
19.2: Crescimento populacional e regulação
- Page ID
- 179051

Os ecologistas populacionais usam uma variedade de métodos para modelar a dinâmica populacional. Um modelo preciso deve ser capaz de descrever as mudanças que ocorrem em uma população e prever mudanças futuras.
Crescimento populacional
Os dois modelos mais simples de crescimento populacional usam equações determinísticas (equações que não levam em conta eventos aleatórios) para descrever a taxa de mudança no tamanho de uma população ao longo do tempo. O primeiro desses modelos, crescimento exponencial, descreve populações teóricas que aumentam em números sem limites para seu crescimento. O segundo modelo, o crescimento logístico, introduz limites ao crescimento reprodutivo que se tornam mais intensos à medida que o tamanho da população aumenta. Nenhum dos modelos descreve adequadamente as populações naturais, mas eles fornecem pontos de comparação.
Crescimento exponencial
Charles Darwin, ao desenvolver sua teoria da seleção natural, foi influenciado pelo clérigo inglês Thomas Malthus. Malthus publicou seu livro em 1798 afirmando que as populações com recursos naturais abundantes crescem muito rapidamente; no entanto, elas limitam o crescimento ao esgotar seus recursos. O padrão inicial de aceleração do tamanho da população é chamado de crescimento exponencial.
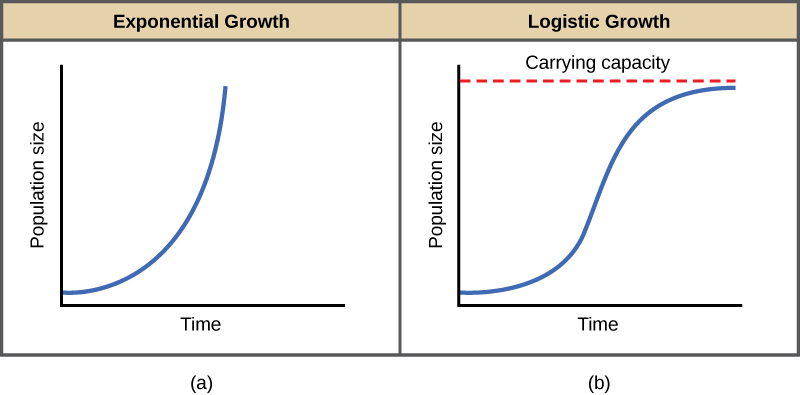
O melhor exemplo de crescimento exponencial em organismos é visto em bactérias. As bactérias são procariontes que se reproduzem em grande parte por fissão binária. Essa divisão leva cerca de uma hora para muitas espécies bacterianas. Se 1000 bactérias forem colocadas em um frasco grande com um suprimento abundante de nutrientes (para que os nutrientes não se esgotem rapidamente), o número de bactérias terá dobrado de 1000 para 2000 em apenas uma hora. Em outra hora, cada uma das 2000 bactérias se dividirá, produzindo 4000 bactérias. Após a terceira hora, deve haver 8000 bactérias no frasco. O conceito importante de crescimento exponencial é que a taxa de crescimento - o número de organismos adicionados em cada geração reprodutiva - está aumentando em si mesma; ou seja, o tamanho da população está aumentando a uma taxa cada vez maior. Após 24 desses ciclos, a população teria aumentado de 1000 para mais de 16 bilhões de bactérias. Quando o tamanho da população, N, é traçado ao longo do tempo, uma curva de crescimento em forma de J é produzida (Figura\(\PageIndex{1}\) a).
O exemplo da bactéria em um frasco não é verdadeiramente representativo do mundo real, onde os recursos geralmente são limitados. No entanto, quando uma espécie é introduzida em um novo habitat que considere adequado, ela pode apresentar crescimento exponencial por um tempo. No caso das bactérias no frasco, algumas bactérias morrerão durante o experimento e, portanto, não se reproduzirão; portanto, a taxa de crescimento é reduzida de uma taxa máxima na qual não há mortalidade. A taxa de crescimento de uma população é amplamente determinada pela subtração da taxa de mortalidade, D, (número de organismos que morrem durante um intervalo) da taxa de natalidade, B, (número de organismos que nascem durante um intervalo). A taxa de crescimento pode ser expressa em uma equação simples que combina as taxas de natalidade e mortalidade em um único fator: r. Isso é mostrado na seguinte fórmula:
\[\text{Population growth} = rN \nonumber\]
O valor de r pode ser positivo, significando que a população está aumentando de tamanho (a taxa de variação é positiva); ou negativo, significando que a população está diminuindo de tamanho; ou zero, caso em que o tamanho da população é imutável, uma condição conhecida como crescimento populacional zero.
