
3.3: 原核细胞的独特特征
- Page ID
- 200655

学习目标
- 解释原核细胞的区别特征
- 描述原核细胞的常见细胞形态和典型的细胞排列,并解释细胞如何保持其形态
- 从物理结构、化学结构和功能方面描述原核细胞的内部和外部结构
- 比较细菌和古细胞的区别特征
细胞理论指出,细胞是生命的基本单位。 但是,细胞的大小、形状、结构和功能差异很大。 在最简单的结构层面上,所有细胞都具有一些基本成分。 其中包括细胞质(一种由水和生长所需的溶解化学物质组成的凝胶状物质),它包含在质膜(也称为细胞膜或细胞质膜)中;一条或多条含有细胞遗传蓝图的染色体;以及核糖体,即使用的细胞器用于生产蛋白质。
除了这些基本成分外,生物体之间的细胞差异很大,甚至在同一个多细胞生物体内也是如此。 最大的两类细胞——原核细胞和真核细胞——是由几种细胞结构的主要差异定义的。 原核细胞缺乏被复杂核膜包围的核,通常在核样体中有一条圆形的染色体。 真核细胞的细胞核被复杂的核膜包围,该核膜包含多条棒状染色体。 1
所有植物细胞和动物细胞都是真核细胞。 有些微生物由原核细胞组成,而另一些则由真核细胞组成。 原核微生物被归类为古细菌和细菌域,而真核生物被归类为真核生物域。
