5.2: Definindo o gênero Homo
- Page ID
- 185802

Ao final desta seção, você poderá:
Colocando o Homo em contexto
Antes de aprender sobre as espécies de hominídeos que compõem a categoria gênero Homo, será útil familiarizar-se com os principais períodos arqueológicos aos quais o Homo está associado. As espécies e os desenvolvimentos culturais mencionados abaixo serão explorados com mais detalhes nas seções a seguir.
O desafio de definir o gênero Homo
O capítulo anterior apresentou os australopitecinos, que eram diversos em suas características físicas (graciosos e robustos), com mandíbulas e dentes grandes e cérebro de pequeno tamanho. Uma característica fundamental compartilhada pelos australopitecinos e pelo gênero Homo é o bipedalismo. A transição para o bipedalismo está ligada a várias mudanças anatômicas, incluindo pernas mais longas, mudanças na curvatura da coluna vertebral e o desenvolvimento de arcos nos pés para conservar energia e aumentar o equilíbrio ao caminhar.
Quais critérios além do bipedalismo podem ser usados para classificar uma espécie sob o gênero Homo? Muitos antropólogos tentaram estabelecer critérios específicos a serem usados na determinação da classificação do Homo. Os paleoantropólogos Mary Leakey, Louis Leakey e John Napier, bem como o primatologista Phillip Tobias, foram os primeiros a estudar extensivamente os fósseis do Homo habilis, considerada uma das primeiras espécies do gênero Homo. H. habilis tinha um tamanho de cérebro de cerca de 661—700 cc, que era maior que o das australopitecinas, com mãos capazes da destreza necessária para fazer ferramentas, devido a mudanças na estrutura óssea e ao reposicionamento do polegar, o que permitia uma melhor aderência.
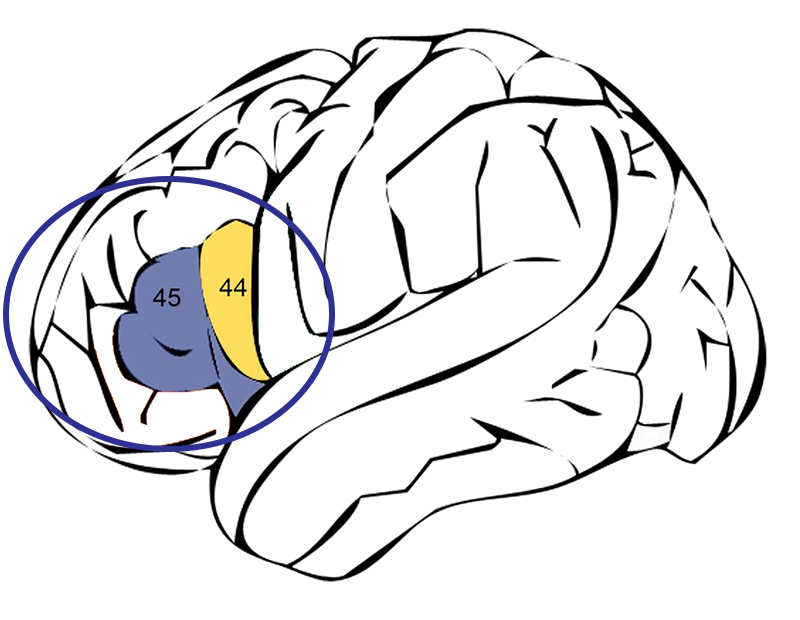
O espécime-tipo OH 7 de H. habilis datava entre 2 e 1,7 MYA e foi encontrado em 1960 no Desfiladeiro de Olduvai por Jonathan e Mary Leakey. Foi descrito por Louis Leakey em 1964. O espécime-tipo se refere a um espécime que serve como padrão para o táxon ou grupo de classificação dessa espécie. OH 7 é o número de identificação ou acesso desse espécime específico e significa “Olduvai Hominid #7”. O espécime consistia em um crânio juvenil parcial, ossos da mão e do pé. Ele possuía dentes muito menores do que os de qualquer australopitecino e possivelmente coexistia com os robustos australopitecinos (Paranthropus). Com base em um molde endocraniano (uma impressão do interior da caixa cerebral), foi determinado que H. habilis pode ter possuído o que é chamado de área de Broca no cérebro. A área de Broca, que inclui duas áreas de Brodmann (referidas como 44 e 45), está localizada no meio do córtex cerebral esquerdo do cérebro e é especialmente importante para o desenvolvimento da fala (Figura 5.2). Alguns cientistas sugeriram que o H. habilis começou a desenvolver as redes neurais necessárias para a fala humana, enquanto outros argumentam que H. habilis provavelmente já tinha a fala.