Crescimento logístico
O crescimento exponencial estendido só é possível quando recursos naturais infinitos estão disponíveis; esse não é o caso no mundo real. Charles Darwin reconheceu esse fato em sua descrição da “luta pela existência”, que afirma que os indivíduos competirão (com membros de sua própria espécie ou de outra espécie) por recursos limitados. Os bem-sucedidos têm maior probabilidade de sobreviver e transmitir as características que os tornaram bem-sucedidos para a próxima geração em um ritmo maior (seleção natural). Para modelar a realidade dos recursos limitados, os ecologistas populacionais desenvolveram o modelo de crescimento logístico.
Capacidade de carga e o modelo logístico
No mundo real, com seus recursos limitados, o crescimento exponencial não pode continuar indefinidamente. O crescimento exponencial pode ocorrer em ambientes onde há poucos indivíduos e recursos abundantes, mas quando o número de indivíduos fica grande o suficiente, os recursos se esgotam e a taxa de crescimento diminui. Eventualmente, a taxa de crescimento se estabilizará ou se estabilizará (Figura\(\PageIndex{1}\) b). Esse tamanho populacional, que é determinado pelo tamanho máximo da população que um ambiente específico pode sustentar, é chamado de capacidade de carga, ou K. Em populações reais, uma população crescente geralmente ultrapassa sua capacidade de carga, e a taxa de mortalidade aumenta além da taxa de natalidade, fazendo com que o tamanho da população diminua de volta à capacidade de carga ou abaixo dela. A maioria das populações geralmente flutua em torno da capacidade de carga de forma ondulada, em vez de existir diretamente nela.
A fórmula usada para calcular o crescimento logístico adiciona a capacidade de carga como uma força moderadora na taxa de crescimento. A expressão “K — N” é igual ao número de indivíduos que podem ser adicionados a uma população em um determinado momento, e “K — N” dividido por “K” é a fração da capacidade de carga disponível para um maior crescimento. Assim, o modelo de crescimento exponencial é restringido por esse fator para gerar a equação de crescimento logístico:
\[\text{Population growth} = rN \left[\dfrac{K-N}{K}\right] \nonumber\]
Observe que quando N é quase zero, a quantidade entre colchetes é quase igual a 1 (ou K/K) e o crescimento é próximo do exponencial. Quando o tamanho da população é igual à capacidade de carga, ou N = K, a quantidade entre colchetes é igual a zero e o crescimento é igual a zero. Um gráfico dessa equação (crescimento logístico) produz a curva em forma de S (Figura\(\PageIndex{1}\) b). É um modelo mais realista de crescimento populacional do que crescimento exponencial. Há três seções diferentes em uma curva em forma de S. Inicialmente, o crescimento é exponencial porque há poucos indivíduos e amplos recursos disponíveis. Então, à medida que os recursos começam a ficar limitados, a taxa de crescimento diminui. Finalmente, a taxa de crescimento se estabiliza na capacidade de carga do meio ambiente, com pouca mudança no número da população ao longo do tempo.
Papel da competição intraespecífica
O modelo logístico pressupõe que todo indivíduo dentro de uma população terá igual acesso aos recursos e, portanto, uma chance igual de sobrevivência. Para as plantas, a quantidade de água, luz solar, nutrientes e espaço para crescer são os recursos importantes, enquanto nos animais, recursos importantes incluem comida, água, abrigo, espaço de nidificação e parceiros.
No mundo real, a variação fenotípica entre indivíduos dentro de uma população significa que alguns indivíduos estarão melhor adaptados ao ambiente do que outros. A competição resultante por recursos entre membros da população da mesma espécie é denominada competição intraespecífica. A competição intraespecífica pode não afetar populações que estão bem abaixo de sua capacidade de carga, pois os recursos são abundantes e todos os indivíduos podem obter o que precisam. No entanto, à medida que o tamanho da população aumenta, essa competição se intensifica. Além disso, o acúmulo de resíduos pode reduzir a capacidade de carga em um ambiente.