细胞内部的结构类似于人体内部的器官,具有适合特定功能的独特结构。 在原核细胞中发现的某些结构与某些真核细胞中发现的结构相似;其他结构是原核生物所独有的。 尽管有一些例外,但真核细胞往往比原核细胞大。 真核细胞的体积相对较大,这决定了使用复杂的膜结合细胞器在细胞的不同区域内划分各种化学过程的必要性。 相比之下,原核细胞通常缺乏膜结合的细胞器;但是,它们通常含有分隔其细胞质的内含物。 该图\(\PageIndex{1}\)说明了通常与原核细胞相关的结构。 下一节将更详细地介绍这些结构。
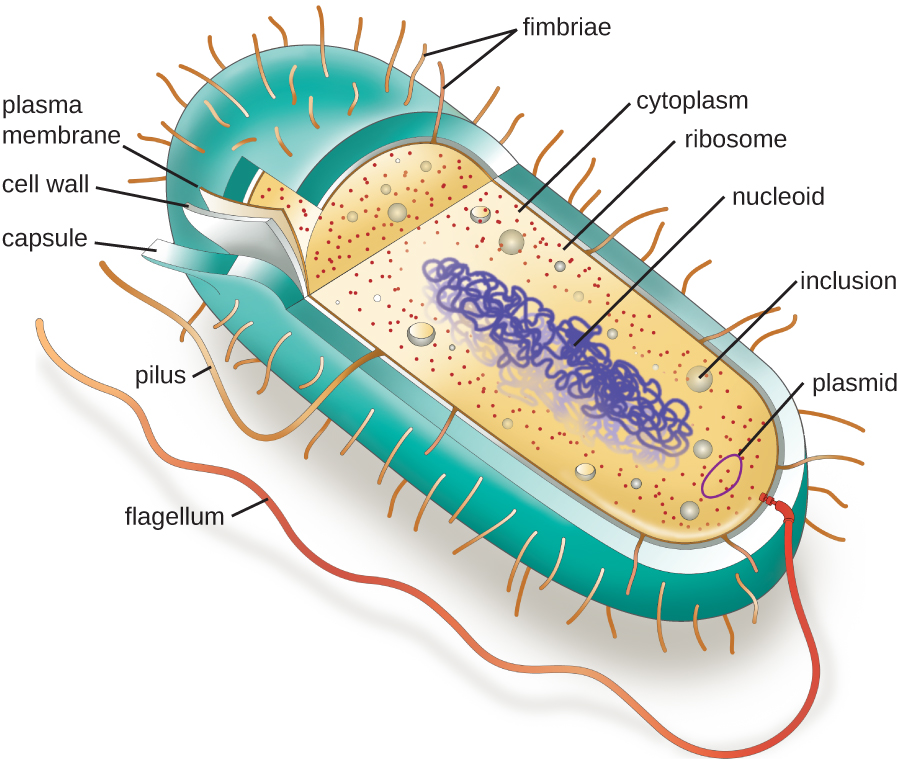
常见的细胞形态和排列
特定原核生物的单个细胞在形状或细胞形态上通常相似。 尽管已经鉴定出成千上万的原核生物,但在显微镜下通常只能看到少数几种细胞形态。 用图形\(\PageIndex{2}\)命名并说明原核细胞中常见的细胞形态。 除了细胞形状外,相同物种的原核细胞可能根据细胞分裂平面以某些不同的排列组合在一起。 一些常见的安排如图所示\(\PageIndex{3}\)。
