
3.4: 真核细胞的独特特征
- Page ID
- 200638

学习目标
- 解释真核细胞的区别特征
- 从物理结构、化学结构和功能方面描述原核细胞的内部和外部结构
- 识别和描述真核细胞特有的结构和细胞器
- 比较和对比在原核细胞和真核细胞中发现的相似结构
真核生物包括原生动物、藻类、真菌、植物和动物。 一些真核细胞是独立的单细胞微生物,而另一些则是多细胞生物的一部分。 真核生物的细胞具有几个显著特征。 最重要的是,真核细胞的定义是存在被复杂的核膜包围的核。 此外,真核细胞的特征是细胞质中存在膜结合的细胞器。 线粒体、内质网(ER)、高尔基仪器、溶酶体和过氧化物酶体等细胞器由细胞骨架固定,细胞骨架是支持细胞内成分运输并有助于维持细胞形状的内部网络(图\(\PageIndex{1}\))。 真核细胞的基因组封装在多条棒状染色体中,而不是大多数原核细胞特征的单个圆形染色体。 表\(\PageIndex{1}\)将真核细胞结构的特征与细菌和古细菌的特征进行了比较。

| 细胞结构 | 原核生物 | 真核生物 | |
|---|---|---|---|
| 细菌 | 古细菌 | ||
| 大小 | 大约 0.5—1 μM | 大约 0.5—1 μM | 大约 5—20 μM |
| 表面积与体积比 | 高 | 高 | 低 |
| Nucleus | 不是 | 不是 | 是的 |
| 基因组特征 |
|
|
|
| 细胞分裂 | 二元裂变 | 二元裂变 | 有丝分裂、减数分裂 |
| 膜脂质成分 |
|
|
|
| 细胞壁成分 |
|
|
|
| 运动结构 | 由鞭毛组成的刚性螺旋鞭毛 | 由古鞭毛组成的刚性螺旋鞭毛 | 由微管组成的柔性鞭毛和纤毛 |
| 膜结合细胞器 | 不是 | 不是 | 是的 |
| 内膜系统 | 不是 | 不是 | 是的(ER、Golgi、溶酶体) |
| 核糖体 | 70 | 70 |
|
细胞形态
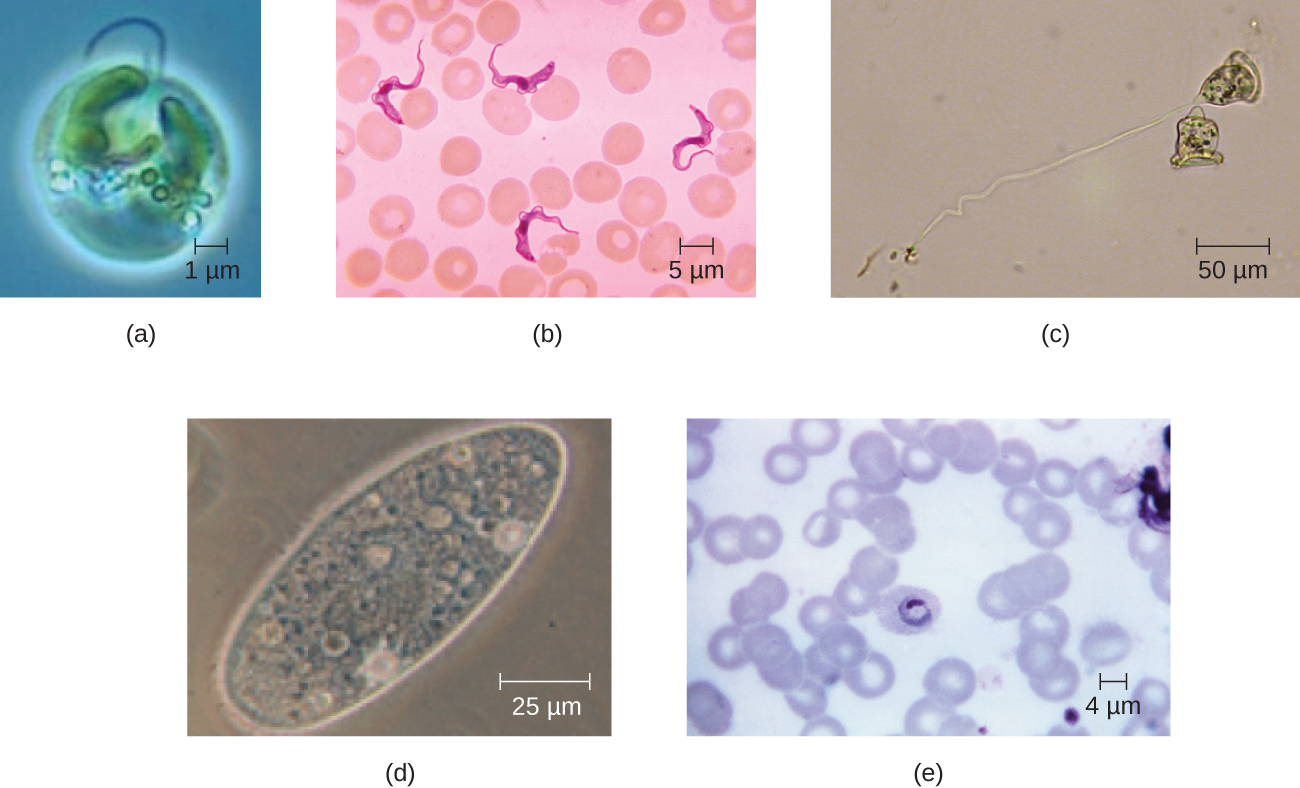
真核细胞表现出各种不同的细胞形态。 可能的形状包括球体、卵形、长方体、圆柱体、扁平体、柱状体、梭形、盘形、新月形、环状星状和多边形(图\(\PageIndex{2}\))。 有些真核细胞的形状不规则,有些能够改变形状。 特定类型的真核细胞的形状可能会受到诸如其主要功能、细胞骨架的组织、细胞质的粘度、细胞膜或细胞壁(如果有的话)的刚性以及周围环境对其施加的物理压力等因素的影响和/或相邻的细胞。
练习\(\PageIndex{1}\)
找出真核细胞和原核细胞之间的两个区别。
Nucleus
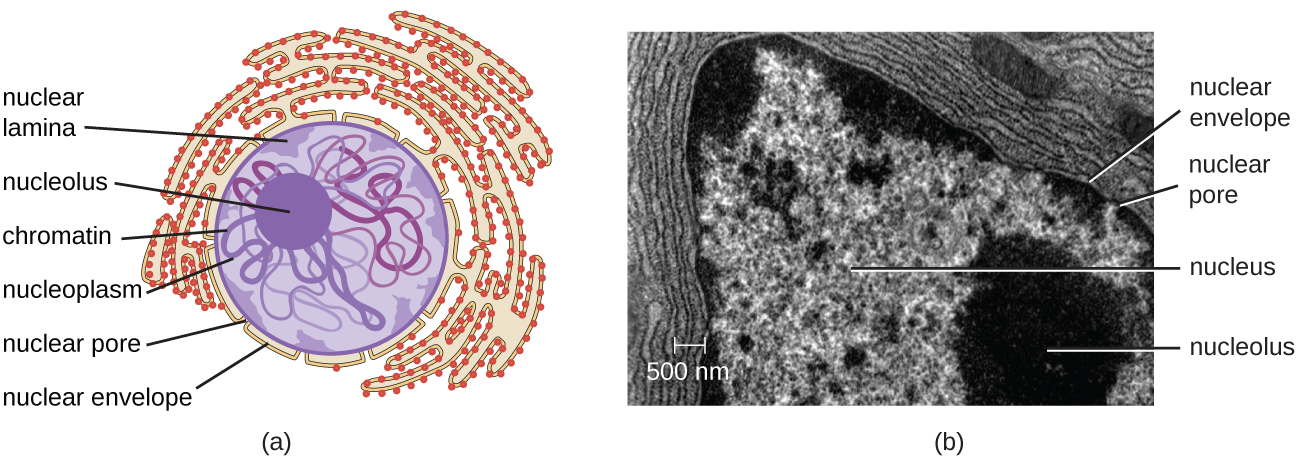
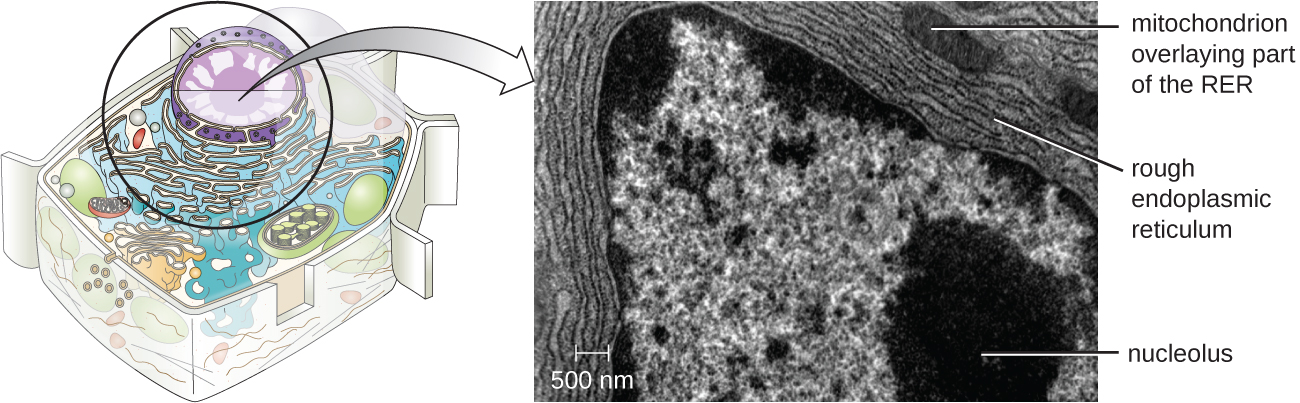
与原核细胞不同,在原核细胞中,DNA松散地包含在核样体区域中,而真核细胞拥有一个核,周围环绕着装有DNA基因组的复杂核膜(图\(\PageIndex{3}\))。 通过包含细胞的DNA,细胞核最终控制了细胞的所有活动,并在繁殖和遗传中起着至关重要的作用。 真核细胞通常将其 DNA 组织成多条线性染色体。 细胞核内的DNA是高度有组织的,经过浓缩以适应细胞核,这是通过将DNA包裹在称为组蛋白的蛋白质周围来实现的。