Exemplos de crescimento logístico
O fermento, um fungo microscópico usado para fazer pão e bebidas alcoólicas, exibe a curva clássica em forma de S quando cultivada em um tubo de ensaio (Figura\(\PageIndex{2}\) a). Seu crescimento diminui à medida que a população esgota os nutrientes necessários para seu crescimento. No mundo real, no entanto, existem variações nessa curva idealizada. Exemplos em populações selvagens incluem ovelhas e focas (Figura\(\PageIndex{2}\) b). Em ambos os exemplos, o tamanho da população excede a capacidade de carga por curtos períodos de tempo e depois cai abaixo da capacidade de carga. Essa flutuação no tamanho da população continua ocorrendo à medida que a população oscila em torno de sua capacidade de carga. Ainda assim, mesmo com essa oscilação, o modelo logístico está confirmado.
CONEXÃO ARTÍSTICA
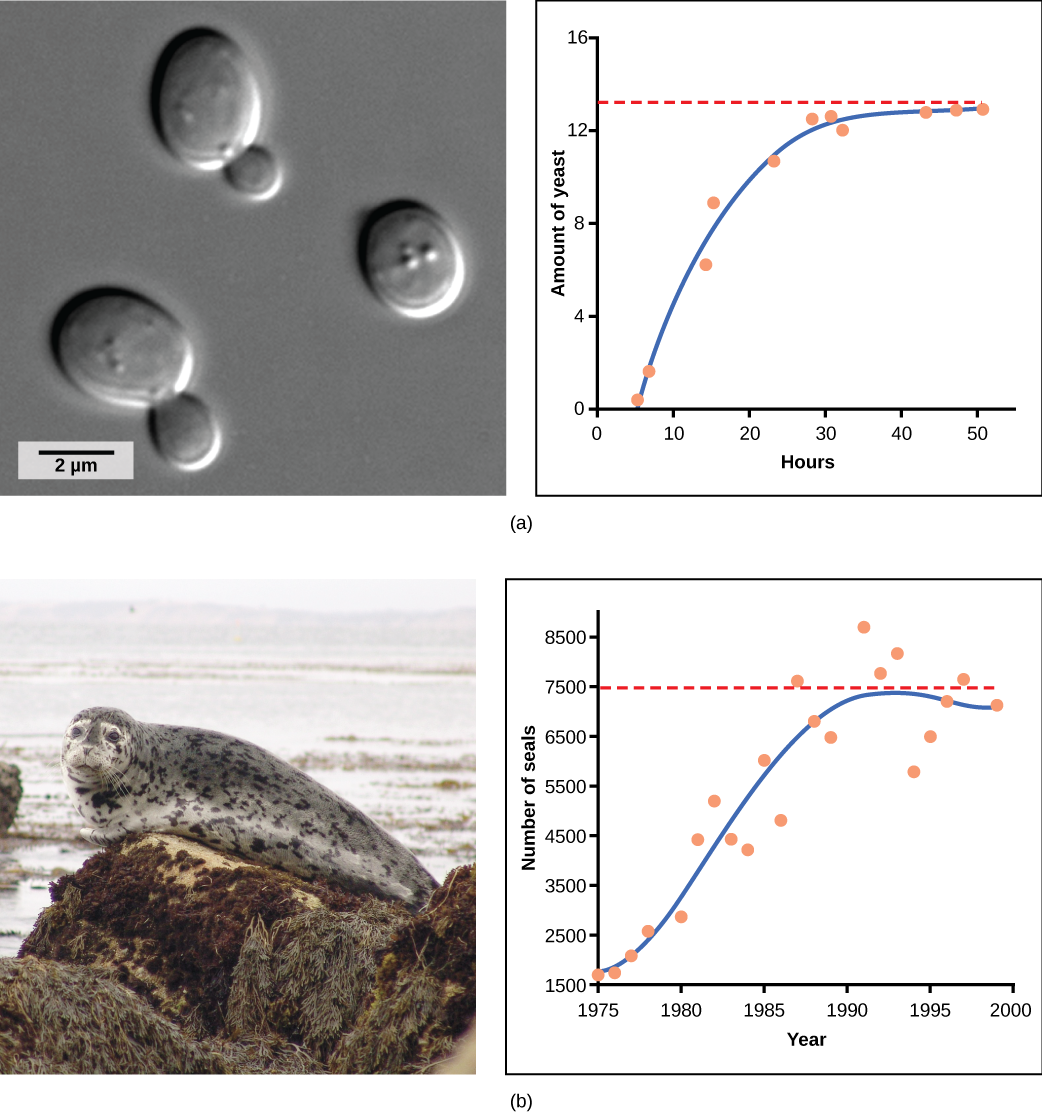
Se a principal fonte alimentar de focas diminuir devido à poluição ou à sobrepesca, qual das seguintes opções provavelmente ocorreria?
