossomo bacteriano é um círculo fechado covalentemente que, ao contrário dos cromossomos eucarióticos, não está organizado em torno de proteínas histonas. A região central da célula na qual o DNA procariótico reside é chamada de nucleóide. Além disso, os procariontes geralmente têm plasmídeos abundantes, que são moléculas circulares de DNA mais curtas que podem conter apenas um ou alguns genes. Os plasmídeos podem ser transferidos independentemente do cromossomo bacteriano durante a divisão celular e geralmente carregam características como resistência a antibióticos.
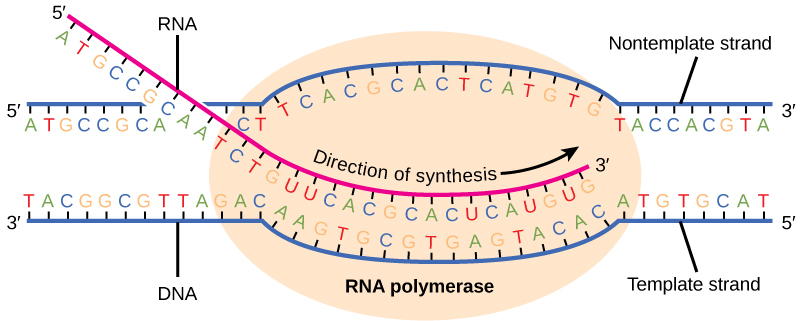
A transcrição em procariontes (e em eucariotos) requer que a dupla hélice do DNA se desenrole parcialmente na região da síntese de mRNA. A região de desenrolamento é chamada de bolha de transcrição. A transcrição sempre procede da mesma fita de DNA para cada gene, que é chamada de fita modelo. O produto de mRNA é complementar à fita modelo e é quase idêntico à outra fita de DNA, chamada de fita não modelo. A única diferença é que no mRNA, todos os nucleotídeos T são substituídos por nucleotídeos U. Em uma dupla hélice de RNA, A pode ligar U por meio de duas ligações de hidrogênio, assim como no emparelhamento A-T em uma dupla hélice de DNA.
O par de nucleotídeos na dupla hélice do DNA que corresponde ao local a partir do qual o primeiro nucleotídeo de 5' mRNA é transcrito é chamado de sítio +1, ou local de iniciação. Os nucleotídeos que precedem o local de iniciação recebem números negativos e são designados a montante. Por outro lado, os nucleotídeos após o local de iniciação são indicados com a numeração “+” e são chamados de nucleotídeos a jusante.
Início da transcrição em procariontes
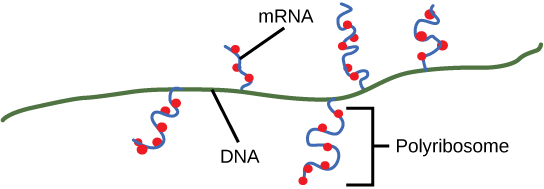
Os procariontes não têm núcleos fechados por membranas. Portanto, os processos de transcrição, tradução e degradação do mRNA podem ocorrer simultaneamente. O nível intracelular de uma proteína bacteriana pode ser rapidamente amplificado por vários eventos de transcrição e tradução que ocorrem simultaneamente no mesmo modelo de DNA. A transcrição procariótica geralmente cobre mais de um gene e produz mRNAs policistrônicos que especificam mais de uma proteína.
Nossa discussão aqui exemplificará a transcrição descrevendo esse processo em Escherichia coli, uma espécie bacteriana bem estudada. Embora existam algumas diferenças entre a transcrição em E. coli e a transcrição em arquéias, uma compreensão da transcrição de E. coli pode ser aplicada a praticamente todas as espécies bacterianas.