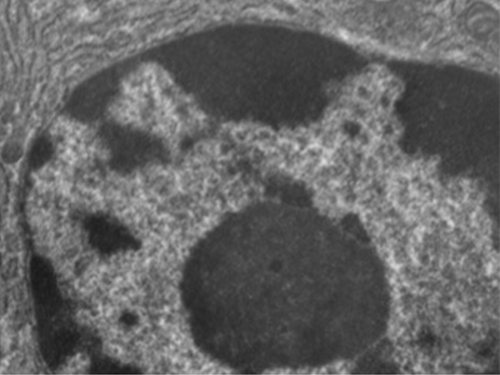
尽管大多数真核细胞只有一个核,但也有例外。 例如,Paramecium 属的原生动物通常有两个完整的核:一个用于繁殖的小核(微核)和一个指导细胞新陈代谢的大核(macronucleus)。 此外,一些真菌在有性生殖过程中会短暂形成具有两个核的细胞,称为异核细胞。 细胞核分裂但细胞质不分裂的细胞被称为共核细胞。
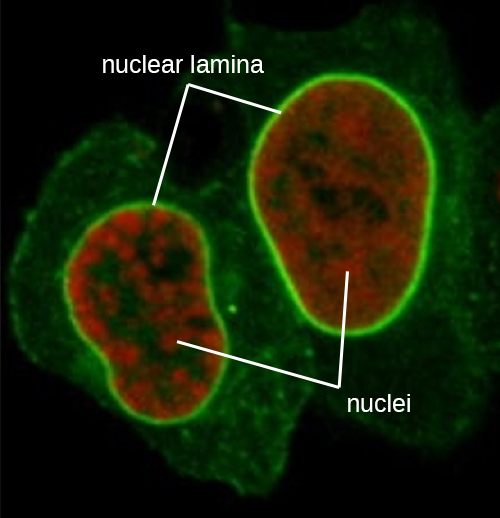
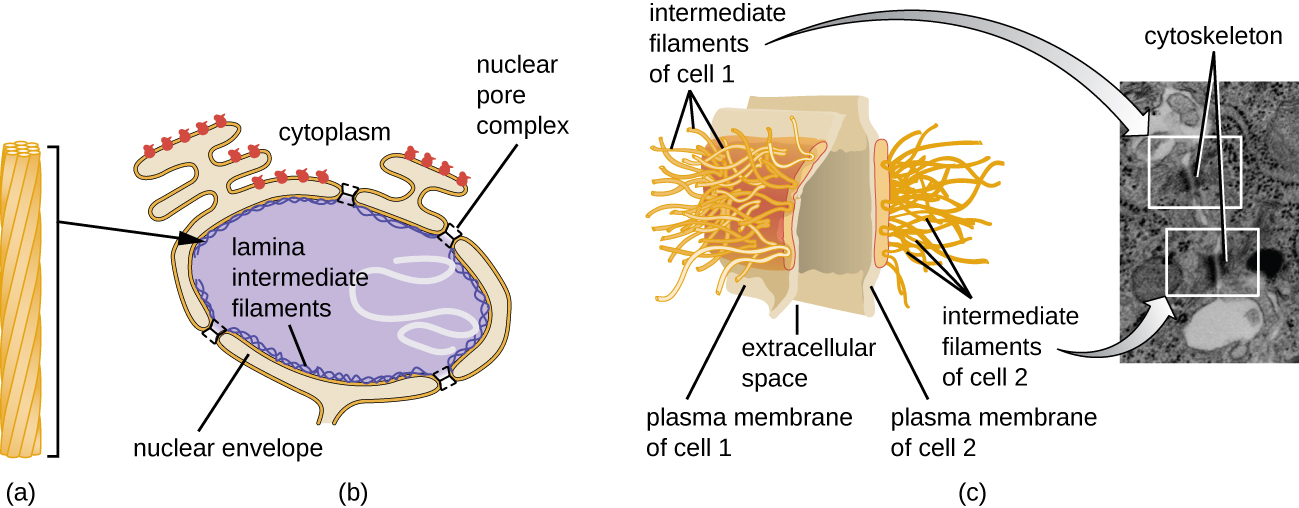
原子核由一个复杂的核膜结合,通常称为核包膜,它由两个彼此相邻的不同脂质双层组成(图\(\PageIndex{4}\))。 尽管内膜和外膜之间存在这些联系,但每个膜的内外表面都含有独特的脂质和蛋白质。 核包膜含有核孔,这些孔是玫瑰花形的大型蛋白质复合物,控制物质进出原子核。 原子核的整体形状由核层决定,核层是由位于核包膜内部的中间细丝组成的网状结构。 在细胞核外,额外的中间细丝形成更松散的网格,用于将细胞核固定在细胞内的位置。
Nucleolus
核仁是核内发生核体RNA(rRNA)生物合成的密集区域。 此外,核仁也是开始组装核糖体的部位。 Preribosomal 复合物由核仁中的 rRNA 和蛋白质组装而成;然后它们被运送到细胞质,在那里完成核糖体的组装(图\(\PageIndex{5}\))。
核糖体
线粒体或叶绿体等真核细胞器中发现的核糖体具有 70S 的核糖体,与原核生物核糖体的大小相同。 但是,真核细胞中的非器官相关核糖体是80年代的核糖体,由40S的小亚基和60年代的大亚基组成。 就大小和成分而言,这使它们与原核细胞的核糖体区分开来。
两种类型的非细胞器相关真核生物核糖体由它们在细胞中的位置定义:游离核糖体和膜结合的核糖体。 游离核糖体存在于细胞质中,用于合成水溶性蛋白质;膜结合的核糖体被发现附着在粗糙的内质网上,产生用于插入细胞膜的蛋白质或注定从细胞出口的蛋白质。
真核生物和原核生物核糖体之间的差异具有临床意义,因为某些抗生素药物是针对其中一种或另一种而设计的。 例如,环己酰亚胺靶向真核作用,而氯霉素靶向原核生物核糖体。 1 由于人体细胞是真核生物,因此它们通常不会受到破坏细菌中原核生物核糖体的抗生素的伤害。 但是,有时可能会出现负面副作用,因为人体细胞中的线粒体含有原核核糖体。
内膜系统
内膜系统是真核细胞独有的,它是一系列膜状小管、囊和扁平盘,它们合成了许多细胞成分并在细胞内移动物质(图\(\PageIndex{6}\))。 由于真核细胞的细胞体积较大,因此需要该系统来输送仅靠扩散无法分散的物质。 内膜系统包括多个细胞器和它们之间的连接,包括内质网、高尔基设备、溶酶体和囊泡。
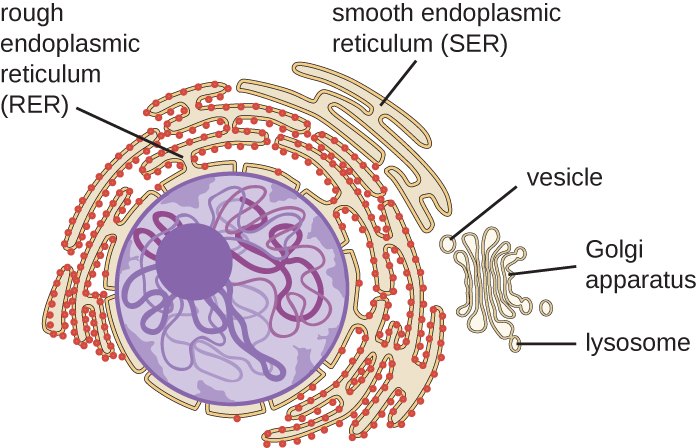
内质网
