
3.3 : Caractéristiques uniques des cellules procaryotes
- Page ID
- 188023

Objectifs d'apprentissage
- Expliquer les caractéristiques distinctives des cellules procaryotes
- Décrire les morphologies cellulaires communes et les arrangements cellulaires typiques des cellules procaryotes et expliquer comment les cellules conservent leur morphologie
- Décrire les structures internes et externes des cellules procaryotes en termes de structure physique, de structure chimique et de fonction
- Comparez les caractéristiques distinctives des cellules bactériennes et archéologiques
La théorie cellulaire affirme que la cellule est l'unité fondamentale de la vie. Cependant, la taille, la forme, la structure et la fonction des cellules varient considérablement. Au niveau de construction le plus simple, toutes les cellules possèdent quelques composants fondamentaux. Il s'agit notamment du cytoplasme (une substance semblable à un gel composée d'eau et de substances chimiques dissoutes nécessaires à la croissance), contenu dans une membrane plasmique (également appelée membrane cellulaire ou membrane cytoplasmique) ; un ou plusieurs chromosomes, qui contiennent les plans génétiques de la cellule ; et les ribosomes, les organites utilisés pour la production de protéines.
Au-delà de ces composants de base, les cellules peuvent varier considérablement d'un organisme à l'autre, et même au sein d'un même organisme multicellulaire. Les deux plus grandes catégories de cellules, les cellules procaryotes et les cellules eucaryotes, sont définies par des différences majeures dans plusieurs structures cellulaires. Les cellules procaryotes n'ont pas de noyau entouré d'une membrane nucléaire complexe et possèdent généralement un seul chromosome circulaire situé dans un nucléoïde. Les cellules eucaryotes ont un noyau entouré d'une membrane nucléaire complexe qui contient de multiples chromosomes en forme de bâtonnets. 1
Toutes les cellules végétales et animales sont eucaryotes. Certains microorganismes sont composés de cellules procaryotes, tandis que d'autres sont composés de cellules eucaryotes. Les microorganismes procaryotes sont classés dans les domaines Archaea et Bactéries, tandis que les organismes eucaryotes sont classés dans le domaine Eucarya.
Les structures à l'intérieur d'une cellule sont analogues aux organes du corps humain, avec des structures uniques adaptées à des fonctions spécifiques. Certaines structures présentes dans les cellules procaryotes sont similaires à celles que l'on trouve dans certaines cellules eucaryotes ; d'autres sont propres aux procaryotes. À quelques exceptions près, les cellules eucaryotes ont tendance à être plus grosses que les cellules procaryotes. La taille relativement plus grande des cellules eucaryotes impose de compartimenter divers processus chimiques au sein de différentes zones de la cellule, à l'aide d'organites complexes liés à la membrane. En revanche, les cellules procaryotes sont généralement dépourvues d'organites liés à la membrane ; cependant, elles contiennent souvent des inclusions qui compartimentent leur cytoplasme. La figure\(\PageIndex{1}\) illustre les structures généralement associées aux cellules procaryotes. Ces structures sont décrites plus en détail dans la section suivante.
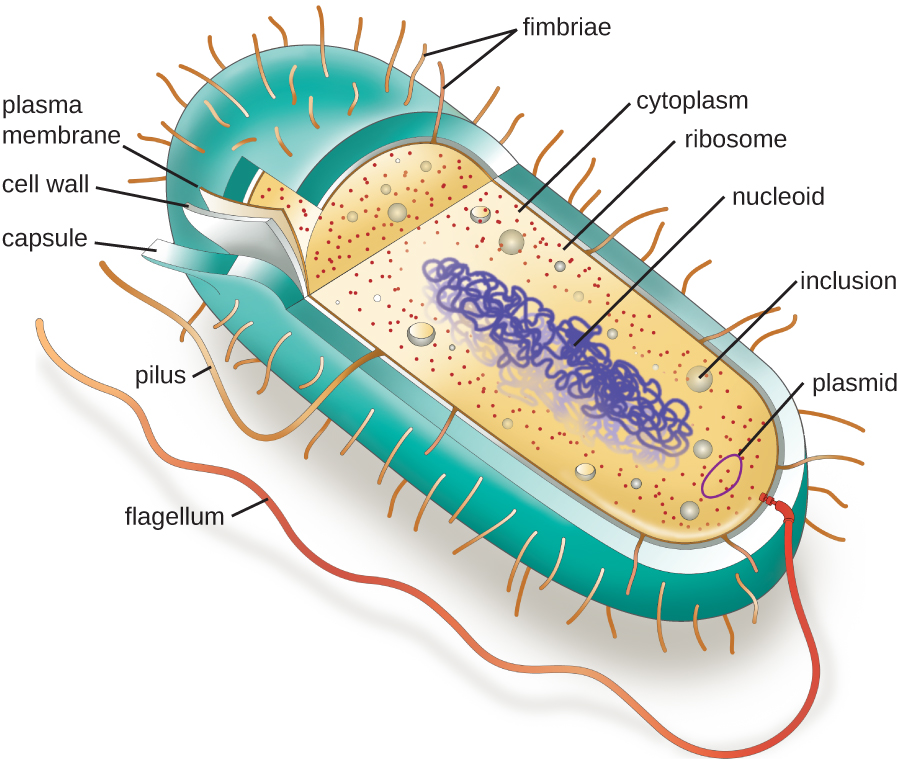
Morphologies et arrangements cellulaires courants