- A capacidade de carga das focas diminuiria, assim como a população de focas.
- A capacidade de carga das focas diminuiria, mas a população de focas permaneceria a mesma.
- O número de mortes por focas aumentaria, mas o número de nascimentos também aumentaria, então o tamanho da população permaneceria o mesmo.
- A capacidade de carga das focas permaneceria a mesma, mas a população de focas diminuiria.
Dinâmica e regulação populacional
O modelo logístico de crescimento populacional, embora válido em muitas populações naturais e seja um modelo útil, é uma simplificação da dinâmica populacional do mundo real. Implícito no modelo está que a capacidade de carga do ambiente não muda, o que não é o caso. A capacidade de carga varia anualmente. Por exemplo, alguns verões são quentes e secos, enquanto outros são frios e úmidos; em muitas áreas, a capacidade de carga durante o inverno é muito menor do que no verão. Além disso, eventos naturais como terremotos, vulcões e incêndios podem alterar o ambiente e, portanto, sua capacidade de carga. Além disso, as populações geralmente não existem de forma isolada. Eles compartilham o ambiente com outras espécies, competindo com elas pelos mesmos recursos (competição interespecífica). Esses fatores também são importantes para entender como uma população específica crescerá.
O crescimento populacional é regulado de várias maneiras. Eles são agrupados em fatores dependentes da densidade, nos quais a densidade da população afeta a taxa de crescimento e a mortalidade, e fatores independentes da densidade, que causam mortalidade em uma população independentemente da densidade populacional. Os biólogos da vida selvagem, em particular, querem entender os dois tipos porque isso os ajuda a gerenciar populações e evitar a extinção ou a superpopulação.
Regulação dependente da densidade
A maioria dos fatores dependentes da densidade são de natureza biológica e incluem predação, competição inter e intraespecífica e parasitas. Normalmente, quanto mais densa for a população, maior será sua taxa de mortalidade. Por exemplo, durante a competição intra e interespecífica, as taxas reprodutivas da espécie geralmente serão menores, reduzindo a taxa de crescimento de suas populações. Além disso, a baixa densidade de presas aumenta a mortalidade de seu predador porque ele tem mais dificuldade em localizar sua fonte de alimento. Além disso, quando a população é mais densa, as doenças se espalham mais rapidamente entre os membros da população, o que afeta a taxa de mortalidade.
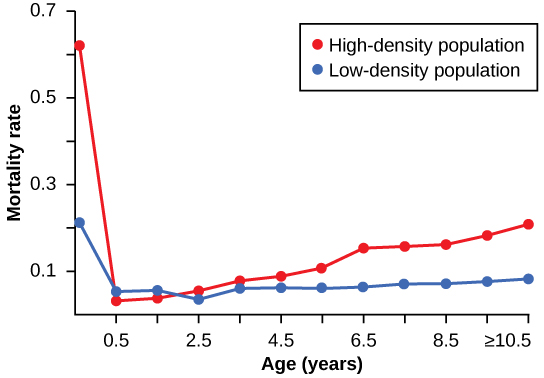
A regulação dependente da densidade foi estudada em um experimento natural com populações de burros selvagens em dois locais na Austrália. 1 Em um local, a população foi reduzida por um programa de controle populacional; a população no outro local não recebeu nenhuma interferência. A parcela de alta densidade era duas vezes mais densa que a parcela de baixa densidade. De 1986 a 1987, a parcela de alta densidade não viu nenhuma mudança na densidade de burros, enquanto a parcela de baixa densidade viu um aumento na densidade de burros. A diferença nas taxas de crescimento das duas populações foi causada pela mortalidade, não por uma diferença nas taxas de natalidade. Os pesquisadores descobriram que o número de filhos nascidos por cada mãe não foi afetado pela densidade. As taxas de crescimento nas duas populações foram diferentes, principalmente por causa da mortalidade juvenil causada pela desnutrição da mãe devido à escassez de alimentos de alta qualidade na população densa. A figura\(\PageIndex{3}\) mostra a diferença nas mortalidades específicas por idade nas duas populações.