内质网(ER)是由小管和蓄水池(扁平囊)组成的相互连接的阵列,具有单个脂质双层(图\(\PageIndex{7}\))。 蓄水池内部的空间被称为急诊室的内腔。 急诊室有两种类型:粗糙内质网 (RER) 和平滑内质网 (SER)。 这两种不同类型的急诊室是合成截然不同类型的分子的位点。 RER 上布满了结合在膜细胞质侧的核糖体。 这些核糖体产生运往质膜的蛋白质(图\(\PageIndex{}\))。 合成后,这些蛋白质被插入到RER的膜中。 然后,含有这些新合成蛋白质的 RER 的小囊会作为转运囊泡发芽,然后移至高尔基设备进行进一步处理,直接移到质膜,到另一个细胞器的膜,或者移出细胞。 运输囊泡是单脂、双层、膜状球体,内部空心,携带分子。 SER 没有核糖体,因此看起来 “光滑”。 它参与脂质的生物合成、碳水化合物代谢和细胞内有毒化合物的排毒。
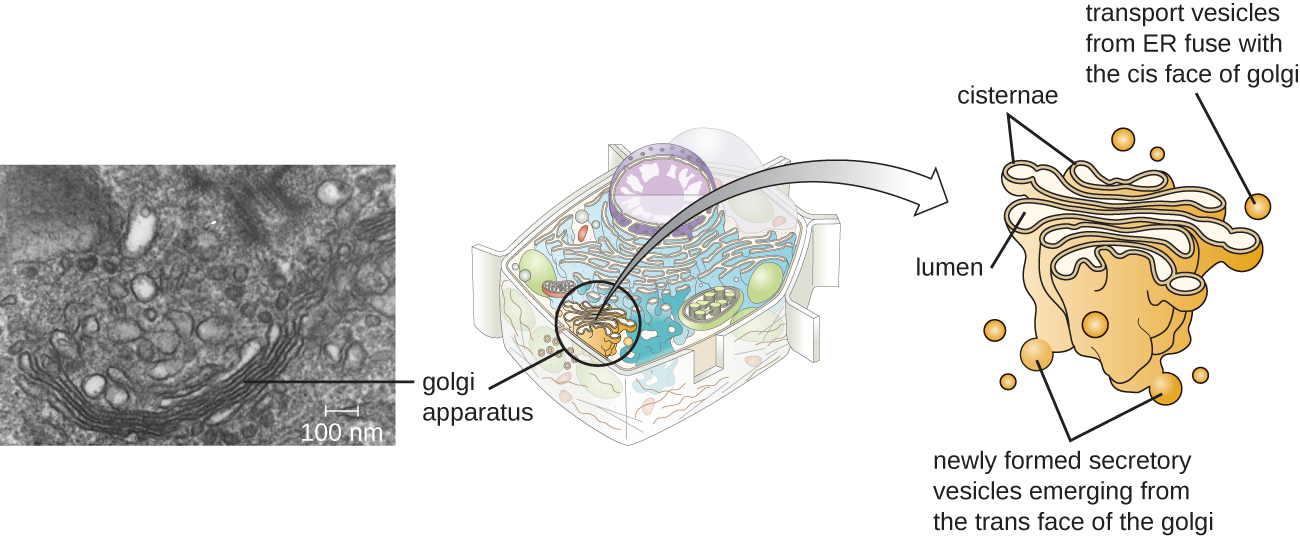
高尔基仪器
1898 年,意大利科学家卡米洛·高尔基(Camillo Golgi)(1843—1926 年)在内膜系统中发现了高尔基仪器,他开发了一种新颖的染色技术,该技术显示了疟原虫细胞内的层叠膜结构。 高尔基仪器由一系列名为 dictyosomes 的膜状圆盘组成,每个盘都有一个单一的脂质双层,它们堆叠在一起(图\(\PageIndex{8}\))。
高尔基仪器中的酶会改变从急诊室运送到高尔基的脂质和蛋白质,经常向它们添加碳水化合物成分,产生糖脂、糖蛋白或蛋白聚糖。 糖脂和糖蛋白通常被插入质膜中,对于其他细胞或传染性颗粒的信号识别很重要。 不同类型的细胞可以通过其血浆膜中所含的糖脂和糖蛋白的结构和排列来区分彼此。 这些糖脂和糖蛋白通常也充当细胞表面受体。
运送囊泡离开急诊室保险丝,其接收面或顺式面上装有高尔基仪器。 这些蛋白质在高尔基仪器内进行处理,然后含有改性蛋白质和脂质的额外转运囊泡在高尔基仪器外出或反面时从高尔基仪器中挤出。 这些流出的囊泡移动到质膜或其他细胞器的膜并与之融合。
Exocytosis 是分泌囊泡(球状膜囊)将其内容物释放到细胞外部的过程(图\(\PageIndex{8}\))。 所有细胞都有组成分泌途径,分泌囊泡通过这些途径转运可持续(组成地)从细胞中释放的可溶性蛋白。 某些特殊细胞也有调节的分泌途径,用于在分泌囊泡中储存可溶性蛋白。 受管制的分泌物涉及仅在响应某些事件或信号时才释放的物质。 例如,人体免疫系统的某些细胞(例如肥大细胞)会因体内存在异物或病原体而分泌组胺。 组胺是一种触发免疫系统消灭病原体的各种机制的化合物。
溶酶体
1960年代,比利时科学家克里斯蒂安·德·杜夫(Christian de Duve)(1917-2013)发现了溶酶体,即内膜系统中含有消化酶的膜结合细胞器。 某些类型的真核细胞使用溶酶体来分解各种颗粒,例如食物、受损的细胞器或细胞碎片、微生物或免疫复合物。 溶酶体内消化酶的分隔使细胞能够在不损害细胞的细胞质成分的情况下有效地消化物质。
练习\(\PageIndex{2}\)
命名内膜系统的组成部分并描述每个成分的功能。
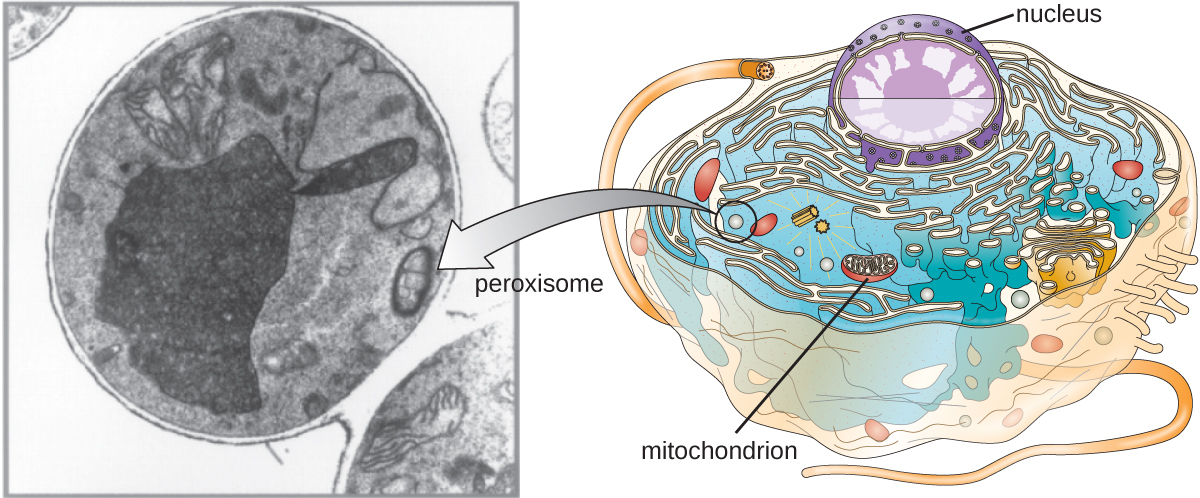
过氧化物酶体
克里斯蒂安·德·杜夫(Christian de Duve)还因发现过氧化物酶体而受到赞誉,过氧化物酶体是膜结合的细胞器,不属于内膜系统(图\(\PageIndex{9}\))。 过氧化物酶体通过游离核糖体合成过氧化物蛋白以及将这些过氧化物蛋白掺入现有过氧化物酶体中而在细胞质中独立形成。 然后,生长的过氧化物酶体除以类似于二元裂变的过程。