As características pós-cranianas (estruturas esqueléticas no corpo além do crânio) do Homo habilis não estão tão bem estabelecidas, como é o caso de muitos outros fósseis de hominídeos primitivos. Isso pode ser problemático, pois muitas espécies de hominídeos coexistiram com características sobrepostas. Da mesma forma, pode ser problemático ter material pós-craniano e não o crânio ou o crânio. O crânio geralmente serve como uma ferramenta de diagnóstico quando os materiais pós-cranianos não fornecem evidências suficientes ou fornecem evidências confusas.
Com base em suas pesquisas sobre H. habilis, Mary Leakey, Louis Leakey e John Napier propuseram os seguintes critérios para classificar o Homo: um tamanho cerebral acima de 600 cc; um crânio redondo e globular; uso de ferramentas; prognatismo reduzido (protrusão da mandíbula) e mandíbulas e mandíbulas menores; semelhante ao humano características pós-cranianas; e pés totalmente adaptados para caminhar (Leakey, Tobias e Napier 1964). Embora essa lista tenha estabelecido orientações específicas e bastante abrangentes, a diversidade de características e as formas pelas quais elas se sobrepunham nem sempre estavam de acordo com os critérios.
H. habilis tem estado no centro de vários debates sobre sua posição taxonômica e relação com outras espécies primitivas de Homo arcaicas. Por exemplo, acreditava-se inicialmente que H. habilis era um ancestral humano direto através da linhagem do Homo erectus e depois dos humanos modernos. Esse ponto de vista agora é debatido e resultou em uma divisão científica entre aqueles que apoiam H. habilis e aqueles que sugerem outra espécie de Homo, H. rudolfensis, como sendo a ancestral do H. erectus. H. rudolfensis é um Homo arcaico datado de cerca de 2 MYA, que coexistiu com outras espécies de Homo durante esse período. Um crânio foi descoberto em 1972 ao longo do Lago Turkana, no Quênia, por Bernard Ngeneo, um queniano local. O espécime foi posteriormente descrito pelo paleoantropólogo Richard Leakey. Muito não se sabe sobre essa espécie; faltam materiais pós-cranianos aos cientistas e, até o momento, nenhuma ferramenta foi encontrada. Há hipóteses que propõem que H. rudolfensis pode ser um H. habilis do sexo masculino, exibindo um crânio maior do que o observado em uma mulher H. habilis. Outros sugerem que é uma espécie completamente diferente. Outra polêmica gira em torno do uso de ferramentas. Embora o Homo habilis tenha sido considerado por muito tempo como o primeiro hominídeo a usar ferramentas de pedra, foi determinado, com base em evidências de marcas de corte, que pelo menos um australopitecino (A. garhi) usou ferramentas de pedra antes de H. habilis, por volta de 2,6 MYA (Semaw et al. 1997).

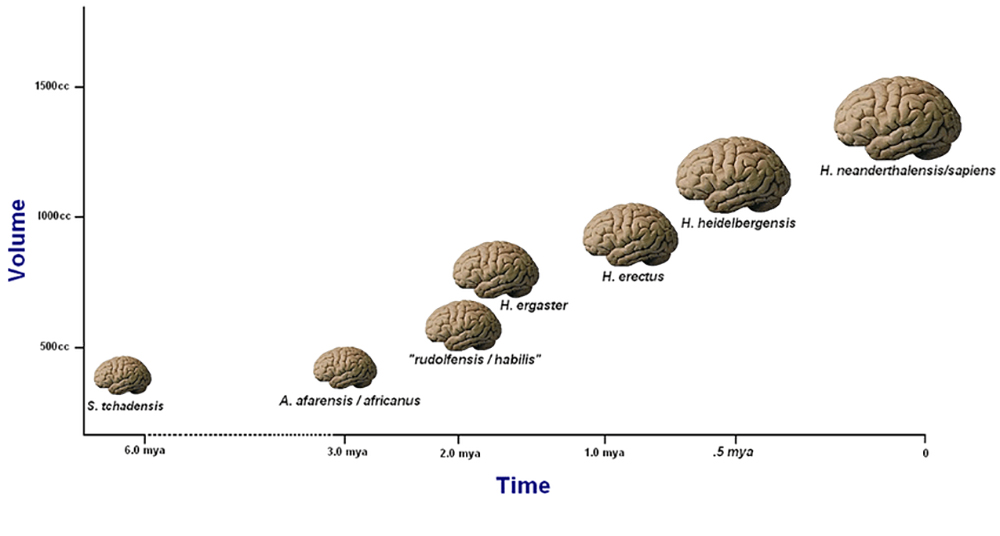
Uma das principais considerações ao classificar o H. habilis como Homo e não como Australopithecus foi sua capacidade craniana, que é uma medida que indica o tamanho do cérebro. Com algumas exceções, a capacidade craniana pode servir como um indicador de onde um fóssil de hominídeo pode pertencer à árvore filogenética dos hominídeos. A encefalização se refere a um aumento progressivo no tamanho do cérebro ao longo do tempo. Na evolução humana, podemos observar a encefalização começando com o Homo habilis e progredindo mais rapidamente através do H. erectus. A