Les cellules individuelles d'un organisme procaryote particulier ont généralement une forme ou une morphologie cellulaire similaires. Bien que des milliers d'organismes procaryotes aient été identifiés, seule une poignée de morphologies cellulaires sont couramment observées au microscope. La figure\(\PageIndex{2}\) nomme et illustre les morphologies cellulaires que l'on trouve couramment dans les cellules procaryotes. Outre leur forme cellulaire, les cellules procaryotes d'une même espèce peuvent se regrouper selon certains arrangements distincts en fonction du plan de division cellulaire. Certains arrangements courants sont illustrés dans la figure\(\PageIndex{3}\).
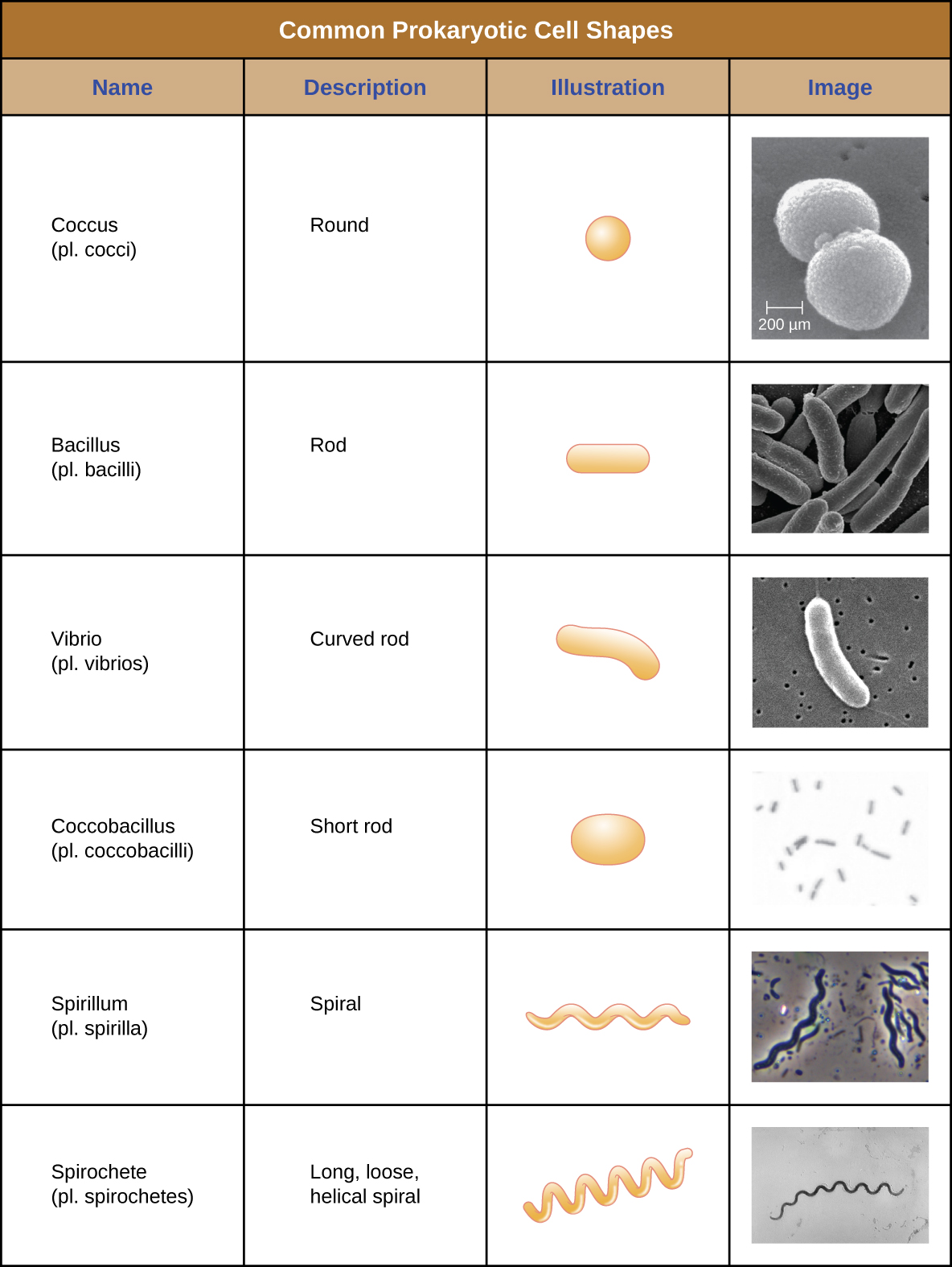
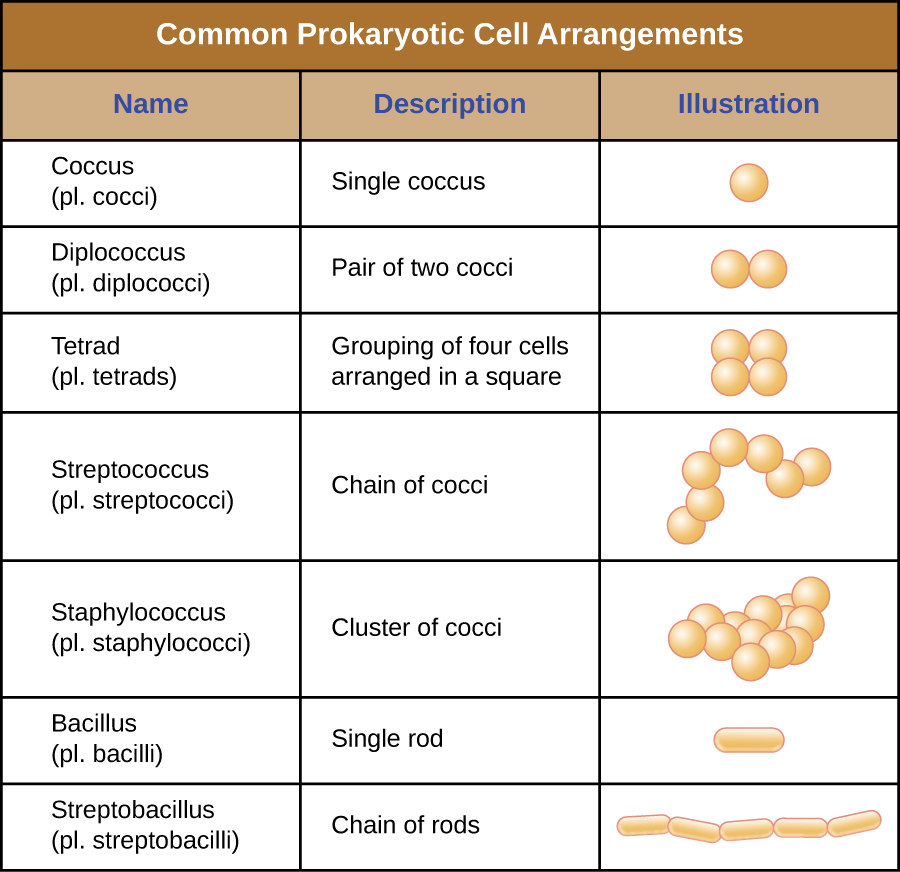
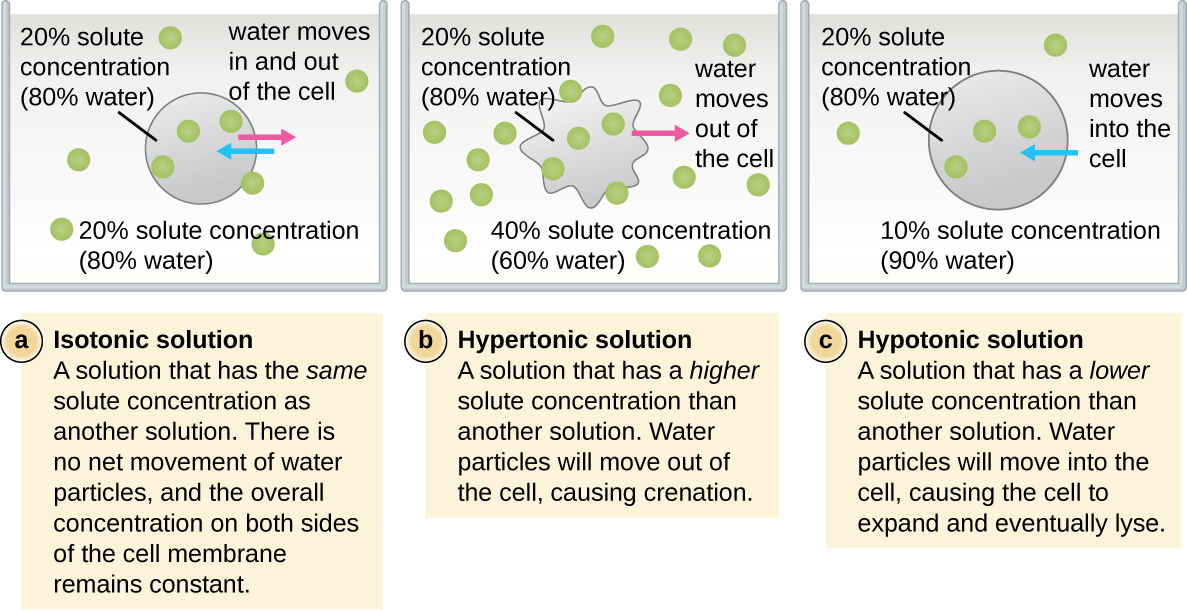
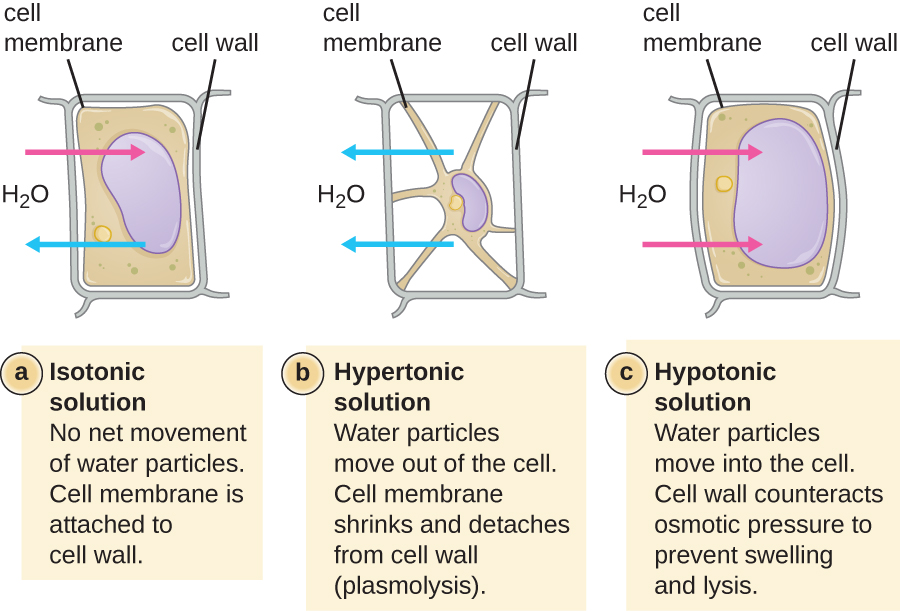
Dans la plupart des cellules procaryotes, la morphologie est maintenue par la paroi cellulaire en combinaison avec des éléments du cytosquelette. La paroi cellulaire est une structure que l'on retrouve chez la plupart des procaryotes et chez certains eucaryotes ; elle enveloppe la membrane cellulaire, protégeant ainsi la cellule des variations de pression osmotique (Figure\(\PageIndex{4}\)). La pression osmotique se produit en raison des différences de concentration des solutés sur les côtés opposés d'une membrane semi-perméable. L'eau peut traverser une membrane semi-perméable, mais les solutés (molécules dissoutes comme les sels, les sucres et autres composés) ne le peuvent pas. Lorsque la concentration de solutés est plus élevée d'un côté de la membrane, l'eau diffuse à travers la membrane du côté où la concentration est la plus faible (plus d'eau) vers le côté où la concentration est la plus élevée (moins d'eau) jusqu'à ce que les concentrations des deux côtés deviennent égales. Cette diffusion de l'eau est appelée osmose et peut provoquer une pression osmotique extrême sur une cellule lorsque son environnement extérieur change.
L'environnement externe d'une cellule peut être décrit comme un milieu isotonique, hypertonique ou hypotonique. Dans un milieu isotonique, les concentrations de soluté à l'intérieur et à l'extérieur de la cellule sont approximativement égales, de sorte qu'il n'y a pas de mouvement net de l'eau à travers la membrane cellulaire. Dans un milieu hypertonique, la concentration de soluté à l'extérieur de la cellule est supérieure à celle à l'intérieur de la cellule, de sorte que l'eau diffuse hors de la cellule vers le milieu externe. Dans un milieu hypotonique, la concentration de soluté à l'intérieur de la cellule est supérieure à celle à l'extérieur de la cellule, de sorte que l'eau se déplace par osmose dans la cellule. Cela fait gonfler la cellule et potentiellement se lyser, ou éclater.
La mesure dans laquelle une cellule particulière est capable de résister aux changements de pression osmotique est appelée tonicité. Les cellules qui ont une paroi cellulaire sont mieux à même de résister aux variations subtiles de la pression osmotique et de conserver leur forme. Dans les environnements hypertoniques, les cellules dépourvues de paroi cellulaire peuvent se déshydrater, provoquant le crénage ou le ratatinage de la cellule ; la membrane plasmique se contracte et apparaît festonnée ou entaillée (Figure\(\PageIndex{4}\)). En revanche, les cellules qui possèdent une paroi cellulaire subissent une plasmolyse plutôt qu'une crénation. Lors de la plasmolyse, la membrane plasmique se contracte et se détache de la paroi cellulaire, et le volume intérieur diminue, mais la paroi cellulaire reste intacte, ce qui permet à la cellule de conserver une certaine forme et une certaine intégrité pendant un certain temps (Figure\(\PageIndex{5}\)). De même, les cellules dépourvues de paroi cellulaire sont plus sujettes à la lyse dans les environnements hypotoniques. La présence d'une paroi cellulaire permet à la cellule de conserver sa forme et son intégrité plus longtemps avant de se lyser (Figure\(\PageIndex{5}\)).