过氧化物酶体最初因其产生过氧化氢的能力而得名,过氧化氢是一种高活性分子,有助于分解尿酸、氨基酸和脂肪酸等分子。 过氧化物酶体还含有过氧化氢酶,它可以降解过氧化氢。 除了 SER 外,过氧化物酶体还在脂质生物合成中发挥作用。 与溶酶体一样,这些降解分子在细胞器内的分隔有助于保护细胞质内容物免受不必要的损害。
某些生物的过氧化物酶体专门用于满足其特定的功能需求。 例如,glyoxysomes 是酵母和植物细胞的改性过氧化物酶体,具有多种代谢功能,包括糖分子的产生。 同样,糖体是由某些锥虫产生的改性过氧化物酶体,锥虫是导致恰加斯病和非洲昏睡病的致病原生动物。
细胞骨架
真核细胞具有由微丝、中间细丝和微管组成的内部细胞骨架。 这种纤维和管道的矩阵提供了结构支撑以及一个网络,材料可以在细胞内运输,细胞器可以固定在网络上(图\(\PageIndex{10}\))。 例如,exocytosis 过程涉及囊泡通过细胞骨架网络移动到质膜,在那里它可以释放其内容物。
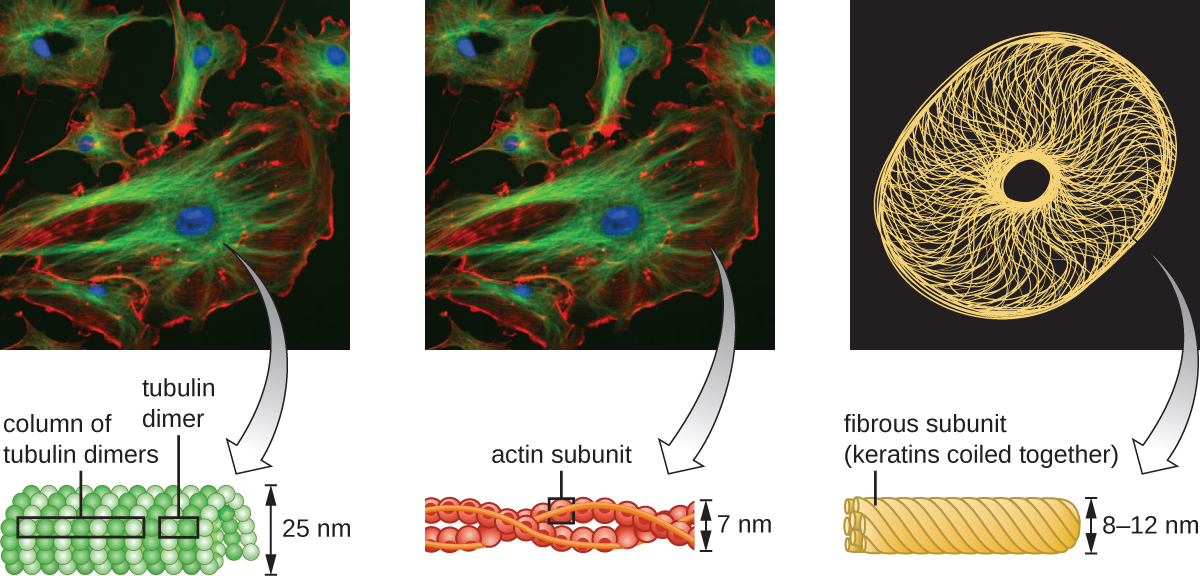
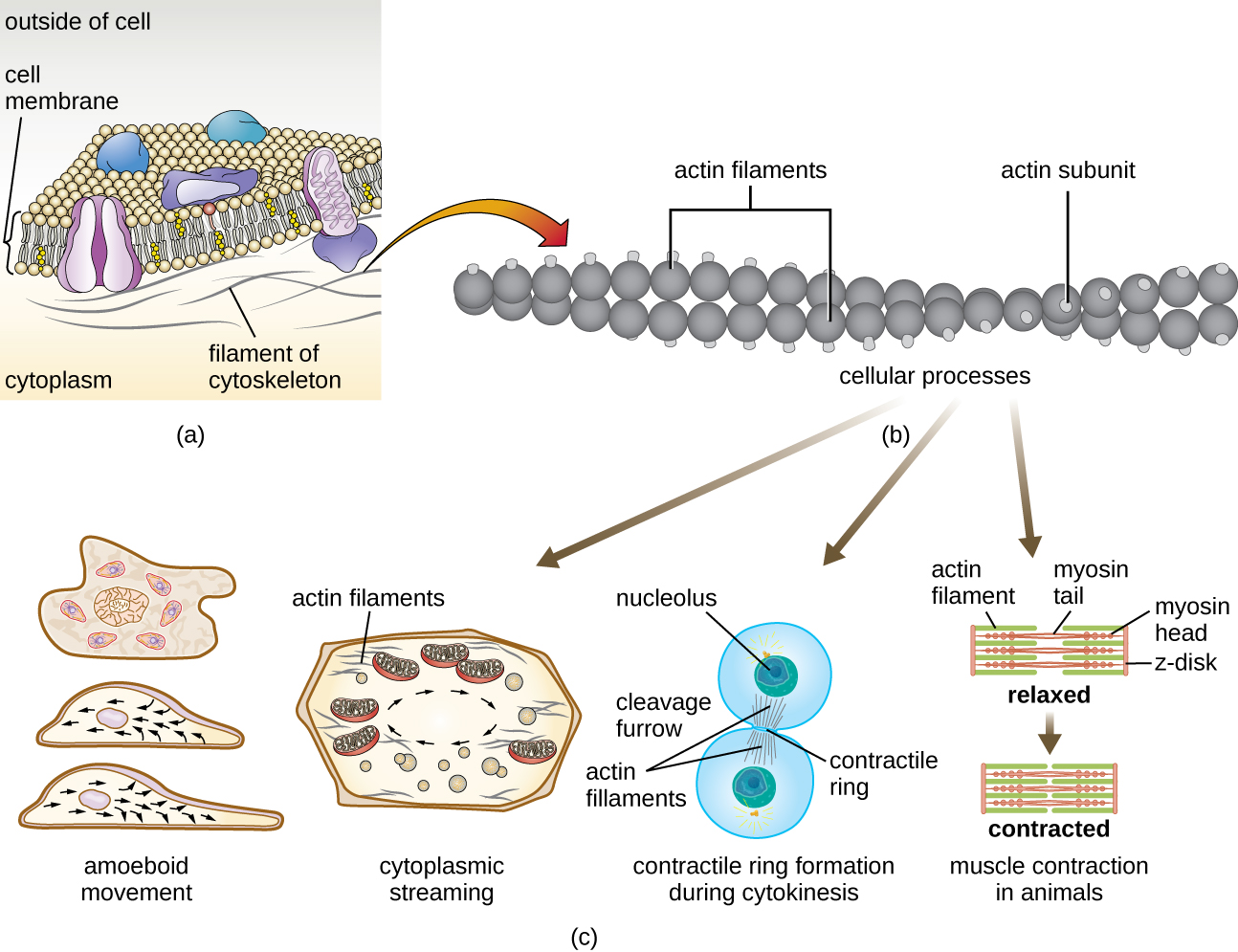
微丝由两条交织在一起的肌动蛋白链组成,每股由肌动蛋白单体组成,形成直径为6 nm的丝状电缆(图\(\PageIndex{11}\))。 肌动蛋白丝与肌球蛋白等运动蛋白一起作用,影响动物的肌肉收缩或某些真核微生物的变形体运动。 在 ameboid 生物体中,肌动蛋白有两种形式:更硬的聚合凝胶形式和更流体、未聚合的可溶性形式。 凝胶形式的肌动蛋白在外质中产生稳定性,外质是位于阿米巴体原生动物质膜内的凝胶状细胞质区域。
被称为 pseudopodia(意思是 “假脚”)的细胞质膜的临时延伸是通过可溶性肌动蛋白丝向前流入伪足体产生的,然后是肌动蛋白丝的凝胶溶胶循环,从而产生细胞活力。 一旦细胞质向外延伸,形成伪足体,剩余的细胞质就会向上流动,加入前沿,从而产生向前运动。 除了变形虫运动外,微丝还参与真核细胞中的各种其他过程,包括细胞质流(细胞内细胞质的运动或循环)、细胞分裂期间的裂沟形成以及动物的肌肉运动(图\(\PageIndex{11}\))。 这些功能是微丝的动态特性的结果,微丝可以相对容易地聚合和解聚,以响应细胞信号,以及它们与不同类型的真核细胞中的分子马达相互作用。
中间细丝(图\(\PageIndex{12}\))是一组多样化的细胞骨架细丝,充当细胞内的电缆。 它们之所以被称为 “中间体”,是因为它们的直径为10 nm,比肌动蛋白的直径更厚,但比微管的直径更薄。 3 它们由几股聚合亚基组成,而这些亚基又由各种各样的单体组成。 中间的细丝往往在细胞中更持久,并维持细胞核的位置。 它们还在核外壳内形成核层(衬里或层)。 此外,中间细丝在将动物组织中的细胞固定在一起方面起着作用。 中间长丝蛋白 desmin 存在于 desmosomes 中,desmosomes 是将肌肉细胞连接在一起并帮助它们抵抗外部物理力量的蛋白质结构。 中间长丝蛋白角蛋白是一种存在于头发、皮肤和指甲中的结构蛋白。
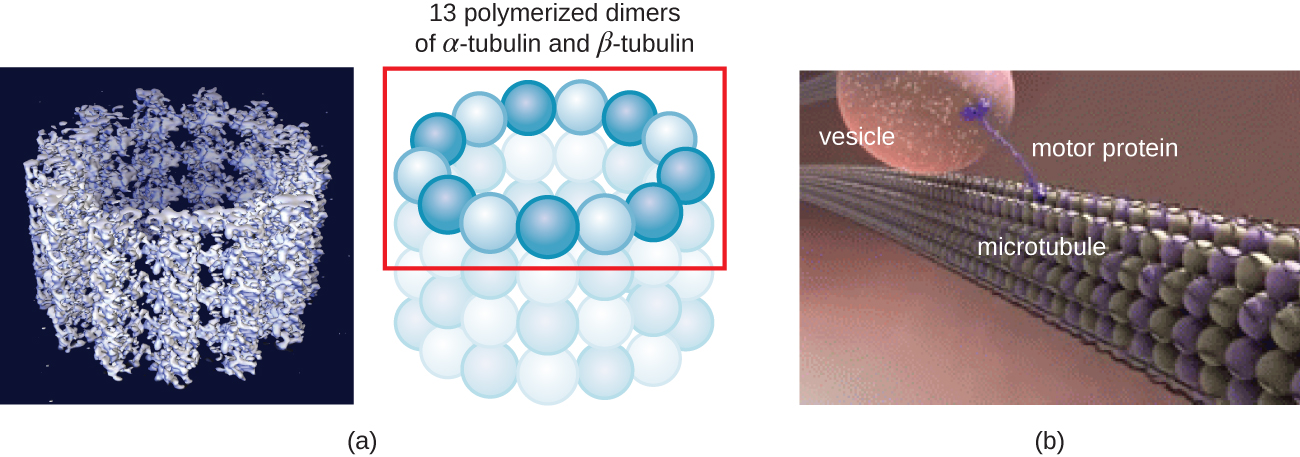