Regulação independente da densidade e interação com fatores dependentes da densidade
Muitos fatores de natureza tipicamente física causam a mortalidade de uma população, independentemente de sua densidade. Esses fatores incluem clima, desastres naturais e poluição. Um cervo individual será morto em um incêndio florestal, independentemente de quantos cervos estejam nessa área. Suas chances de sobrevivência são as mesmas, independentemente de a densidade populacional ser alta ou baixa. O mesmo vale para o clima frio do inverno.
Em situações da vida real, a regulação populacional é muito complicada e fatores independentes e dependentes da densidade podem interagir. Uma população densa que sofre mortalidade por uma causa independente da densidade será capaz de se recuperar de forma diferente de uma população esparsa. Por exemplo, uma população de cervos afetada por um inverno rigoroso se recuperará mais rapidamente se houver mais cervos para se reproduzir.
EVOLUÇÃO EM AÇÃO: Por que o mamute peludo foi extinto?

Os mamutes peludos começaram a se extinguir há cerca de 10.000 anos, logo após paleontólogos acreditarem que humanos capazes de caçá-los começaram a colonizar a América do Norte e o norte da Eurásia (Figura\(\PageIndex{4}\)). Uma população gigantesca sobreviveu na Ilha Wrangel, no Mar da Sibéria Oriental, e ficou isolada do contato humano até 1700 aC. Sabemos muito sobre esses animais a partir de carcaças encontradas congeladas no gelo da Sibéria e de outras regiões do norte.
É comum pensar que as mudanças climáticas e a caça humana levaram à sua extinção. Um estudo de 2008 estimou que a mudança climática reduziu o alcance do mamute de 3.000.000 milhas quadradas 42.000 anos atrás para 310.000 milhas quadradas 6.000 anos atrás. 2 Por meio de evidências arqueológicas de locais de morte, também está bem documentado que humanos caçaram esses animais. Um estudo de 2012 concluiu que nenhum fator foi o único responsável pela extinção dessas criaturas magníficas. 3 Além das mudanças climáticas e da redução do habitat, os cientistas demonstraram que outro fator importante na extinção do mamute foi a migração de caçadores humanos pelo Estreito de Bering para a América do Norte durante a última era glacial, há 20.000 anos.
A manutenção de populações estáveis foi e é muito complexa, com muitos fatores de interação determinando o resultado. É importante lembrar que os humanos também fazem parte da natureza. Uma vez contribuímos para o declínio de uma espécie usando apenas a tecnologia de caça primitiva.
Modelos populacionais com base demográfica
Ecologistas populacionais levantaram a hipótese de que conjuntos de características podem evoluir em espécies que levam a adaptações específicas em seus ambientes. Essas adaptações impactam o tipo de crescimento populacional que suas espécies experimentam. Características da história de vida, como taxas de natalidade, idade na primeira reprodução, número de filhos e até taxas de mortalidade, evoluem da mesma forma que a anatomia ou o comportamento, levando a adaptações que afetam o crescimento populacional. Ecologistas populacionais descreveram um continuum de “estratégias” de história de vida com espécies selecionadas por K em uma extremidade e espécies selecionadas por r na outra. As espécies selecionadas por K são adaptadas a ambientes estáveis e previsíveis. Populações de espécies selecionadas por K tendem a existir perto de sua capacidade de carga. Essas espécies tendem a ter descendentes maiores, mas menores, e contribuem com grandes quantidades de recursos para cada filhote. Os elefantes seriam um exemplo de uma espécie selecionada por K. As espécies selecionadas por r são adaptadas a ambientes instáveis e imprevisíveis. Eles têm um grande número de filhos pequenos. Animais selecionados não fornecem muitos recursos ou cuidados parentais aos filhotes, e os filhotes são relativamente autossuficientes ao nascer. Exemplos de espécies selecionadas por r são invertebrados marinhos, como medusas, e plantas como o dente-de-leão. As duas estratégias extremas estão nas duas extremidades de um continuum no qual existirão histórias reais de vida de espécies. Além disso, as estratégias de história de vida não precisam evoluir como suítes, mas podem evoluir independentemente umas das outras, então cada espécie pode ter algumas características que tendem para um extremo ou outro.