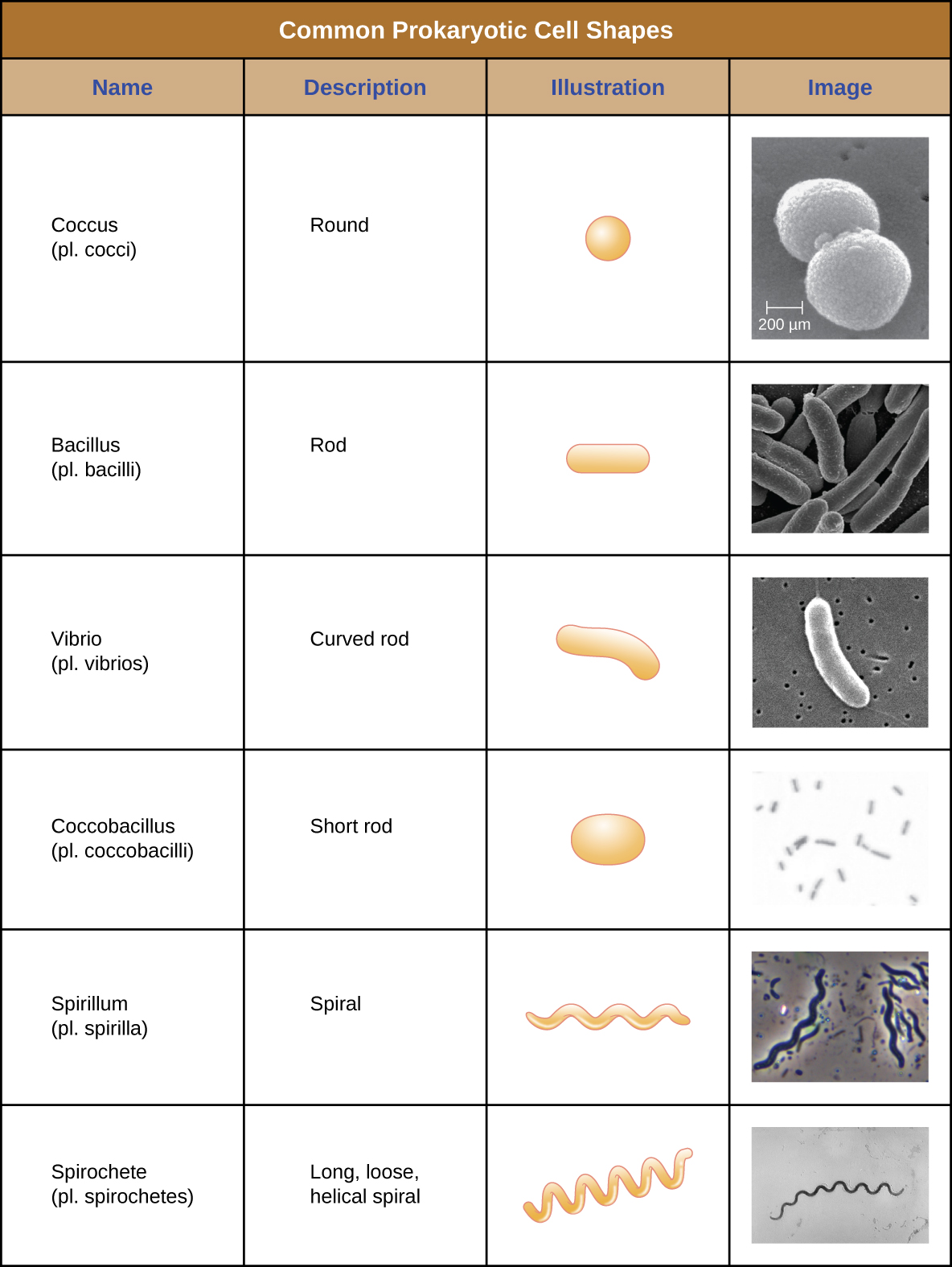
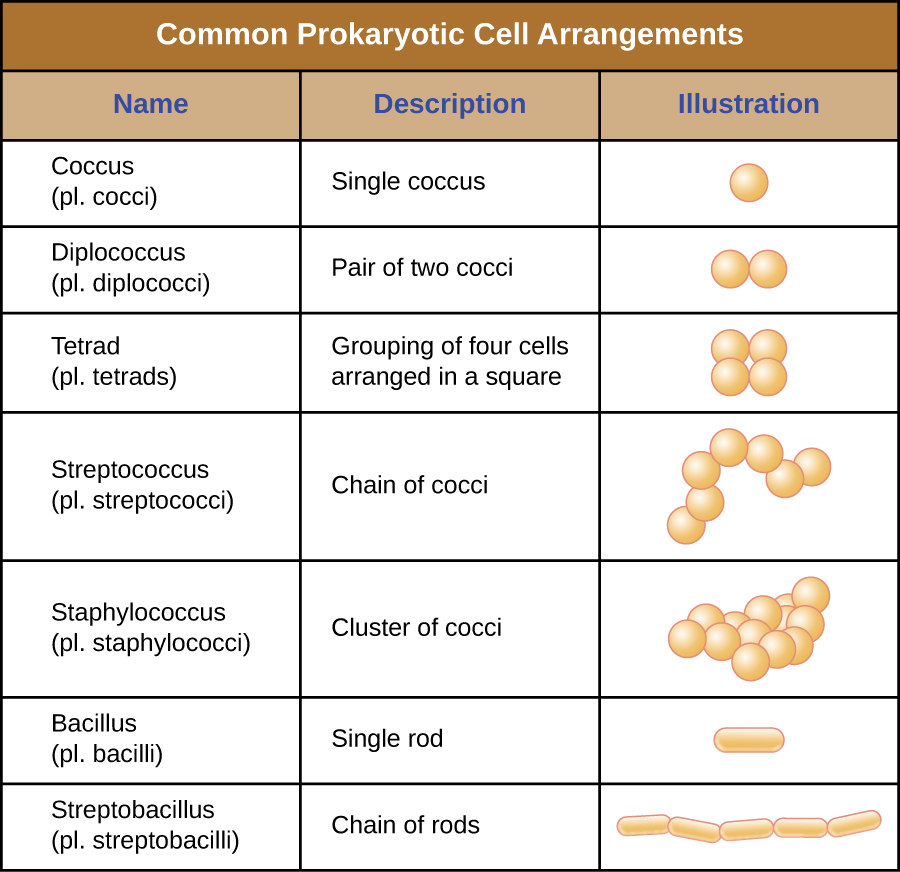
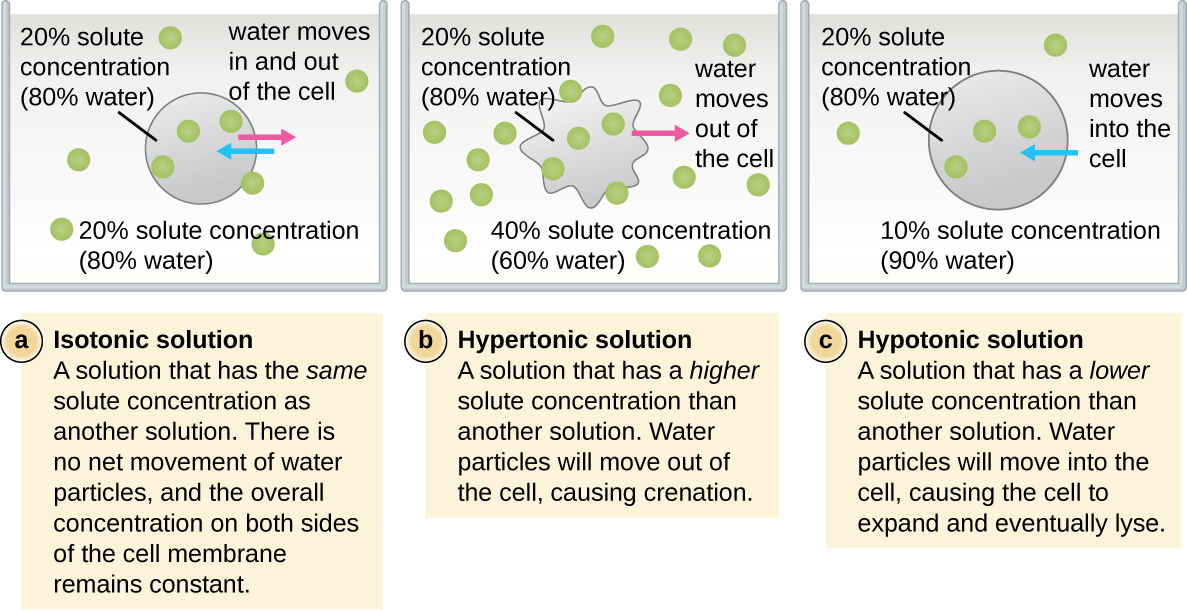
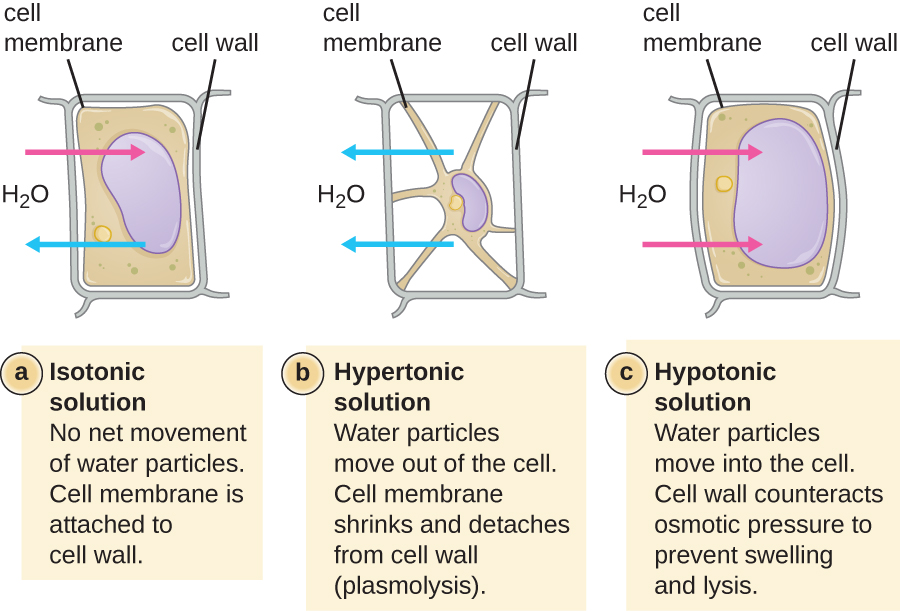
在大多数原核细胞中,形态由细胞壁与细胞骨架元素结合来维持。 细胞壁是大多数原核生物和一些真核生物中存在的一种结构;它包裹细胞膜,保护细胞免受渗透压变化的影响(图\(\PageIndex{4}\))。 渗透压的产生是由于半透膜两侧的溶质浓度存在差异。 水能够穿过半透膜,但溶质(溶解的分子,如盐、糖和其他化合物)不能。 当膜一侧的溶质浓度较高时,水从浓度较低的一侧(更多的水)扩散到浓度较高的一侧(较少的水)穿过膜,直到两侧的浓度变得相等。 这种水的扩散被称为渗透,当细胞的外部环境发生变化时,它会对细胞造成极大的渗透压力。