encefalização se correlaciona com um aumento na complexidade comportamental, cognitiva e cultural. Os desenvolvimentos cognitivos correspondem à nossa capacidade de construir e formar ideias, incluindo a habilidade de pensar e se comunicar por meio de linguagem simbólica e abstrata, como a usada na narrativa, no ritual e na arte. No entanto, sempre há exceções, como a H. floresiensis, que vive em uma ilha e cérebro pequeno, que será apresentada posteriormente neste capítulo. Apesar de ter um cérebro muito pequeno, H. floresiensis fabricava e usava ferramentas e construía fogueiras. Essa descoberta desafiou o que pensávamos saber sobre a correlação entre o tamanho do cérebro e o desenvolvimento cognitivo na evolução humana.
O quociente de encefalização (EQ) pode servir como um bom indicador (com algumas exceções) da classificação dentro do gênero Homo. O quociente de encefalização é um cálculo obtido pela comparação da razão entre o tamanho real do cérebro (determinado com um cálculo de massa ou volume) e o tamanho esperado do cérebro. O tamanho corporal é um fator nessas medidas, pois o tamanho esperado do cérebro reflete a relação entre o cérebro e o tamanho do corpo para um determinado grupo taxonômico (Jerison 1973). Quanto maior o peso cerebral em relação ao peso corporal geral, maior a probabilidade de o cérebro ser usado para tarefas cognitivas mais complexas. Harry J. Jerison (1973) foi o primeiro a desenvolver medições de EQ. A fórmula que ele usou para calcular o EQ em pássaros e mamíferos é massa cerebral/0,12 × (massa corporal) 0,66. Outras fórmulas também foram propostas, como EQ = massa cerebral (11,22 × massa corporal 0,76) (Martin 1981). Embora o EQ seja uma ferramenta forte para estudar o tamanho do cérebro nos primeiros hominídeos, sempre há margens potenciais de erro ao lidar com fósseis fragmentários, e formas cada vez mais alternativas de medições estão sendo propostas. Um estudo propõe que o EQ não deve mais ser usado como uma ferramenta no cálculo do tamanho do cérebro em primatas e outras espécies de vertebrados, com base na premissa de que o desempenho cognitivo não depende do tamanho do corpo e, portanto, o tamanho do corpo não deve ser incluído na fórmula (Schaik et al. 2021). Outras teorias consideram o número de neurônios corticais e conexões neurais como mais importantes quando se considera a capacidade cognitiva (Roth and Dicke 2012). De acordo com essa abordagem, a densidade do córtex está mais associada à inteligência do que ao tamanho do cérebro. Essas abordagens alternativas talvez expliquem melhor essas exceções nos registros fósseis, como H. floresiensis. Outra pesquisa interessante está analisando os níveis potenciais de cognição e memória no que se refere aos níveis de complexidade da ferramenta (Read and van der Leeuw 2008).
Apesar dessas críticas, muitos veem as medições de EQ como fornecendo resultados bastante consistentes. Os humanos modernos (Homo sapiens) têm um EQ de aproximadamente 6,0 a 7,0 (o que significa que sua massa cerebral é seis a sete vezes maior do que se esperaria encontrar em um mamífero comparável do mesmo tamanho corporal). H. erectus tem um EQ de 4,0, enquanto para um australopitecino o EQ é de cerca de 2,5 a 3,0 (Fuente 2012, 227). A Figura 5.4 mostra aumentos no tamanho médio do cérebro de várias espécies ao longo do tempo.