微管(图\(\PageIndex{13}\))是第三种由微管蛋白二聚体(α微管蛋白和β微管蛋白)组成的细胞骨架纤维。 它们形成直径为 23 nm 的空心管,用作细胞骨架内的大梁。 4 像微丝一样,微管是动态的,能够快速组装和拆卸。 微管还与运动蛋白(例如 dynein 和 kinesin)一起作用,在细胞质内移动细胞器和囊泡。 此外,微管是真核生物鞭毛和纤毛的主要成分,构成细丝和基体成分(图\(\PageIndex{20}\))。
此外,微管参与细胞分裂,形成有丝分裂纺锤,用于在有丝分裂和减数分裂期间分离染色体。 有丝分裂纺锤由位于细胞两端的两个中心体产生,它们本质上是微管组织中心。 每个中心体由一对彼此成直角的中心体组成,每个中心体是由九个以三重形式排列的平行微管组成的阵列(图\(\PageIndex{14}\))。
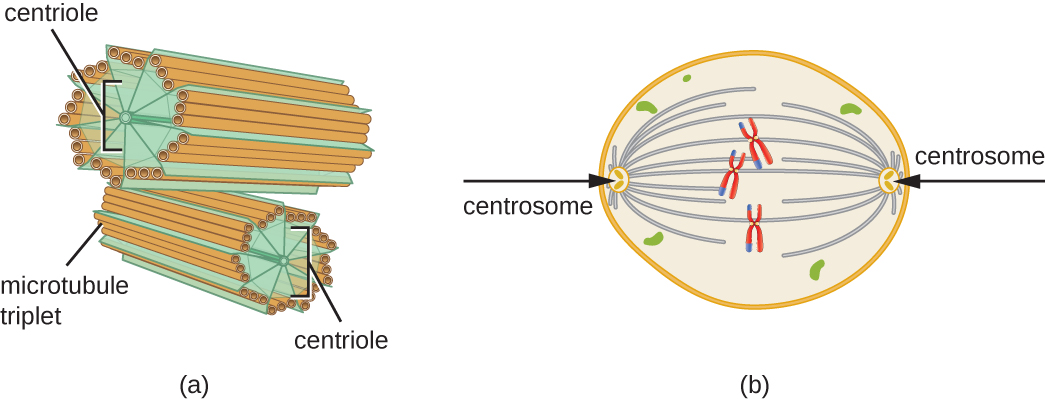
练习\(\PageIndex{3}\)
比较和对比本节中描述的三种类型的细胞骨架结构。
线粒体
真核细胞中发生有氧细胞呼吸的大型复杂细胞器被称为线粒体(图\(\PageIndex{15}\))。 “线粒体” 一词最初是由德国微生物学家卡尔·本达在1898年创造的,后来与奥托·沃尔堡在1913年的呼吸过程联系起来。 1960年代,科学家发现线粒体有自己的基因组和70年代的核糖体。 线粒体基因组在 1976 年进行测序时被发现是细菌。 这些发现最终支持了林恩·马古利斯提出的内共生理论,该理论指出,线粒体最初是通过内共生事件产生的,在这种事件中,一种能够进行有氧细胞呼吸的细菌被吞噬吸收到宿主细胞中,并作为可存活的细胞内细胞继续存在组件。
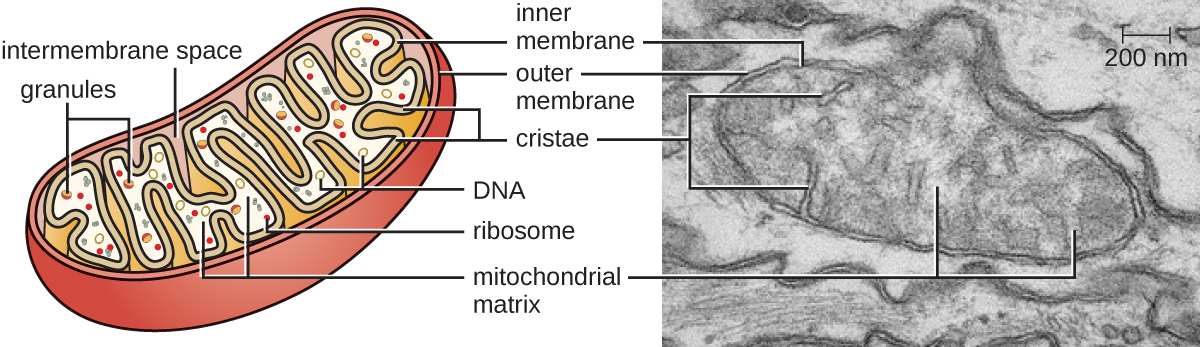
每个线粒体都有两个脂质膜。 外膜是原始宿主细胞膜结构的残余物。 内膜来自细菌质膜。 用于有氧呼吸的电子传输链使用嵌入内膜的整体蛋白质。 线粒体基质对应于原始细菌细胞质的位置,是许多代谢酶的当前位置。 它还包含线粒体DNA和70S核糖体。 被称为 cristae 的内膜的侵入演变为增加生化反应位置的表面积。 cristae 的折叠模式因不同类型的真核细胞而异,用于区分不同的真核生物。
叶绿体
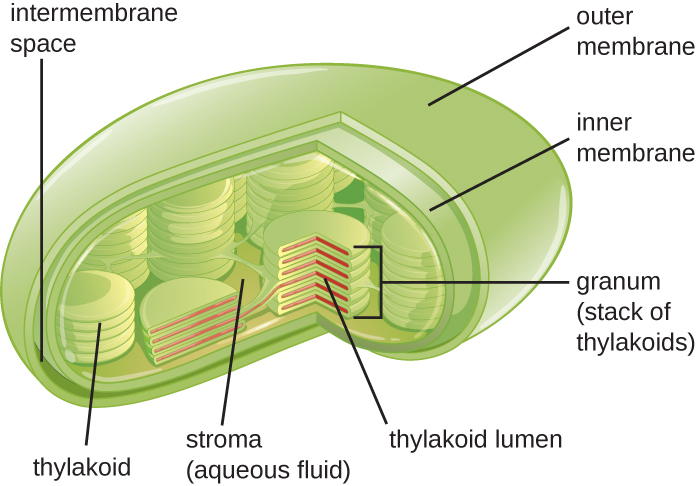
植物细胞和藻类细胞含有叶绿体,叶绿体是发生光合作用的细胞器(图\(\PageIndex{16}\))。 所有叶绿体都有至少三个膜系统:外膜、内膜和类囊体膜系统。 外膜和内膜内部是叶绿体基质,这是一种凝胶状液体,构成叶绿体体积的大部分,tylakoid 系统漂浮在其中漂浮。 thylakoid 系统是一个高度动态的折叠膜囊集合。 这是发现绿色光合色素叶绿素的地方,也是光合作用的光反应发生的地方。 在大多数植物叶绿体中,thylakoids 以名为 grana(单数:granum)的堆叠排列,而在某些藻类叶绿体中,thylakoids 是自由漂浮的。
其他与线粒体相似的细胞器也出现在其他类型的真核生物中,但它们的作用不同。 氢体存在于一些厌氧真核生物中,是厌氧氢产生的地点。 氢体通常缺乏自身的 DNA 和核糖体。 Kinetoplasts 是某些真核病原体中发现的线粒体的一种变体。 在这些生物体中,每个细胞都有一个单一的、长的、分支的线粒体,其中发现以多个圆形 DNA 片段组织的 kinetoplast DNA 集中在细胞的一个极点。