Exercice\(\PageIndex{1}\)
- Expliquez la différence entre la morphologie et la disposition des cellules.
- Quels avantages les parois cellulaires offrent-elles aux cellules procaryotes ?
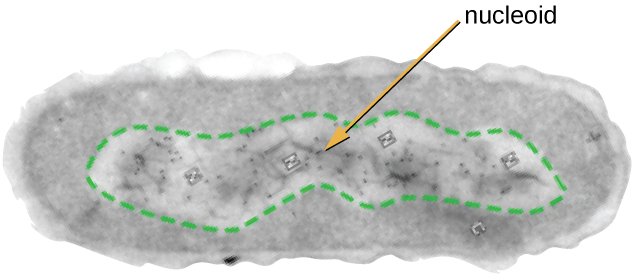
Le nucléoïde
Toute vie cellulaire possède un génome d'ADN organisé en un ou plusieurs chromosomes. Les chromosomes procaryotes sont généralement circulaires, haploïdes (non appariés) et ne sont pas liés par une membrane nucléaire complexe. L'ADN procaryote et les protéines associées à l'ADN sont concentrés dans la région nucléoïde de la cellule (Figure\(\PageIndex{6}\)). En général, l'ADN procaryote interagit avec les protéines associées aux nucléoïdes (NAPs) qui contribuent à l'organisation et au conditionnement du chromosome. Chez les bactéries, les NAPs fonctionnent de la même manière que les histones, qui sont les protéines organisatrices de l'ADN présentes dans les cellules eucaryotes. Chez les archées, le nucléoïde est organisé soit par des NAP, soit par des protéines organisatrices de l'ADN de type histone.
Plasmides
Les cellules procaryotes peuvent également contenir de l'ADN extrachromosomique ou de l'ADN ne faisant pas partie du chromosome. Cet ADN extrachromosomal se trouve dans les plasmides, qui sont de petites molécules d'ADN bicaténaires circulaires. Les cellules qui contiennent des plasmides en contiennent souvent des centaines dans une seule cellule. Les plasmides se trouvent plus fréquemment dans les bactéries, mais des plasmides ont été trouvés dans les archées et les organismes eucaryotes. Les plasmides sont souvent porteurs de gènes qui confèrent des caractéristiques avantageuses telles que la résistance aux antibiotiques ; ils sont donc importants pour la survie de l'organisme. Nous aborderons les plasmides plus en détail dans Mécanismes de génétique microbienne.
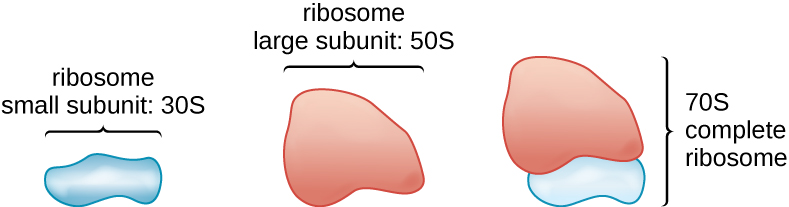
Ribosomes
Toute la vie cellulaire synthétise des protéines, et les organismes des trois domaines de la vie possèdent des ribosomes, structures responsables de la synthèse des protéines. Cependant, les ribosomes de chacun des trois domaines sont structurellement différents. Les ribosomes eux-mêmes sont construits à partir de protéines et d'ARN ribosomal (ARNr). Les ribosomes procaryotes se trouvent dans le cytoplasme. Ils sont appelés ribosomes 70S parce qu'ils ont une taille de 70S (Figure\(\PageIndex{7}\)), alors que les ribosomes cytoplasmiques eucaryotes ont une taille de 80S. (Le S représente l'unité de Svedberg, une mesure de la sédimentation dans une ultracentrifugeuse, qui est basée sur la taille, la forme et les qualités de surface de la structure analysée). Bien qu'ils soient de la même taille, les ribosomes bactériens et archéologiques contiennent des protéines et des molécules d'ARNr différentes, et les versions archées ressemblent davantage à leurs homologues eucaryotes qu'à celles trouvées chez les bactéries.
Inclusions
En tant qu'organismes unicellulaires vivant dans des environnements instables, certaines cellules procaryotes ont la capacité de stocker l'excès de nutriments dans des structures cytoplasmiques appelées inclusions. Le stockage des nutriments sous forme polymérisée est avantageux car cela réduit l'accumulation de pression osmotique qui se produit lorsqu'une cellule accumule des solutés. Différents types d'inclusions stockent du glycogène et des amidons, qui contiennent du carbone auquel les cellules peuvent accéder pour produire de l'énergie. Les granulés de volutin, également appelés granulés métachromatiques en raison de leurs caractéristiques de coloration, sont des inclusions qui stockent du phosphate inorganique polymérisé qui peut être utilisé dans le métabolisme et aider à la formation de biofilms. Les microbes connus pour contenir des granules de volutine incluent l'archée Methanosarcina, la bactérie Corynebacterium diphtheriae et l'algue eucaryote unicellulaire Chlamydomonas. Les granules de soufre, un autre type d'inclusion, se trouvent dans les bactéries soufrées du genre Thiobacillus ; ces granules stockent du soufre élémentaire, que les bactéries utilisent pour le métabolisme.
Parfois, certains types d'inclusions sont entourés d'une monocouche de phospholipides incorporée à des protéines. Le polyhydroxybutyrate (PHB), qui peut être produit par des espèces de Bacillus et de Pseudomonas, est un exemple d'inclusion présentant ce type de structure monocouche. Sur le plan industriel, le PHB a également été utilisé comme source de polymères biodégradables pour les bioplastiques. Plusieurs types d'inclusions sont illustrés sur la figure\(\PageIndex{8}\).

Certaines cellules procaryotes présentent d'autres types d'inclusions qui servent à d'autres fins que le stockage des nutriments. Par exemple, certaines cellules procaryotes produisent des vacuoles gazeuses, des accumulations de petites vésicules de gaz recouvertes de protéines. Ces vacuoles gazeuses permettent aux cellules procaryotes qui les synthétisent de modifier leur flottabilité afin qu'elles puissent ajuster leur position dans la colonne d'eau. Les bactéries magnétotactiques, telles que Magnetospirillum magnetotacticum, contiennent des magnétosomes, qui sont des inclusions d'oxyde de fer magnétique ou de sulfure de fer entourées d'une couche lipidique. Ils permettent aux cellules de s'aligner le long d'un champ magnétique, facilitant ainsi leur mouvement (Figure\(\PageIndex{8}\)). Les cyanobactéries telles que Anabaena cylindrica et les bactéries telles que Halothiobacillus neapolitanus produisent des inclusions de carboxysomes. Les carboxysomes sont composés d'enveloppes extérieures de milliers de sous-unités protéiques. Leur intérieur est rempli de ribulose-1,5-bisphosphate carboxylase/oxygénase (RuBisCO) et d'anhydrase carbonique. Ces deux composés sont utilisés pour le métabolisme du carbone. Certaines cellules procaryotes possèdent également des carboxysomes qui séquestrent des enzymes fonctionnellement apparentées en un seul endroit. Ces structures sont considérées comme des proto-organites car elles compartimentent des composés ou des réactions chimiques importants, un peu comme de nombreux organites eucaryotes.
Endospores
Les cellules bactériennes sont généralement observées sous forme de cellules végétatives, mais certains genres de bactéries ont la capacité de former des endospores, des structures qui protègent essentiellement le génome bactérien à l'état dormant lorsque les conditions environnementales sont défavorables. Les endospores (à ne pas confondre avec les spores reproductrices formées par les champignons) permettent à certaines cellules bactériennes de survivre pendant de longues périodes sans nourriture ni eau, sans exposition à des produits chimiques, à des températures extrêmes et même à des radiations. Le tableau\(\PageIndex{1}\) compare les caractéristiques des cellules végétatives et des endospores.
| Cellules végétatives | Endospores |
|---|---|
| Sensible aux températures et aux radiations extrêmes | Résistant aux températures extrêmes et aux radiations |
| Gram positif | N'absorbez pas les taches de Gram, mais uniquement les taches spéciales d'endospores (voir Coloration d'échantillons microscopiques) |
| Teneur en eau et activité enzymatique normales | Déshydraté ; aucune activité métabolique |
| Capable d'une croissance et d'un métabolisme actifs | Dormant ; pas de croissance ni d'activité métabolique |
Le processus par lequel les cellules végétatives se transforment en endospores est appelé sporulation, et il commence généralement lorsque les nutriments s'épuisent ou que les conditions environnementales deviennent défavorables (Figure\(\PageIndex{9}\)). Le processus commence par la formation d'un septum dans la cellule bactérienne végétative. Le septum divise la cellule de façon asymétrique, séparant une spire d'ADN de la cellule mère. La prépore, qui formera le cœur de l'endospore, est essentiellement une copie des chromosomes de la cellule et est séparée de la cellule mère par une seconde membrane. Un cortex se forme progressivement autour de la forepore en déposant des couches de calcium et d'acide dipicolinique entre les membranes. Une enveloppe de spores protéiques se forme alors autour du cortex tandis que l'ADN de la cellule mère se désintègre. La maturation ultérieure de l'endospore se produit avec la formation d'un exosporium le plus externe. L'endospore est libérée lors de la désintégration de la cellule mère, achevant ainsi la sporulation.