细胞的外部环境可以描述为等渗介质、高渗介质或低渗介质。 在等渗介质中,细胞内外的溶质浓度大致相等,因此水不会在细胞膜上净流动。 在高渗培养基中,细胞外的溶质浓度超过细胞内部的溶质浓度,因此水会从细胞中扩散到外部培养基中。 在低渗培养基中,细胞内的溶质浓度超过细胞外部的溶质浓度,因此水会通过渗透进入细胞。 这会导致细胞肿胀,并可能裂解或破裂。
特定细胞能够承受渗透压变化的程度称为张力。 有细胞壁的细胞能够更好地承受渗透压的微妙变化并保持其形状。 在高渗环境中,缺乏细胞壁的细胞可能会脱水,从而导致细胞生长或萎缩;质膜收缩并呈扇形或缺口(图\(\PageIndex{4}\))。 相比之下,拥有细胞壁的细胞会发生血浆分解而不是结晶化。 在血浆分解中,质膜收缩并脱离细胞壁,内部体积减少,但细胞壁保持完好无损,从而使细胞能够在一段时间内保持一定的形状和完整性(图\(\PageIndex{5}\))。 同样,缺少细胞壁的细胞在低张环境中更容易发生裂解。 细胞壁的存在使细胞在裂解之前能够在更长的时间内保持其形状和完整性(图\(\PageIndex{5}\))。
练习\(\PageIndex{1}\)
- 解释细胞形态和排列之间的区别。
- 细胞壁为原核细胞提供哪些优势?