原生动物寄生虫中的线粒体相关细胞器
许多原生动物,包括几种引起人类感染的原生动物寄生虫,可以通过其不寻常的外观来识别。 区别特征可能包括复杂的细胞形态、独特的细胞器的存在或没有常见的细胞器。 原生动物寄生虫 Giardia lamblia 和 Trichomonas vaginalis 就是两个例子。
G. lambli a 是人类和许多其他动物腹泻的常见原因,是一种厌氧寄生虫,具有两个细胞核和几个鞭毛。 它的高尔基仪器和内质网已大大减少,并且完全缺乏线粒体。 但是,它确实有被称为有丝体的细胞器,即双膜结合的细胞器,线粒体似乎严重减少。 这使科学家们相信 G. lambli a 的祖先曾经拥有进化为有丝体的线粒体。 导致性传播感染@@ 阴道炎的阴道杆菌是另一种缺乏传统线粒体的原生动物寄生虫。 相反,它拥有氢体、线粒体相关的、双膜结合的细胞器,可产生用于细胞代谢的分子氢。 科学家们认为,像有丝分体这样的氢体也是从线粒体进化而来的。 5
质膜
真核细胞的质膜在结构上与原核质膜相似,因为它主要由磷脂组成,形成带有嵌入外周和整体蛋白的双层(图\(\PageIndex{17}\))。 根据流体镶嵌模型,这些膜组件在膜的平面内移动。 但是,与原核细胞膜不同,真核细胞膜含有固醇,包括胆固醇,会改变膜的流动性。 此外,许多真核细胞含有一些特殊的脂质,包括鞘脂,这些脂质被认为在维持膜稳定性以及参与信号转导途径和细胞间通信方面起着作用。

膜传输机制
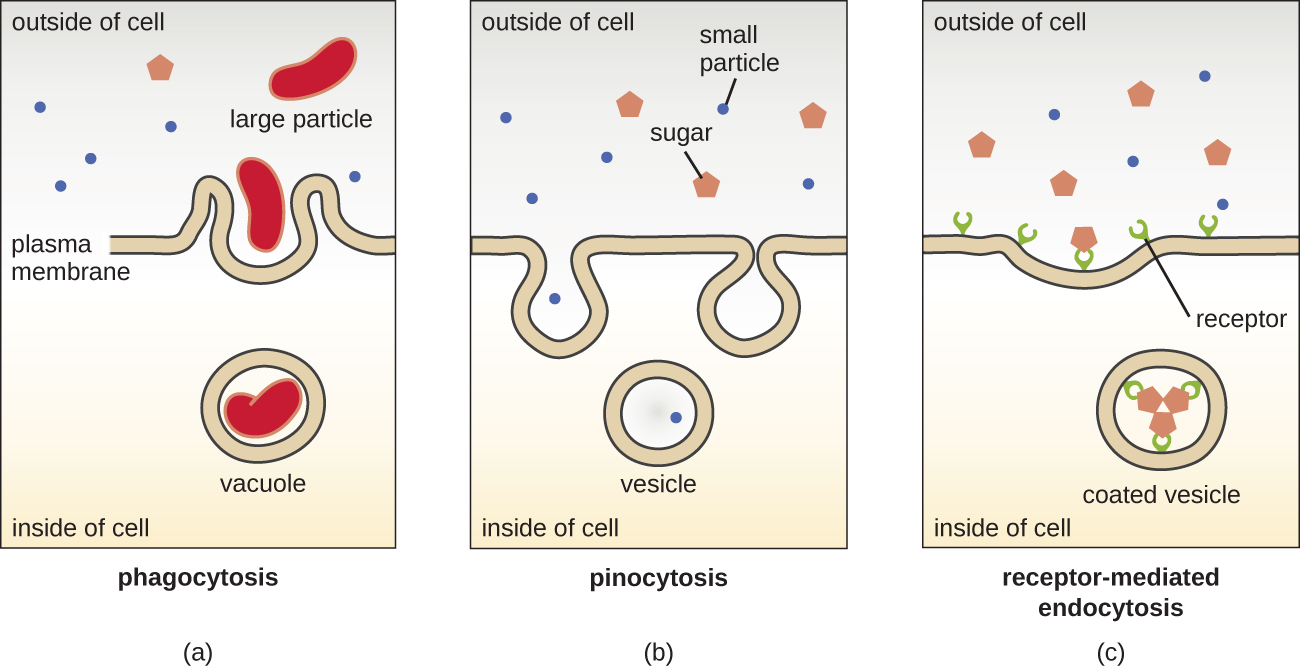
在真核细胞和原核细胞中都使用简单扩散、促进扩散和活性转运的过程。 但是,真核细胞还具有进行各种类型的内吞作用、通过质膜侵入和液泡/囊泡形成吸收物质的独特能力(图\(\PageIndex{18}\))。 一种涉及通过膜侵入吞没大颗粒的内吞作用称为吞噬作用,意思是 “吃细胞”。 在吞噬作用中,颗粒(或其他细胞)被封闭在膜内的口袋中,然后膜从膜上挤出,形成完全包围颗粒的液泡。 另一种类型的内吞作用叫做 pinocytosis,意思是 “细胞饮用”。 在 pinocytosis 中,溶解的小物质和液体通过小囊泡被吸入细胞。 例如,腐生真菌主要通过 pinocytosis 从死亡和腐烂的物质中获取营养。
受体介导的内吞作用是一种内吞作用,由称为配体的特定分子在与膜上的细胞表面受体结合时引发。 受体介导的内吞作用是肽和胺衍生激素进入细胞的机制,也被各种病毒和细菌用来进入宿主细胞。
分泌囊泡将其内容物释放到细胞外部的过程称为外吞作用。 囊泡向质膜移动,然后与质膜融合,将其内容物排出细胞。 细胞使用 Exocytosis 去除废物,也可用于释放可被其他细胞吸收的化学信号。
细胞墙
除了质膜外,一些真核细胞还有细胞壁。 真菌、藻类、植物乃至某些原生生物的细胞都有细胞壁。 根据真核细胞的类型,细胞壁可以由多种材料制成,包括纤维素(真菌和植物);生物二氧化硅、碳酸钙、琼脂和角叉菜胶(原生生物和藻类);或甲壳素(真菌)。 总的来说,所有细胞壁都为细胞提供结构稳定性,并保护细胞免受干燥、渗透压变化和创伤等环境压力的影响。 6