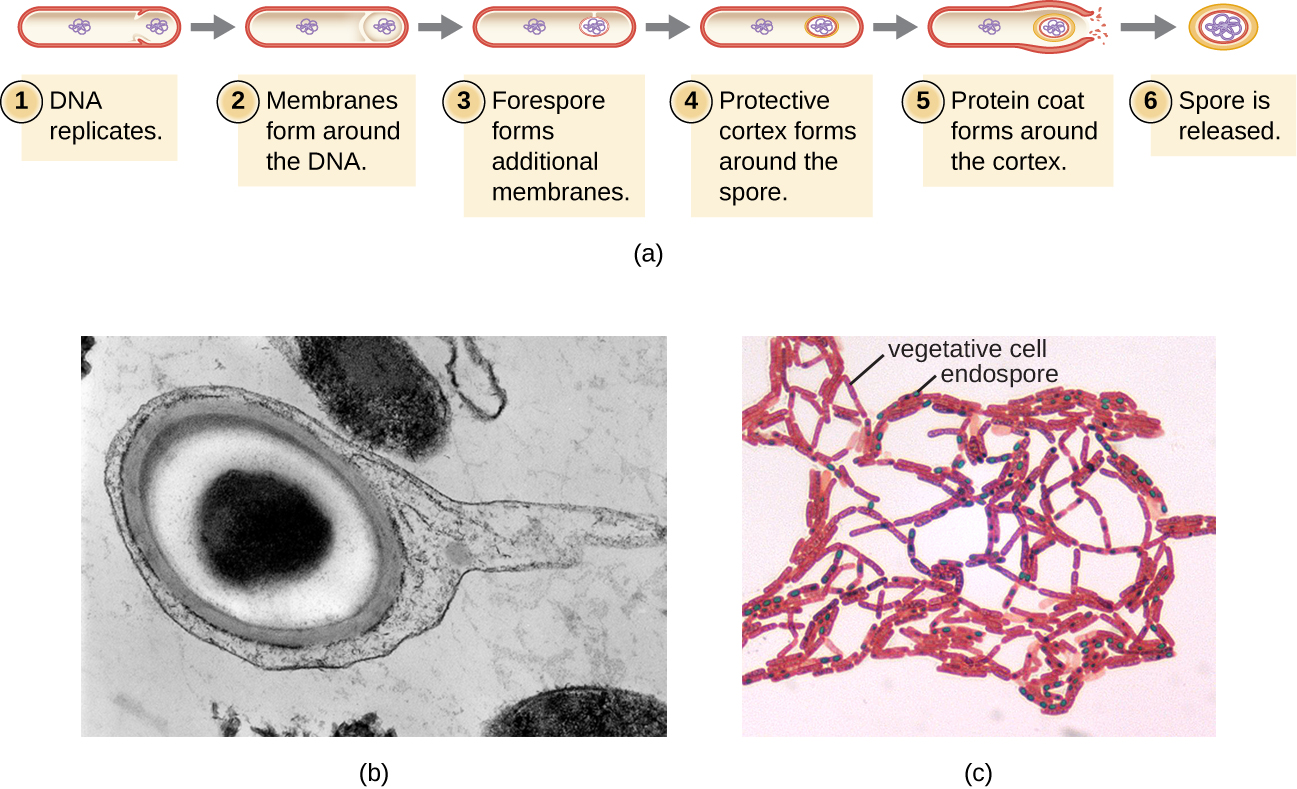
Il a été démontré que les endospores de certaines espèces persistent à l'état dormant pendant de longues périodes, allant jusqu'à des milliers d'années. 2 Cependant, lorsque les conditions de vie s'améliorent, les endospores germent et retournent à l'état végétatif. Après la germination, la cellule redevient métaboliquement active et est capable de remplir toutes ses fonctions normales, y compris la croissance et la division cellulaire.
Toutes les bactéries ne sont pas capables de former des endospores ; toutefois, il existe un certain nombre de bactéries gram-positives cliniquement significatives des genres Bacillus et Clostridium qui forment des endospores. Il s'agit notamment de B. anthracis, l'agent responsable de l'anthrax, qui produit des endospores capables de survivre pendant de nombreuses décennies 3 ; C. tetani (cause le tétanos) ; C. difficile (cause la colite pseudomembraneuse) ; C. perfringens (cause la gangrène gazeuse) ; et C. botulinum (cause le botulisme). De tels agents pathogènes sont particulièrement difficiles à combattre car leurs endospores sont si difficiles à tuer. Des méthodes de stérilisation spéciales pour les bactéries formant des endospores sont abordées dans la section Contrôle de la croissance microbienne.
Exercice\(\PageIndex{2}\)
- Qu'est-ce qu'une inclusion ?
- Quelle est la fonction d'une endospore ?
Membrane plasmique
Les structures qui entourent le cytoplasme et les structures internes de la cellule sont appelées collectivement enveloppe cellulaire. Dans les cellules procaryotes, les structures de l'enveloppe cellulaire varient en fonction du type de cellule et d'organisme. La plupart des cellules procaryotes (mais pas toutes) possèdent une paroi cellulaire, mais la composition de cette paroi varie. Toutes les cellules (procaryotes et eucaryotes) possèdent une membrane plasmique (également appelée membrane cytoplasmique ou membrane cellulaire) qui présente une perméabilité sélective, permettant à certaines molécules d'entrer ou de sortir de la cellule tout en limitant le passage d'autres.
La structure de la membrane plasmique est souvent décrite en termes de modèle de mosaïque fluide, qui fait référence à la capacité des composants de la membrane à se déplacer de manière fluide dans le plan de la membrane, ainsi qu'à la composition en mosaïque des composants, qui comprennent un large éventail de composants lipidiques et protéiques ( \(\PageIndex{10}\)Chiffre). La structure de la membrane plasmique de la plupart des types de cellules bactériennes et eucaryotes est une bicouche composée principalement de phospholipides formés par des liaisons ester et des protéines. Ces phospholipides et protéines ont la capacité de se déplacer latéralement dans le plan des membranes ainsi qu'entre les deux couches de phospholipides.

Les membranes archéologiques sont fondamentalement différentes des membranes bactériennes et eucaryotes à bien des égards. Tout d'abord, les phospholipides des membranes archéales se forment avec des liaisons éthers, contrairement aux liaisons ester présentes dans les membranes des cellules bactériennes ou eucaryotes. Deuxièmement, les phospholipides archéaux possèdent des chaînes ramifiées, tandis que celles des cellules bactériennes et eucaryotes sont à chaîne droite. Enfin, bien que certaines membranes archéales puissent être formées de bicouches comme celles que l'on trouve chez les bactéries et les eucaryotes, d'autres membranes plasmiques archéales sont des monocouches lipidiques.
Les protéines présentes à la surface de la cellule sont importantes pour diverses fonctions, notamment la communication intercellulaire et la détection des conditions environnementales et des facteurs de virulence pathogènes. Les protéines membranaires et les phospholipides peuvent être associés à des glucides (sucres) et sont appelés glycoprotéines ou glycolipides, respectivement. Ces complexes de glycoprotéines et de glycolipides s'étendent à partir de la surface de la cellule, permettant à celle-ci d'interagir avec l'environnement extérieur (Figure\(\PageIndex{10}\)). La composition chimique des glycoprotéines et des glycolipides de la membrane plasmique peut varier considérablement entre les archées, les bactéries et les eucaryotes, ce qui permet aux scientifiques de les utiliser pour caractériser des espèces uniques.
Les membranes plasmiques de différents types de cellules contiennent également des phospholipides uniques, qui contiennent des acides gras. Comme décrit dans Utilisation de la biochimie pour identifier les microorganismes, les profils d'analyse des acides gras dérivés des phospholipides (PLFA) peuvent être utilisés pour identifier des types uniques de cellules en fonction des différences d'acides gras. Les archées, les bactéries et les eucaryotes ont chacun un profil PFLA unique.
Mécanismes de transport membran
L'une des fonctions les plus importantes de la membrane plasmique est de contrôler le transport des molécules à l'intérieur et à l'extérieur de la cellule. Les conditions internes doivent être maintenues dans une certaine plage malgré toute modification de l'environnement extérieur. Le transport de substances à travers la membrane plasmique permet aux cellules de le faire.