核样体
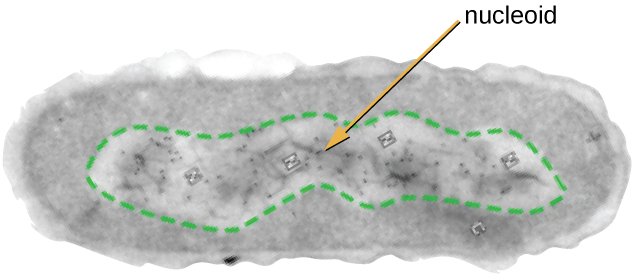
所有细胞生命都有一个组织成一条或多条染色体的DNA基因组。 原核染色体通常是圆形的、单倍体(未配对)的,不受复杂的核膜结合。 原核生物 DNA 和 DNA 相关蛋白集中在细胞的核样区域内(图\(\PageIndex{6}\))。 一般而言,原核生物 DNA 与有助于组织和包装染色体的核酸相关蛋白 (NAP) 相互作用。 在细菌中,NAP 的功能类似于组蛋白,后者是真核细胞中发现的 DNA 组织蛋白。 在古细菌中,核样体由NAP或组蛋白样DNA组织蛋白组织。
质粒
原核细胞还可能含有染色体外的 DNA,或不属于染色体的 DNA。 这种染色体外的 DNA 存在于质粒中,质粒是小的、圆形的双链 DNA 分子。 具有质粒的细胞通常在一个细胞内有数百个质粒。 质粒更常见于细菌中;但是,在古细菌和真核生物中发现了质粒。 质粒通常携带具有优势特征的基因,例如抗生素耐药性;因此,它们对生物体的存活很重要。 我们将在《微生物遗传学机制》中更详细地讨论质粒。
核糖体
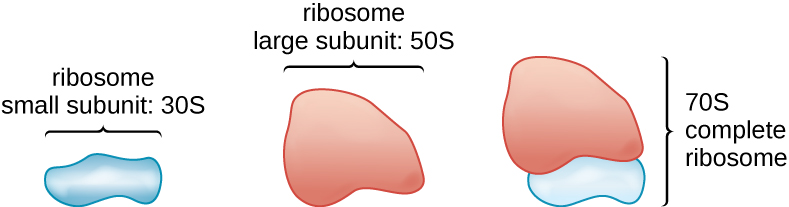
所有细胞生命都会合成蛋白质,而生命所有三个领域的生物都拥有核糖体,核糖体是负责蛋白质合成的结构。 但是,这三个结构域中每个结构域中的核糖体在结构上都不同。 核糖体本身由蛋白质和核糖体RNA(rRNA)构成。 在细胞质中发现了原核生物核糖体。 它们之所以被称为70S核糖体,是因为它们的大小为70S(图\(\PageIndex{7}\)),而真核细胞质核糖体的大小为80S。 (S 代表 Svedberg 单位,用于衡量超离心机中的沉淀量,基于正在分析的结构的大小、形状和表面质量)。 尽管它们的大小相同,但细菌和古生物核糖体具有不同的蛋白质和rRNA分子,而古生物版本与真核生物对应物更相似,而不是与细菌中发现的核糖体相似。
内含物