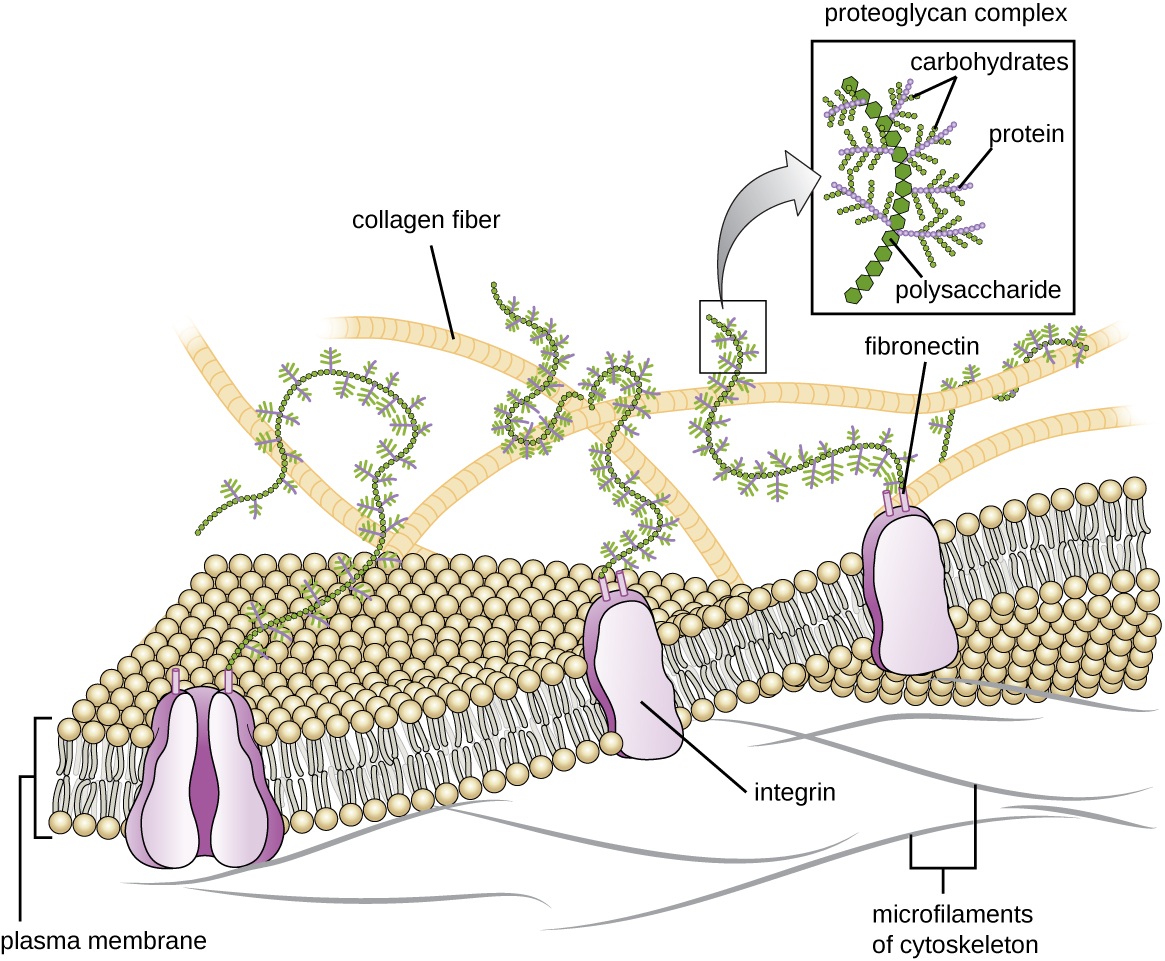
细胞外基质
动物和一些原生动物的细胞没有细胞壁来帮助维持形状和提供结构稳定性。 相反,这些类型的真核细胞会为此目的产生细胞外基质。 它们将粘稠的碳水化合物和蛋白质分泌到相邻细胞之间的空间中(图\(\PageIndex{19}\))。 一些蛋白质成分聚成基底膜,剩余的细胞外基质成分粘附在基底膜上。 蛋白聚糖通常形成细胞外基质的体积块,而纤维蛋白,如胶原蛋白,则提供强度。 蛋白聚糖和胶原蛋白都附着在纤维连接蛋白上,而纤维连接蛋白反过来又附着在整合素蛋白上。 这些整合素蛋白与缺乏细胞壁的真核细胞质膜中的跨膜蛋白相互作用。
在动物细胞中,细胞外基质使组织内的细胞能够承受外部压力,并将信号从细胞外部传输到内部。 在各种类型的结缔组织中,细胞外基质的量相当广泛,细胞外基质的变化可以赋予不同类型的组织不同的特性。 此外,宿主细胞的细胞外基质通常是微生物病原体附着以建立感染的部位。 例如,化脓性链球菌是一种引起链球菌性喉炎和其他各种感染的细菌,它与口咽内细胞(喉咙上部区域)的细胞外基质中的纤维连接蛋白结合。
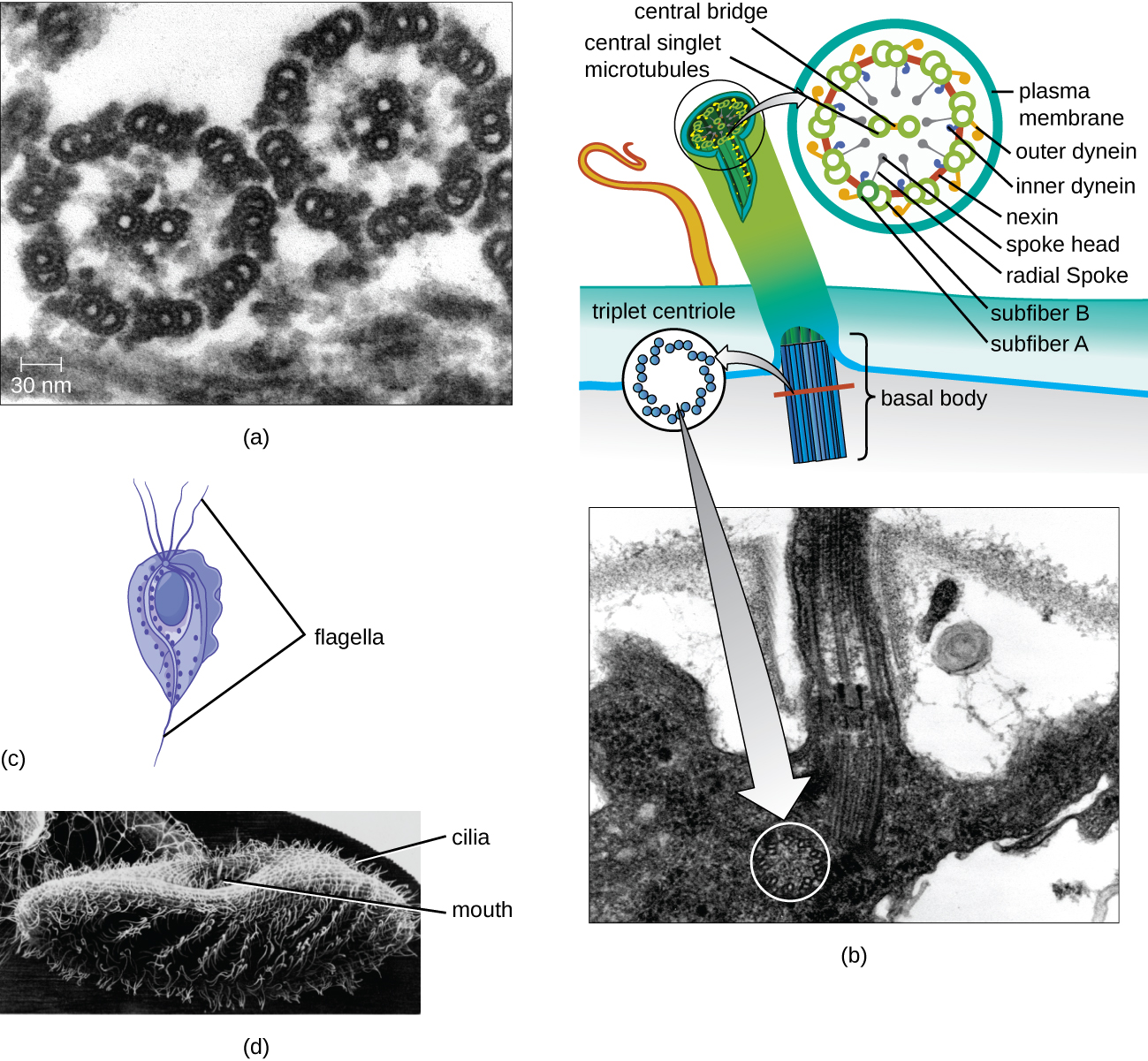
鞭毛和纤毛
一些真核细胞使用鞭毛进行运动;但是,真核鞭毛在结构上与原核细胞中发现的鞭毛不同。 原核鞭毛是一种坚硬的旋转结构,而真核生物鞭毛更像是一根柔性鞭子,由围绕一对中心微管的平行微管组成。 这种排列方式被称为 9+2 数组(图\(\PageIndex{20}\))。 平行微管使用 dynein 运动蛋白相互移动,导致鞭毛弯曲。
纤毛(单数:纤毛)是在一些真核细胞中发现的一种类似的外部结构。 纤毛是真核生物独有的,它比鞭毛短,通常覆盖细胞的整个表面;但是,它们在结构上与鞭毛(一个 9+2 的微管阵列)相似,并且使用相同的运动机制。 在每根纤毛和鞭毛的底部都发现了一种叫做基体的结构。 将纤毛或鞭毛附着在细胞上的基体由一系列类似于中心体的三重微管组成,但嵌入在质膜中。 由于纤毛的长度较短,因此使用快速、灵活、挥舞的动作。 除运动外,纤毛还可能具有其他功能,例如将颗粒扫过或进入细胞。 例如,纤毛的原生动物利用清扫纤毛将食物颗粒移入口腔,哺乳动物呼吸道中的纤毛细胞同步跳动,将粘液和碎屑从肺部上下扫出(图\(\PageIndex{20}\))。
练习\(\PageIndex{4}\)
- 解释真核细胞的细胞包膜与原核细胞的细胞包膜相比如何。
- 解释真核生物和原核鞭毛之间的区别。
临床重点:分辨率
由于阿莫西林尚未解决芭芭拉的肺炎病例,巴勒斯坦权力机构开了另一种抗生素阿奇霉素,该抗生素靶向细菌核糖体而不是肽聚糖。 按照指示服用阿奇霉素后,芭芭拉的症状消失了,她终于开始恢复了自己的感觉。 假设不涉及对阿莫西林的耐药性,并考虑到阿奇霉素的有效性,芭芭拉肺炎的病原体很可能是肺炎支原体。 尽管这种细菌是原核细胞,但它不会受到阿莫西林的抑制,因为它没有细胞壁,因此不会产生肽聚糖。
关键概念和摘要
- 真核细胞的定义是存在一个包含 DNA 基因组的核,并由一个由两个脂质双层组成的核膜(或核包膜)结合,这两个脂质双层调节物质通过核孔进出细胞的运输。