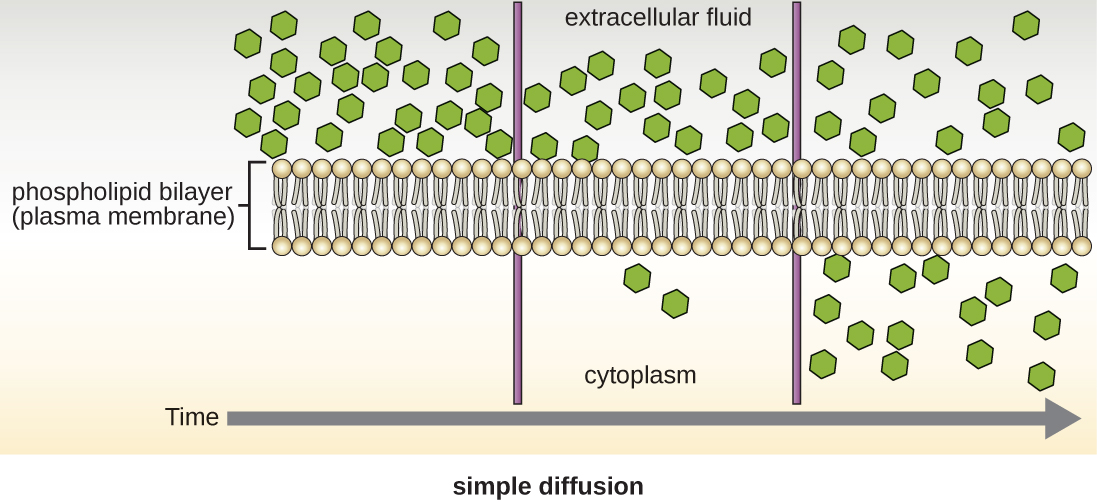
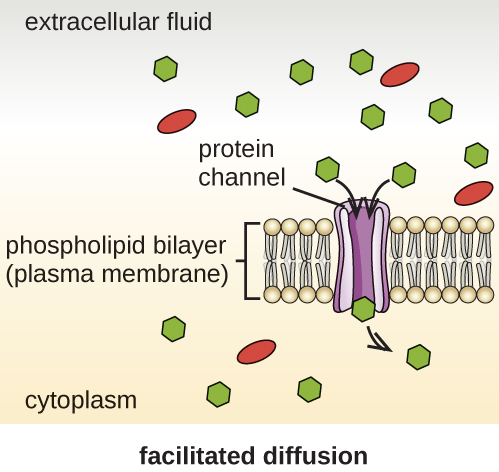
Les cellules utilisent différents modes de transport à travers la membrane plasmique. Par exemple, les molécules passant d'une concentration plus élevée à une concentration plus faible avec le gradient de concentration sont transportées par simple diffusion, également appelée transport passif (Figure\(\PageIndex{11}\)). Certaines petites molécules, comme le dioxyde de carbone, peuvent traverser la bicouche membranaire directement par simple diffusion. Cependant, les molécules chargées, ainsi que les grosses molécules, ont besoin de l'aide de supports ou de canaux dans la membrane. Ces structures transportent les molécules à travers la membrane, un processus connu sous le nom de diffusion facilitée (Figure\(\PageIndex{12}\)).
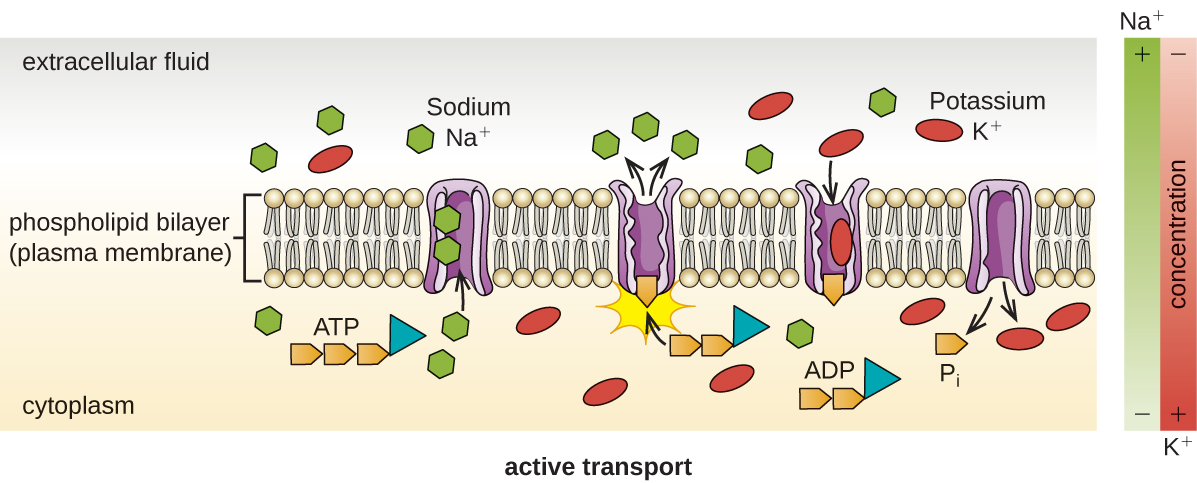
Le transport actif se produit lorsque les cellules déplacent des molécules à travers leur membrane contre des gradients de concentration (Figure\(\PageIndex{13}\)). L'une des principales différences entre le transport passif et le transport actif est que le transport actif nécessite de l'adénosine triphosphate (ATP) ou d'autres formes d'énergie pour déplacer les molécules « vers le haut ». Par conséquent, les structures de transport actives sont souvent appelées « pompes ».
La translocation de groupe transporte également des substances dans les cellules bactériennes. Dans ce cas, lorsqu'une molécule entre dans une cellule à l'encontre de son gradient de concentration, elle est modifiée chimiquement de telle sorte qu'elle n'a pas besoin d'être transportée contre un gradient de concentration défavorable. Un exemple courant de ce phénomène est le système bactérien de phosphotransférase, une série de supports qui phosphorylent (c'est-à-dire ajoutent des ions phosphate au) glucose ou à d'autres sucres lorsqu'ils pénètrent dans les cellules. Comme la phosphorylation des sucres est nécessaire aux premiers stades du métabolisme des sucres, le système de phosphotransférase est considéré comme un système neutre en énergie.
Structures de membranes photosynthétiques
Certaines cellules procaryotes, notamment les cyanobactéries et les bactéries photosynthétiques, possèdent des structures membranaires qui leur permettent de réaliser la photosynthèse. Ces structures consistent en un repliement de la membrane plasmique qui renferme des pigments photosynthétiques tels que des chlorophylles vertes et des bactériochlorophylles. Chez les cyanobactéries, ces structures membranaires sont appelées thylakoïdes ; chez les bactéries photosynthétiques, elles sont appelées chromatophores, lamelles ou chlorosomes.
Paroi cellulaire
La fonction principale de la paroi cellulaire est de protéger la cellule des conditions difficiles de l'environnement extérieur. Lorsqu'elles sont présentes, il existe des similitudes et des différences notables entre les parois cellulaires des archées, des bactéries et des eucaryotes.
Le principal composant des parois cellulaires bactériennes est le peptidoglycane (ou muréine) ; on ne le trouve que dans les bactéries. Structurellement, le peptidoglycane ressemble à une couche de maillage ou de tissu (Figure\(\PageIndex{14}\)). Chaque couche est composée de longues chaînes de molécules alternées de N-acétylglucosamine (NAG) et d'acide N-acétylmuramique (NAM). La structure des longues chaînes présente une résistance à la traction bidimensionnelle significative en raison de la formation de ponts peptidiques qui relient le NAG et le NAM au sein de chaque couche de peptidoglycane. Chez les bactéries gram-négatives, les chaînes tétrapeptidiques s'étendant à partir de chaque unité NAM sont directement réticulées, tandis que chez les bactéries gram-positives, ces chaînes tétrapeptidiques sont liées par des ponts croisés de pentaglycine. Les sous-unités du peptidoglycane sont fabriquées à l'intérieur de la cellule bactérienne, puis exportées et assemblées en couches, donnant à la cellule sa forme.
Comme le peptidoglycane est propre aux bactéries, de nombreux antibiotiques sont conçus pour interférer avec la synthèse du peptidoglycane, affaiblir la paroi cellulaire et rendre les cellules bactériennes plus sensibles aux effets de la pression osmotique (voir Mécanismes des médicaments antibactériens). De plus, certaines cellules du système immunitaire humain sont capables de « reconnaître » les bactéries pathogènes en détectant le peptidoglycane à la surface d'une cellule bactérienne ; ces cellules engloutissent et détruisent ensuite la cellule bactérienne à l'aide d'enzymes telles que le lysozyme, qui décompose et digère le peptidoglycane dans leurs parois cellulaires (voir Reconnaissance des agents pathogènes et phagocytose).
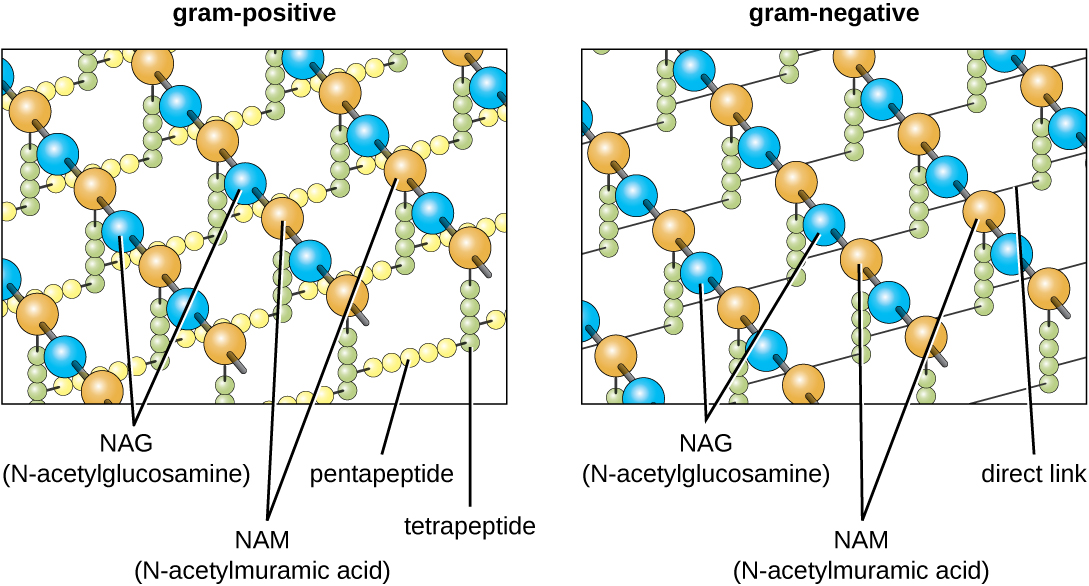
Le protocole de coloration de Gram (voir Coloration d'échantillons microscopiques) est utilisé pour différencier deux types courants de structures de parois cellulaires (Figure\(\PageIndex{15}\)). Les cellules à Gram positif ont une paroi cellulaire constituée de nombreuses couches de peptidoglycane d'une épaisseur totale de 30 à 100 nm. Ces couches de peptidoglycanes sont généralement enrobées d'acides teichoïques (TA), des chaînes de glucides qui s'étendent à travers et au-delà de la couche de peptidoglycane. On pense que le 4 TA stabilise le peptidoglycane en augmentant sa rigidité. Le TA joue également un rôle dans la capacité des bactéries gram-positives pathogènes telles que le streptocoque à se lier à certaines protéines présentes à la surface des cellules hôtes, améliorant ainsi leur capacité à provoquer des infections. En plus du peptidoglycane et du TA, les bactéries de la famille des mycobactériacées ont une couche externe d'acides mycoliques cireux dans leur paroi cellulaire ; comme décrit dans Coloration d'échantillons microscopiques, ces bactéries sont appelées acido-résistantes, car des colorants résistants aux acides doivent être utilisés pour pénétrer dans les mycoliques couche acide à des fins de microscopie (Figure\(\PageIndex{16}\)).