RNA polimerase procariótica
Os procariontes usam a mesma RNA polimerase para transcrever todos os seus genes. Em E. coli, a polimerase é composta por cinco subunidades polipeptídicas, duas das quais são idênticas. Quatro dessas subunidades, denotadas α, α, β e β', compreendem a enzima central da polimerase. Essas subunidades se reúnem toda vez que um gene é transcrito e se desmontam quando a transcrição é concluída. Cada subunidade tem um papel único; as duas subunidades α são necessárias para montar a polimerase no DNA; a subunidade β se liga ao trifosfato ribonucleosídeo que se tornará parte da molécula nascente de mRNA “recém-nascida”; e o β' se liga à fita modelo de DNA. A quinta subunidade, σ, está envolvida somente no início da transcrição. Ele confere especificidade transcricional de tal forma que a polimerase começa a sintetizar o mRNA a partir de um local de iniciação apropriado. Sem σ, a enzima central transcreveria de locais aleatórios e produziria moléculas de mRNA que especificavam bobagens proteicas. A polimerase composta por todas as cinco subunidades é chamada de holoenzima.
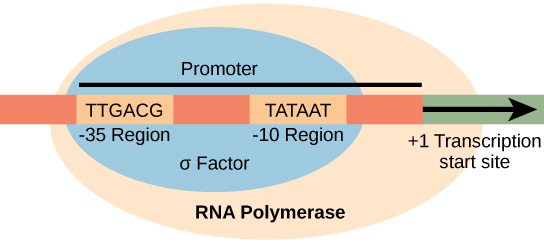
Promotores procarióticos
Um promotor é uma sequência de DNA na qual o mecanismo de transcrição se liga e inicia a transcrição. Na maioria dos casos, os promotores existem a montante dos genes que eles regulam. A sequência específica de um promotor é muito importante porque determina se o gene correspondente é transcrito o tempo todo, em parte ou com pouca frequência. Embora os promotores variem entre os genomas procarióticos, alguns elementos são conservados. Nas regiões -10 e -35 a montante do local de iniciação, existem duas sequências de consenso do promotor, ou regiões que são semelhantes em todos os promotores e em várias espécies bacterianas (Figura\(\PageIndex{1}\)). A sequência de consenso -10, chamada de região -10, é TATAAT. A sequência -35, TTGACA, é reconhecida e limitada por σ. Uma vez que essa interação é feita, as subunidades da enzima central se ligam ao local. A região -10 rica em A — T facilita o desenrolamento do modelo de DNA e várias ligações de fosfodiéster são feitas. A fase de início da transcrição termina com a produção de transcritos abortivos, que são polímeros de aproximadamente 10 nucleotídeos que são feitos e liberados.
Link para o aprendizado
Veja esta animação da MolecularMovies para ver a primeira parte da transcrição e a repetição da sequência básica da caixa TATA.
Alongamento e terminação em procariontes
A fase de alongamento da transcrição começa com a liberação da subunidade σ da polimerase. A dissociação de σ permite que a enzima central prossiga ao longo do modelo de DNA, sintetizando mRNA na direção 5' a 3' a uma taxa de aproximadamente 40 nucleotídeos por segundo. À medida que o alongamento prossegue, o DNA é continuamente desenrolado à frente da enzima central e rebobinado atrás dela (Figura\(\PageIndex{2}\)). O emparelhamento de bases entre DNA e RNA não é estável o suficiente para manter a estabilidade dos componentes da síntese de mRNA. Em vez disso, a RNA polimerase atua como um ligante estável entre o modelo de DNA e as fitas nascentes de RNA para garantir que o alongamento não seja interrompido prematuramente.
Sinais de terminação procariótica
Depois que um gene é transcrito, a polimerase procariótica precisa ser instruída a se dissociar do modelo de DNA e liberar o mRNA recém-criado. Dependendo do gene que está sendo transcrito, existem dois tipos de sinais de terminação. Um é baseado em proteína e o outro é baseado em RNA. A terminação dependente de Rho é controlada pela proteína rho, que segue por trás da polimerase na cadeia crescente de mRNA. Perto do final do gene, a polimerase encontra uma série de nucleotídeos G no modelo de DNA e ela para. Como resultado, a proteína rho colide com a polimerase. A interação com rho libera o mRNA da bolha de transcrição.