- 真核细胞的形态差异很大,可能由各种结构维持,包括细胞骨架、细胞膜和/或细胞壁。
- 位于真核细胞核中的核仁是核糖体合成的部位,也是核糖体组装的第一阶段。
- 真核细胞在粗糙的内质网(膜结合的核糖体)和细胞质(游离核糖体)中含有 80 S 的核糖体。 它们在线粒体和叶绿体中含有 70 年代的核糖体。
- 真核细胞已经进化出一种内膜系统,其中含有参与运输的膜结合细胞器。 这些包括囊泡、内质网和高尔基设备。
- 光滑的内质网在脂质生物合成、碳水化合物代谢和有毒化合物的排毒中起着作用。 粗糙的内质网含有膜结合的 80S 核糖体,它们合成了运往细胞膜的蛋白质
- 高尔基仪器通常通过添加糖分子来处理蛋白质和脂质,产生糖蛋白或糖脂,糖脂是用于细胞间通信的质膜成分。
- 溶酶体含有消化酶,可分解内吞作用摄入的小颗粒、吞噬作用摄入的大颗粒或细胞,以及受损的细胞内成分。
- 细胞骨架由微丝、中间细丝和微管组成,为真核细胞提供结构支持,并充当细胞内物质的运输网络。
- 中心体是微管组织中心,在有丝分裂中形成有丝分裂纺锤中很重要。
- 线粒体是细胞呼吸的部位。 它们有两个膜:外膜和带有 cristae 的内膜。 内膜内的线粒体基质包含线粒体DNA、70S核糖体和代谢酶。
- 真核细胞的质膜在结构上与原核细胞中的质膜相似,膜成分根据流体镶嵌模型移动。 但是,真核细胞膜含有改变膜流动性的固醇,以及糖蛋白和糖脂,后者有助于细胞识别其他细胞和传染性颗粒。
- 除了主动转运和被动转运外,真核细胞膜还可以通过内吞作用将物质带入细胞,或者通过外吞作用将物质从细胞中排出。
- 真菌、藻类、植物和一些原生生物的细胞有细胞壁,而动物和一些原生动物的细胞具有粘性细胞外基质,可提供结构支持并介导细胞信号传导。
- 真核生物鞭毛在结构上不同于原核生物鞭毛,但其用途(运动)类似。 纤毛在结构上与真核生物鞭毛相似,但较短;它们可用于细胞外颗粒的运动、喂食或运动。
脚注
- 1 A.E. Barnhill、M.T. Brewer、S.A. Carlson。 “抗微生物药物通过可预测或特异性抑制宿主线粒体成分产生的不良影响。” 抗微生物药物和化疗 56 no.8 (2012): 4046—4051。
- 2 Fuchs E,克利夫兰 DW。 “健康与疾病中中间细丝的结构脚手架。” S@@ c ience 279 第 5350 号 (1998): 514—519。
- 3 E. Fuchs,D.W. Cleveland。 “健康与疾病中中间细丝的结构脚手架。” S@@ c ience 279 第 5350 号 (1998): 514—519。
- 4 E. Fuchs,D.W. Cleveland。 “健康与疾病中中间细丝的结构脚手架。” S@@ c ience 279 第 5350 号 (1998): 514—519。
- 5 N. Yarlett,J.H.P. Hackstein。 “氢体:一个细胞器,多个来源。” BioScienc e 55 第 8 期 (2005): 657—658。
- 6 M. Dudzick。 “原生动物。” OpenStax CNX。 2013 年 11 月 27 日。 http://cnx.org/contents/f7048bb6-e46...ef291cf7049c@1