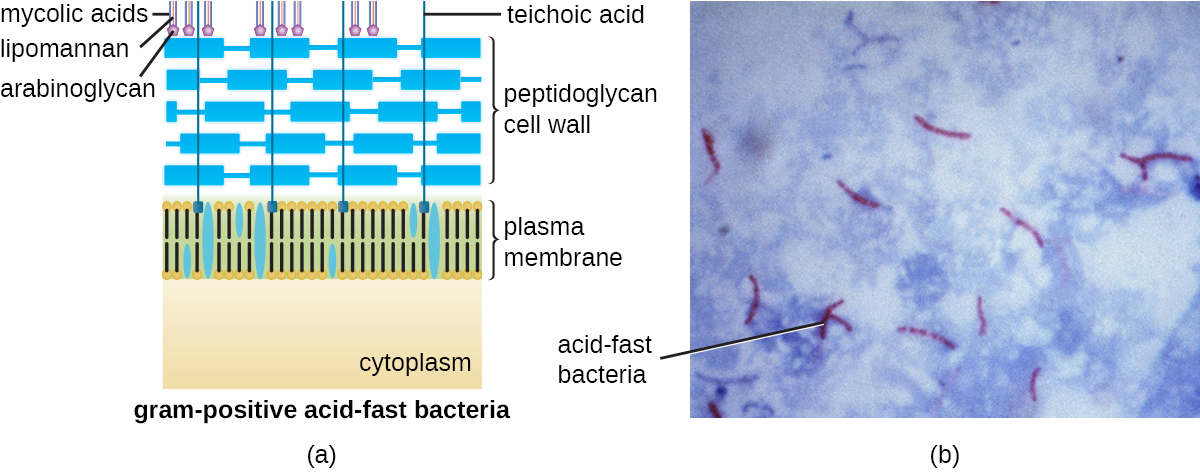
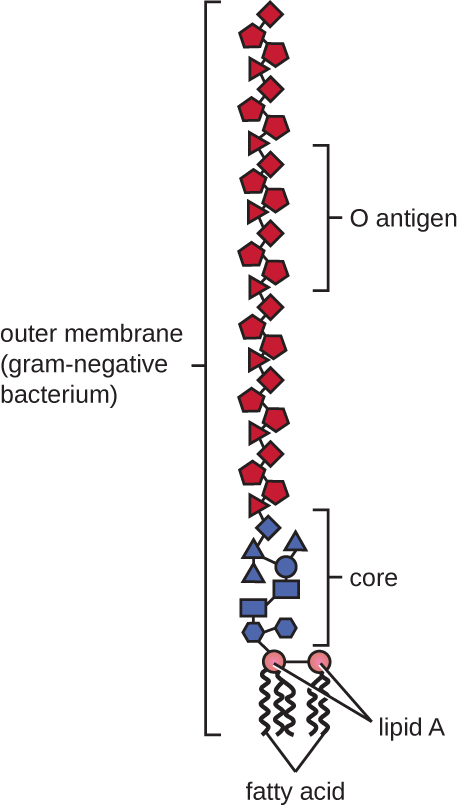
Les cellules à Gram négatif possèdent une couche de peptidoglycane beaucoup plus fine (pas plus d'environ 4 nm d'épaisseur 6) que les cellules à Gram positif, et la structure globale de leur enveloppe cellulaire est plus complexe. Dans les cellules à Gram négatif, une matrice semblable à un gel occupe l'espace périplasmique entre la paroi cellulaire et la membrane plasmique, et il existe une deuxième bicouche lipidique appelée membrane externe, qui est externe à la couche de peptidoglycane (Figure\(\PageIndex{15}\)). Cette membrane externe est fixée au peptidoglycane par la lipoprotéine muréine. Le feuillet externe de la membrane externe contient la molécule lipopolysaccharide (LPS), qui agit comme une endotoxine dans les infections impliquant des bactéries gram-négatives, contribuant à des symptômes tels que fièvre, hémorragie et choc septique. Chaque molécule de LPS est composée de lipide A, d'un polysaccharide central et d'une chaîne latérale O composée de molécules semblables au sucre qui constituent la face externe du LPS (Figure\(\PageIndex{17}\)). La composition de la chaîne latérale O varie selon les espèces et les souches de bactéries. Les parties de la chaîne latérale O appelées antigènes peuvent être détectées à l'aide de tests sérologiques ou immunologiques visant à identifier des souches pathogènes spécifiques comme Escherichia coli O157:H7, une souche mortelle de bactérie qui cause des diarrhées sanglantes et une insuffisance rénale.
La structure de la paroi cellulaire archéologique diffère de celle des bactéries de plusieurs manières importantes. Tout d'abord, les parois cellulaires archéales ne contiennent pas de peptidoglycane ; elles contiennent plutôt un polymère similaire appelé pseudopeptidoglycane (pseudomuréine) dans lequel la NAM est remplacée par une sous-unité différente. D'autres archées peuvent contenir une couche de glycoprotéines ou de polysaccharides qui sert de paroi cellulaire au lieu du pseudopeptidoglycane. Enfin, comme c'est le cas pour certaines espèces bactériennes, il existe quelques archées qui semblent totalement dépourvues de parois cellulaires.
Glycocalyces et couches S
Bien que la plupart des cellules procaryotes aient des parois cellulaires, certaines peuvent présenter des structures d'enveloppe cellulaire supplémentaires à l'extérieur de la paroi cellulaire, telles que des glycocalyces et des couches S. Un glycocalyx est une couche de sucre, qui se divise en deux types importants : les capsules et les couches visqueuses. Une capsule est une couche organisée située à l'extérieur de la paroi cellulaire et généralement composée de polysaccharides ou de protéines (Figure\(\PageIndex{18}\)). Une couche visqueuse est une couche moins bien organisée qui n'est fixée que faiblement à la paroi cellulaire et qui peut être lavée plus facilement. Les couches visqueuses peuvent être composées de polysaccharides, de glycoprotéines ou de glycolipides.
Les glycocalyces permettent aux cellules d'adhérer aux surfaces, contribuant ainsi à la formation de biofilms (colonies de microbes qui se forment en couches sur les surfaces). Dans la nature, la plupart des microbes vivent en communautés mixtes au sein de biofilms, en partie parce que le biofilm leur offre un certain niveau de protection. Les biofilms retiennent généralement l'eau comme une éponge, empêchant ainsi la dessiccation. Ils protègent également les cellules de la prédation et entravent l'action des antibiotiques et des désinfectants. Toutes ces propriétés sont avantageuses pour les microbes vivant dans un biofilm, mais elles présentent des défis en milieu clinique, où l'objectif est souvent d'éliminer les microbes.

La capacité de produire une capsule peut contribuer à la pathogénicité d'un microbe (capacité à provoquer une maladie), car la capsule peut empêcher les cellules phagocytaires (comme les globules blancs) d'engloutir et de tuer le microorganisme. Streptococcus pneumoniae, par exemple, produit une capsule bien connue pour contribuer à la pathogénicité de cette bactérie. Comme expliqué dans la section Coloration d'échantillons microscopiques, les capsules sont difficiles à colorer pour la microscopie ; des techniques de coloration négative sont généralement utilisées.
La couche S est un autre type de structure d'enveloppe cellulaire ; elle est composée d'un mélange de protéines structurales et de glycoprotéines. Chez les bactéries, les couches S se trouvent à l'extérieur de la paroi cellulaire, mais chez certaines archées, la couche S sert de paroi cellulaire. La fonction exacte des couches S n'est pas entièrement comprise et elles sont difficiles à étudier ; mais les preuves disponibles suggèrent qu'elles peuvent jouer diverses fonctions dans différentes cellules procaryotes, notamment en aidant la cellule à résister à la pression osmotique et, pour certains agents pathogènes, en interagissant avec le système immunitaire de l'hôte système.
Orientation clinique : 3e partie
Après avoir diagnostiqué une pneumonie à Barbara, le médecin légiste lui prescrit de l'amoxicilline, un type de dérivé de la pénicilline couramment prescrit. Plus d'une semaine plus tard, bien qu'elle ait suivi le cours complet comme indiqué, Barbara se sent toujours faible et n'est pas complètement rétablie, bien qu'elle soit toujours capable de mener à bien ses activités quotidiennes. Elle retourne au centre de santé pour une visite de suivi.
De nombreux types de bactéries, de champignons et de virus peuvent provoquer une pneumonie. L'amoxicilline cible le peptidoglycane des parois cellulaires bactériennes. L'amoxicilline n'ayant pas résolu les symptômes de Barbara, l'Autorité Palestinienne conclut que l'agent causal est probablement dépourvu de peptidoglycane, ce qui signifie que l'agent pathogène pourrait être un virus, un champignon ou une bactérie dépourvue de peptidoglycane. Il est également possible que l'agent pathogène soit une bactérie contenant du peptidoglycane mais qu'il ait développé une résistance à l'amoxicilline.