作为生活在不稳定环境中的单细胞生物,一些原核细胞能够在称为内含物的细胞质结构中储存多余的营养。 以聚合形式储存营养物质是有利的,因为它可以减少细胞积聚溶质时发生的渗透压积聚。 各种类型的内含物会储存糖原和淀粉,其中含有细胞可以获取能量的碳。 Volutin 颗粒,由于其染色特性,也称为变色颗粒,是储存聚合的无机磷酸盐的内含物,可用于新陈代谢并有助于生物膜的形成。 已知含有 volutin 颗粒的微生物包括古细菌 Methan osarcina、Corynebacterium diphtheriae 细菌和单细胞真核藻衣原体单胞菌。 硫颗粒是另一种内含物,存在于硫杆菌属的硫菌中;这些颗粒储存元素硫,细菌将其用于新陈代谢。
有时,某些类型的内含物会被嵌入蛋白质的磷脂单层包围。 聚羟丁酸盐(PHB)可以由芽孢杆菌和假单胞菌物种产生,是显示这种单层结构的内含物的一个例子。 在工业方面,PHB 还被用作生物塑料的可生物降解聚合物的来源。 图中显示了几种不同类型的内含物\(\PageIndex{8}\)。

一些原核细胞还有其他类型的内含物,这些内含物可用于营养储存以外的用途。 例如,一些原核细胞会产生气泡,即蛋白质衬里的小气囊的积聚。 这些气泡允许合成它们的原核细胞改变其浮力,从而调整其在水柱中的位置。 磁定细菌,例如 Magnetospirillum magnetotacticum,含有磁体,磁体是被脂质层包围的磁性氧化铁或硫化铁的内含物。 它们允许细胞沿磁场对齐,从而帮助其移动(图\(\PageIndex{8}\))。 诸如 Anabaena cillric a 之类的蓝细菌和诸如 Halothiobacillus neapolitanus 之类的细菌会产生羧体内含物。 Carboxysomes 由数千个蛋白质亚单位的外壳组成。 它们的内部充满了核糖-1,5-二磷酸羧化酶/加氧酶(RuBisco)和碳酸酐酶。 这两种化合物都用于碳代谢。 一些原核细胞还具有羧酶体,可以在一个位置隔离功能相关的酶。 这些结构被认为是原始细胞器,因为它们将重要的化合物或化学反应分开,就像许多真核细胞器一样。
Endospores