A terminação independente de Rho é controlada por sequências específicas na fita modelo de DNA. À medida que a polimerase se aproxima do final da transcrição do gene, ela encontra uma região rica em nucleotídeos C—G. O mRNA se dobra sobre si mesmo e os nucleotídeos C — G complementares se unem. O resultado é um gancho estável que faz com que a polimerase pare assim que começa a transcrever uma região rica em nucleotídeos A — T. A região U—A complementar do transcrito do mRNA forma apenas uma interação fraca com o DNA modelo. Isso, juntamente com a polimerase estagnada, induz instabilidade suficiente para que a enzima central se separe e libere a nova transcrição do mRNA.
Após o término, o processo de transcrição é concluído. No momento em que o término ocorre, a transcrição procariótica já teria sido usada para iniciar a síntese de várias cópias da proteína codificada porque esses processos podem ocorrer simultaneamente. A unificação da transcrição, tradução e até mesmo degradação do mRNA é possível porque todos esses processos ocorrem na mesma direção de 5' a 3' e porque não há compartimentalização membranosa na célula procariótica (Figura\(\PageIndex{3}\)). Em contraste, a presença de um núcleo nas células eucarióticas impede a transcrição e tradução simultâneas.
Link para o aprendizado
Visite esta animação do BioStudio para ver o processo de transcrição procariótica.
Resumo
Em procariontes, a síntese de mRNA é iniciada em uma sequência promotora no modelo de DNA compreendendo duas sequências de consenso que recrutam a RNA polimerase. A polimerase procariótica consiste em uma enzima central de quatro subunidades proteicas e uma proteína σ que auxilia apenas na iniciação. O alongamento sintetiza o mRNA na direção 5' a 3' a uma taxa de 40 nucleotídeos por segundo. A terminação libera o mRNA e ocorre pela interação da proteína rho ou pela formação de um gancho de mRNA.
Glossário
- consenso
- Sequência de DNA usada por muitas espécies para realizar funções iguais ou similares
- enzima central
- RNA polimerase procariótica consistindo de α, α, β e β', mas sem σ; esse complexo realiza alongamento
- Rio abaixo
- nucleotídeos seguindo o local de iniciação na direção da transcrição do mRNA; em geral, sequências que estão na extremidade 3' em relação a um local no mRNA
- grampo
- estrutura do RNA quando ele se dobra sobre si mesmo e forma ligações de hidrogênio intramoleculares entre nucleotídeos complementares
- holoenzima
- RNA polimerase procariótica que consiste em α, α, β, β' e σ; esse complexo é responsável pelo início da transcrição
- local de iniciação
- nucleotídeo a partir do qual a síntese de mRNA prossegue na direção 5' a 3'; denotado com um “+1”
- fio não modelo
- fita de DNA que não é usada para transcrever mRNA; essa fita é idêntica ao mRNA, exceto que os nucleotídeos T no DNA são substituídos por nucleotídeos U no mRNA
- plasmídeo
- molécula de DNA circular extracromossômica, covalentemente fechada que pode conter apenas um ou alguns genes; comum em procariontes
- promoter
- Sequência de DNA à qual a RNA polimerase e fatores associados se ligam e iniciam a transcrição
- Terminação dependente de Rho
- em procariontes, terminação da transcrição por uma interação entre a RNA polimerase e a proteína rho em uma série de nucleotídeos G no modelo de DNA
- Independente de RHO
- terminação dependente da sequência de terminação da síntese de mRNA procariótico; causada pela formação de ganchos no mRNA que impede a polimerase
- Caixa TATA
- sequência promotora conservada em eucariotos e procariontes que ajuda a estabelecer o local de iniciação para a transcrição
- vertente modelo
- fita de DNA que especifica a molécula complementar de mRNA
- bolha de transcrição
- região de DNA localmente desenrolado que permite a transcrição do mRNA
- corrente acima
- nucleotídeos que precedem o local de iniciação; em geral, sequências em direção à extremidade 5' em relação a um local no mRNA