Exercice\(\PageIndex{3}\)
- Comment l'Autorité Palestinienne peut-elle identifier définitivement la cause de la pneumonie de Barbara ?
- Quelle forme de traitement l'AP devrait-elle prescrire, étant donné que l'amoxicilline était inefficace ?
Appendices filamenteux
De nombreuses cellules bactériennes possèdent des appendices protéiques intégrés dans leur enveloppe cellulaire qui s'étendent vers l'extérieur, permettant ainsi une interaction avec l'environnement. Ces appendices peuvent se fixer à d'autres surfaces, transférer de l'ADN ou permettre le mouvement. Les appendices filamenteux comprennent les fimbriae, les pili et les flagelles.
Fimbriae et Pili
Les fimbriae et les pili sont structurellement similaires et, comme la différenciation entre les deux est problématique, ces termes sont souvent utilisés de manière interchangeable. 7 8 Le terme fimbriae désigne généralement des protéines ressemblant à des poils courts qui se projettent par centaines de la surface des cellules. Les fimbriae permettent à une cellule de se fixer à des surfaces et à d'autres cellules. Pour les bactéries pathogènes, l'adhérence aux cellules hôtes est importante pour la colonisation, l'infectiosité et la virulence. L'adhérence aux surfaces joue également un rôle important dans la formation du biofilm.
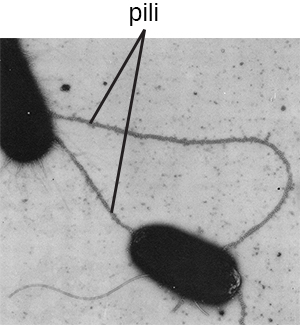
Le terme pili (singulier : pilus) désigne généralement des appendices protéiques plus longs et moins nombreux qui facilitent la fixation aux surfaces (Figure\(\PageIndex{19}\)). Un type spécifique de pilus, appelé pilus F ou pilus sexuel, joue un rôle important dans le transfert d'ADN entre cellules bactériennes, qui se produit entre des membres d'une même génération lorsque deux cellules transfèrent ou échangent physiquement des parties de leur génome respectif (voir Comment les procaryotes asexués atteignent la génétique). Diversité).
Avant que la structure et la fonction des différents composants de l'enveloppe cellulaire bactérienne ne soient bien comprises, les scientifiques utilisaient déjà les caractéristiques de l'enveloppe cellulaire pour classer les bactéries. En 1933, Rebecca Lancefield a proposé une méthode de sérotypage de diverses souches β-hémolytiques d'espèces de streptocoques à l'aide d'un test d'agglutination, une technique utilisant l'agrégation de bactéries pour détecter des antigènes spécifiques de surface cellulaire. Ce faisant, Lancefield a découvert qu'un groupe de S. pyogenes, du groupe A, était associé à diverses maladies humaines. Elle a déterminé que différentes souches de streptocoque du groupe A pouvaient être distinguées les unes des autres en fonction des variations de protéines spécifiques de surface cellulaire qu'elle a appelées protéines M.
Aujourd'hui, plus de 80 souches différentes de streptocoques du groupe A ont été identifiées sur la base des protéines M. Diverses souches du streptocoque du groupe A sont associées à une grande variété d'infections humaines, notamment la pharyngite streptococcique (angine streptococcique), l'impétigo, le syndrome de choc toxique, la scarlatine, le rhumatisme articulaire aigu et la fasciite nécrosante. La protéine M est un facteur de virulence important pour le streptocoque du groupe A, car elle aide ces souches à échapper au système immunitaire. Les modifications des protéines M semblent modifier l'infectiosité d'une souche particulière de streptocoque du groupe A.
Flagelles
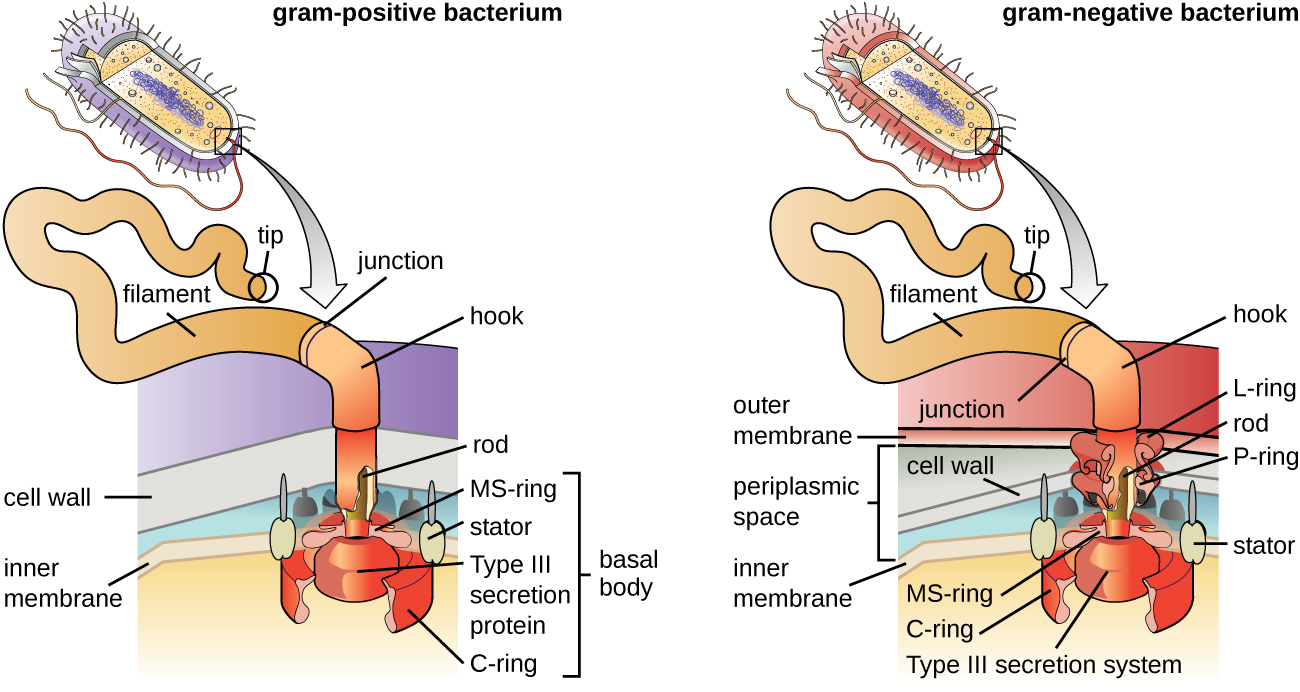
Les flagelles sont des structures utilisées par les cellules pour se déplacer dans des environnements aqueux. Les flagelles bactériens agissent comme des hélices. Ce sont des filaments spiralés rigides composés de sous-unités protéiques de la flagelline qui s'étendent vers l'extérieur à partir de la cellule et tournent en solution. Le corps basal est le moteur du flagelle et est intégré à la membrane plasmique (Figure\(\PageIndex{20}\)). Une région en crochet relie le corps basal au filament. Les bactéries à Gram positif et à Gram négatif ont des configurations basales différentes en raison des différences dans la structure des parois cellulaires.
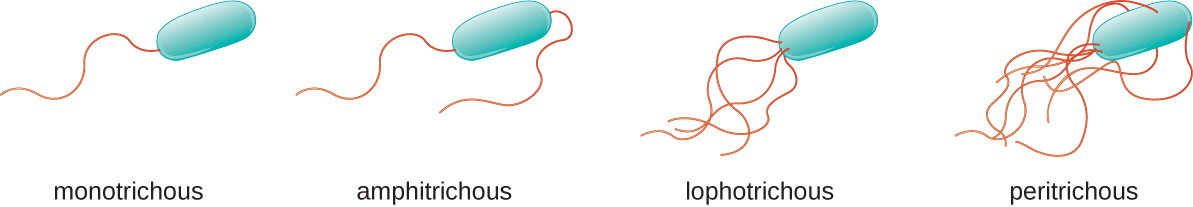
Différents types de bactéries mobiles présentent des arrangements différents de flagelles (Figure\(\PageIndex{21}\)). Une bactérie dotée d'un flagelle singulier, généralement situé à une extrémité de la cellule (polaire), est réputée avoir un flagelle monotriche. Vibrio cholerae, la bactérie à Gram négatif responsable du choléra, est un exemple de bactérie pathogène flagellée monotricheusement. Les cellules porteuses de flagelles amphitriches ont un flagelle ou des touffes de flagelles à chaque extrémité. Par exemple, Spirillum minor, cause de la fièvre spirillaire (asiatique) par morsure de rat ou sodoku. Les cellules porteuses de flagelles lophotriches possèdent une touffe à une extrémité de la cellule. Le bacille à Gram négatif Pseudomonas aeruginosa, un pathogène opportuniste connu pour provoquer de nombreuses infections, notamment des infections de « l'oreille du nageur » et des brûlures, est porteur de flagelles lophotriques. Les flagelles qui recouvrent toute la surface d'une cellule bactérienne sont appelés flagelles péritricheux. La bactérie à Gram négatif E. coli présente un arrangement péritricheux de flagelles.