Resumo da seção
Populações com recursos ilimitados crescem exponencialmente, com uma taxa de crescimento acelerada. Quando os recursos se tornam limitantes, as populações seguem uma curva de crescimento logístico na qual o tamanho da população se nivelará na capacidade de carga.
As populações são reguladas por uma variedade de fatores dependentes da densidade e independentes da densidade. Características da história de vida, como idade na primeira reprodução ou número de filhos, são características que evoluem nas populações, assim como a anatomia ou o comportamento podem evoluir ao longo do tempo. O modelo de seleção r e K sugere que caracteres, e possivelmente conjuntos de caracteres, podem evoluir adaptações à estabilidade da população perto da capacidade de carga (seleção K) ou rápido crescimento e colapso populacional (seleção r). As espécies exibirão adaptações em algum lugar em um continuum entre esses dois extremos.
Exercício art
Figura\(\PageIndex{2}\): Se a principal fonte alimentar de focas diminuir devido à poluição ou à sobrepesca, qual das seguintes opções provavelmente ocorreria?
- A capacidade de carga das focas diminuiria, assim como a população de focas.
- A capacidade de carga das focas diminuiria, mas a população de focas permaneceria a mesma.
- O número de mortes por focas aumentaria, mas o número de nascimentos também aumentaria, então o tamanho da população permaneceria o mesmo.
- A capacidade de carga das focas permaneceria a mesma, mas a população de focas diminuiria.
- Responda
-
R: A capacidade de carga das focas diminuiria, assim como a população de focas.
Notas de pé
- 1 David Choquenot, “Crescimento dependente da densidade, condição corporal e demografia em burros selvagens: testando a hipótese alimentar”, Ecology 72, nº 3 (junho de 1991) :805—813.
- 2 David Nogués-Bravo et al., “Mudanças climáticas, humanos e a extinção do mamute peludo”. PLoS Bill 6 (abril de 2008): e79, doi:10.1371/journal.pbio.0060079.
- 3 G.M. MacDonald et al., “Padrão de extinção do mamute peludo na Beringia”. Nature Communications 3, nº 893 (junho de 2012), doi:10.1038/ncomms1881.
Glossário
- taxa de natalidade
- o número de nascimentos dentro de uma população em um momento específico
- capacidade de carga
- o número máximo de indivíduos de uma população que podem ser sustentados pelos recursos limitados de um habitat
- taxa de mortalidade
- o número de mortes dentro de uma população em um momento específico
- regulação dependente da densidade
- a regulação da população na qual as taxas de natalidade e mortalidade dependem do tamanho da população
- regulação independente da densidade
- a regulação da população na qual a taxa de mortalidade é independente do tamanho da população
- crescimento exponencial
- um padrão de crescimento acelerado observado em populações onde os recursos não são limitantes
- competição intraespecífica
- a competição entre membros da mesma espécie
- curva de crescimento em forma de J
- a forma de uma curva de crescimento exponencial
- K - espécies selecionadas
- uma espécie adequada para ambientes estáveis que produzem poucos filhotes relativamente grandes e fornecem cuidados parentais
- crescimento logístico
- o nivelamento do crescimento exponencial devido à limitação de recursos
- r - espécies selecionadas
- uma espécie adequada para ambientes em mudança que produz muitos filhotes e oferece pouco ou nenhum cuidado parental
- curva de crescimento em forma de S
- a forma de uma curva de crescimento logístico
- crescimento populacional zero
- o tamanho estável da população, onde as taxas de natalidade e mortalidade são iguais