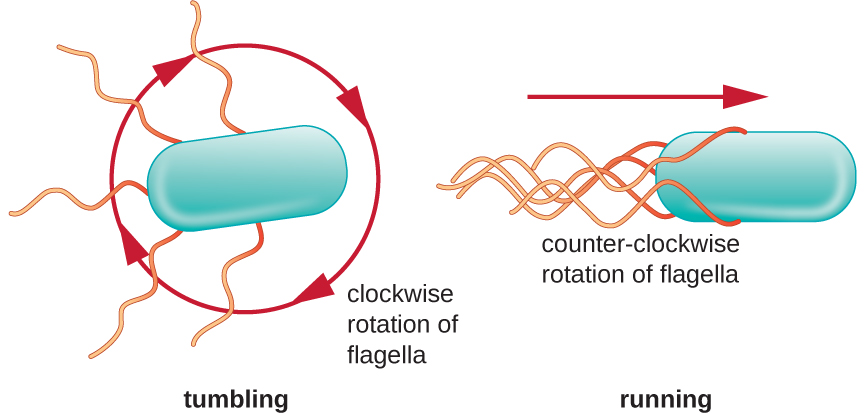
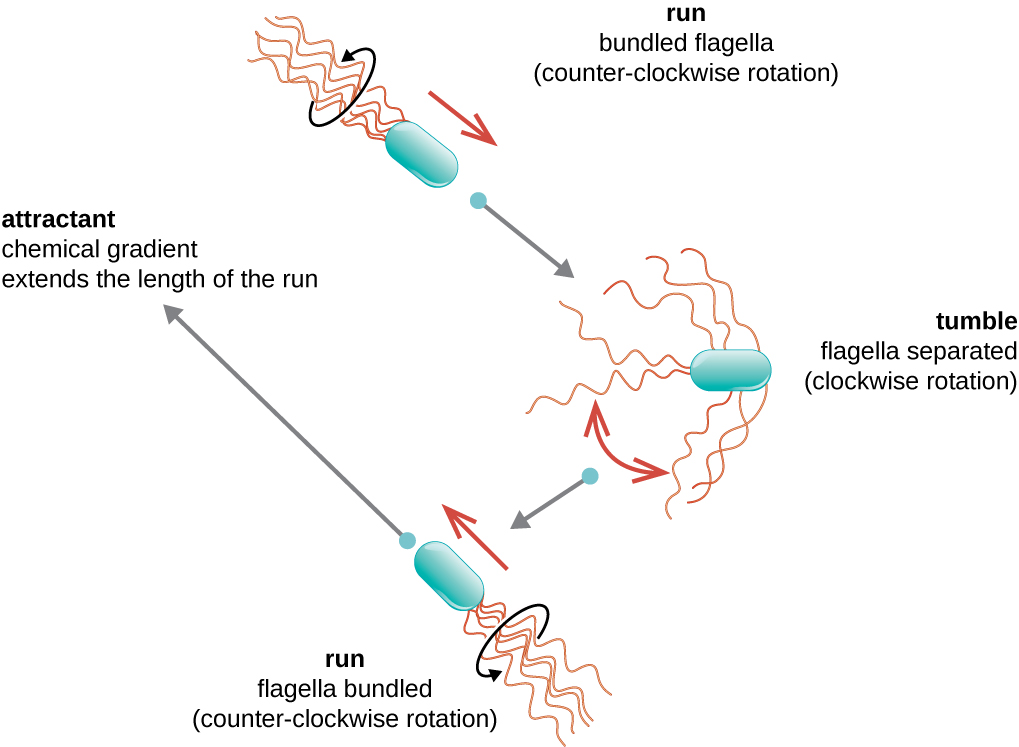
Le mouvement directionnel dépend de la configuration des flagelles. Les bactéries peuvent se déplacer en réponse à divers signaux environnementaux, notamment la lumière (phototaxie), les champs magnétiques (magnétotaxie) à l'aide de magnétosomes et, le plus souvent, les gradients chimiques (chimiotaxie). Un mouvement délibéré vers un attractif chimique, comme une source de nourriture, ou vers un répulsif, comme un produit chimique toxique, est obtenu en augmentant la durée des courses et en diminuant la longueur des culbutes. Lors de la course, les flagelles tournent dans le sens antihoraire, ce qui permet à la cellule bactérienne d'avancer. Dans une bactérie péritricheuse, les flagelles sont tous regroupés de manière très simplifiée (Figure\(\PageIndex{22}\)), ce qui permet un mouvement efficace. Lors du culbutage, les flagelles sont écartés tout en tournant dans le sens des aiguilles d'une montre, ce qui crée un mouvement en boucle et empêche tout mouvement vers l'avant significatif tout en réorientant la cellule dans la direction de l'attractif. Lorsqu'un attractif existe, des descentes et des dégringolades se produisent toujours ; toutefois, la longueur des descentes est plus longue, tandis que la longueur des culbutes est réduite, ce qui permet un mouvement global vers la concentration la plus élevée de l'attractif. En l'absence de gradient chimique, les longueurs des descentes et des culbutes sont plus égales et le mouvement global est plus aléatoire (Figure\(\PageIndex{23}\)).
Exercice\(\PageIndex{4}\)
- Qu'est-ce que la couche de peptidoglycane et en quoi diffère-t-elle entre les bactéries gram-positives et gram-négatives ?
- Comparez et opposez les flagelles monotriches, amphitriches, lophotriches et péritriches.
Résumé
- Les cellules procaryotes se distinguent des cellules eucaryotes en ce que leur matériel génétique est contenu dans un nucléoïde plutôt que dans un noyau lié à une membrane. De plus, les cellules procaryotes sont généralement dépourvues d'organites liés à la membrane.
- Les cellules procaryotes de la même espèce partagent généralement une morphologie cellulaire et un arrangement cellulaires similaires.
- La plupart des cellules procaryotes possèdent une paroi cellulaire qui aide l'organisme à maintenir sa morphologie cellulaire et le protège contre les variations de la pression osmotique.
- En dehors du nucléoïde, les cellules procaryotes peuvent contenir de l'ADN extrachromosomal dans les plasmides.
- Les ribosomes procaryotes que l'on trouve dans le cytoplasme ont une taille de 70S.
- Certaines cellules procaryotes possèdent des inclusions qui stockent des nutriments ou des produits chimiques pour d'autres utilisations.
- Certaines cellules procaryotes sont capables de former des endospores par sporulation pour survivre à l'état de dormance lorsque les conditions ne sont pas favorables. Les endospores peuvent germer et se retransformer en cellules végétatives lorsque les conditions s'améliorent.
- Dans les cellules procaryotes, l'enveloppe cellulaire comprend une membrane plasmique et généralement une paroi cellulaire.
- Les membranes bactériennes sont composées de phospholipides contenant des protéines intégrales ou périphériques. Les acides gras de ces phospholipides sont liés à des esters et sont souvent utilisés pour identifier des types spécifiques de bactéries. Les protéines remplissent diverses fonctions, notamment le transport, la communication intercellulaire et la détection des conditions environnementales. Les membranes archéales se distinguent par le fait qu'elles sont composées d'acides gras liés à des phospholipides par des éthers.
- Certaines molécules peuvent traverser la membrane bactérienne par simple diffusion, mais la plupart des grosses molécules doivent être transportées activement à travers les structures membranaires grâce à l'énergie cellulaire.
- Les parois des cellules procaryotes peuvent être composées de peptidoglycane (bactéries) ou de pseudopeptidoglycane (archées).
- Les cellules bactériennes à Gram positif sont caractérisées par une épaisse couche de peptidoglycane, tandis que les cellules bactériennes à Gram négatif sont caractérisées par une fine couche de peptidoglycane entourée d'une membrane externe.
- Certaines cellules procaryotes produisent des revêtements de glycocalyx, tels que des capsules et des couches visqueuses, qui facilitent la fixation aux surfaces et/ou l'évasion du système immunitaire de l'hôte.
- Certaines cellules procaryotes possèdent des fimbriae ou des pili, des appendices filamenteux qui facilitent la fixation aux surfaces. Les pili sont également utilisés dans le transfert de matériel génétique entre cellules.
- Certaines cellules procaryotes utilisent un ou plusieurs flagelles pour se déplacer dans l'eau. Les bactéries péritricheuses, qui possèdent de nombreux flagelles, utilisent des canaux et des culbutes pour se déplacer délibérément en direction d'un attractif chimique.
Notes
- 1 H 00 Chan, W. F. Marshall. « Propriétés d'écaillage de la taille des cellules et des organites. » Organogenèse 6 no 2 (2010) : 88—96.
- 2 F. Rothfuss, M. Bender, R. Conrad. « Survie et activité des bactéries dans un sédiment lacustre profond et vieilli (lac de Constance). » Écologie microbienne 33 no 1 (1997) :69—77.
- 3 R. Sinclair et coll. « Persistance des agents sélectionnés de catégorie A dans l'environnement. » Microbiologie appliquée et environnementale 74 no. 3 (2008) :555—563.
- 4 T.J. Silhavy, D. Kahne, S. Walker. « L'enveloppe cellulaire bactérienne. » Perspectives de Cold Spring Harbor en biologie 2 n° 5 (2010) :a000414.
- 5 B. Zuber et coll. « Couche granulaire dans l'espace périplasmique de bactéries à Gram positif et structures fines des septa d'Enterococcus gallinarum et de Streptococcus gordonii révélées par cryo-microscopie électronique de coupes vitreuses. » Journal de bactériologie 188 n° 18 (2006) :6652—6660
- 6 L. Gana, S. Chena, G.J. Jensena. « Organisation moléculaire du peptidoglycane à Gram négatif. » Actes de l'Académie nationale des sciences des États-Unis d'Amérique 105 n° 48 (2008) :1895-18957.
- 7 J.A. Garnetta et coll. « Aperçu structurel de la biogenèse et de la formation du biofilm par le pilus commun d'Escherichia coli. » Actes de l'Académie nationale des sciences des États-Unis d'Amérique 109 n° 10 (2012) :3950—3955.
- 8 T. Proft, E. N. Baker. « Les pili dans les bactéries à Gram négatif et à Gram positif : structure, assemblage et rôle dans la maladie. » Sciences de la vie cellulaire et moléculaire 66 (2009) :613.