细菌细胞通常被观察为营养细胞,但有些细菌属有能力形成内孢子,这种结构在环境条件不利时本质上保护处于休眠状态的细菌基因组。 Endospores(不要与真菌形成的繁殖孢子混淆)允许一些细菌细胞在没有食物或水、暴露于化学物质、极端温度甚至辐射的情况下长时间存活。 表中\(\PageIndex{1}\)比较了营养细胞和内孢子的特征。
| 植物细胞 | Endospores |
|---|---|
| 对极端温度和辐射敏感 | 耐极端温度和辐射 |
| 革兰氏阳性 | 不要吸收革兰染色,只能吸收特殊的内孢子污渍(参见染色显微镜标本) |
| 正常的水分含量和酶活性 | 脱水;没有代谢活性 |
| 能够活跃生长和新陈代谢 | 休眠;没有生长或代谢活动 |
营养细胞转化为内孢子的过程称为孢子形成,它通常始于营养物质耗尽或环境条件变得不利(图\(\PageIndex{9}\))。 该过程始于在营养细菌细胞中形成隔膜。 隔膜不对称地分裂细胞,将DNA前孢与母细胞分开。 前孢将构成内孢子的核心,本质上是细胞染色体的复制品,通过第二层膜与母细胞分离。 通过在膜之间铺设钙和二吡啶酸层,在前孔周围逐渐形成皮层。 然后在皮层周围形成蛋白质孢子外套,而母细胞的DNA会分解。 随着最外层的 exosporium 的形成,内孢子的进一步成熟会发生。 内孢子在母细胞分解后释放,完成孢子形成。
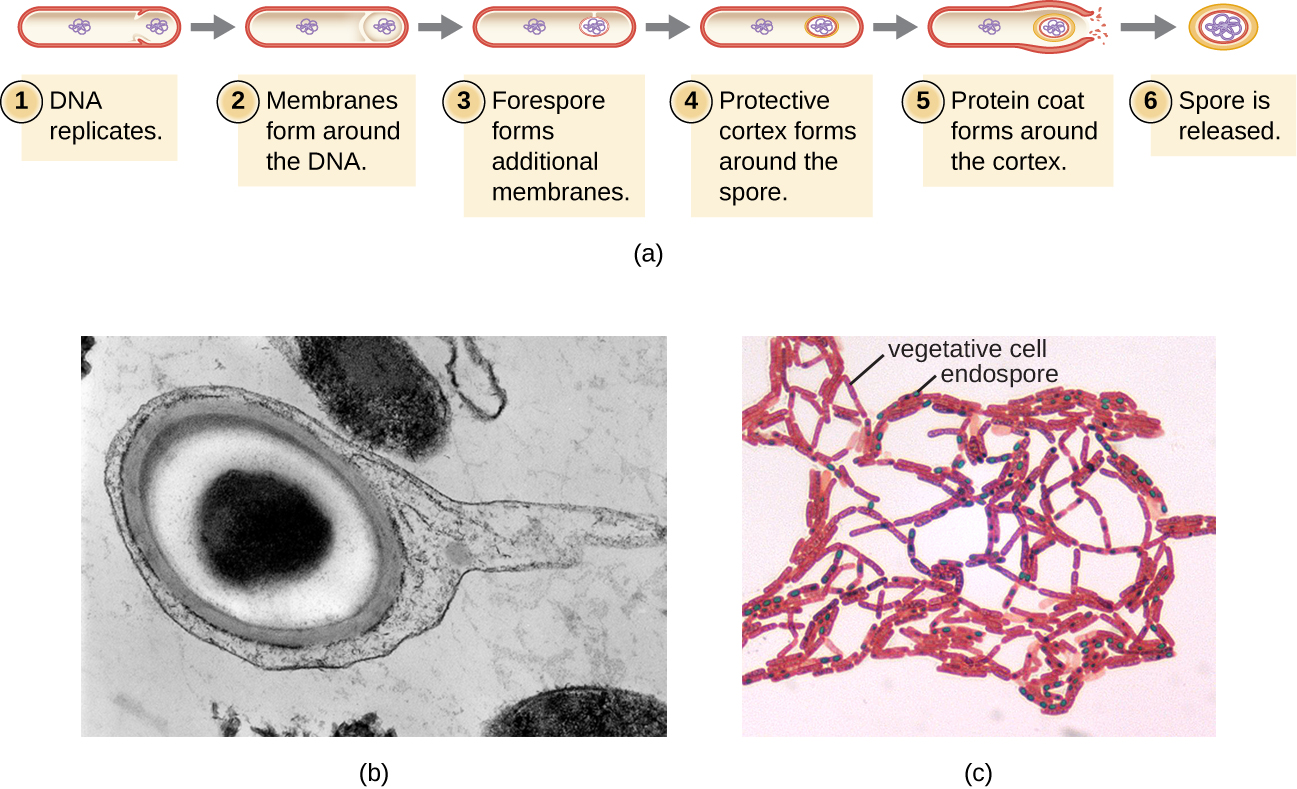
事实证明,某些物种的 Endospores 在休眠状态下会持续很长时间,长达数千年。 2 但是,当生活条件改善时,内孢子会发芽,重新进入植物状态。 发芽后,细胞再次具有新陈代谢活性,并能够发挥其所有正常功能,包括生长和细胞分裂。
并非所有细菌都有能力形成内孢子;但是,有许多具有临床意义的内孢子形成革兰氏阳性细菌,分别是芽孢杆菌和梭菌属。 其中包括炭疽杆菌,炭疽的病原体,它产生的内孢子能够存活数十年 3;破伤风(导致破伤风);艰难梭菌(导致假膜性结肠炎);产气荚膜梭菌(导致气性坏疽);以及C. 肉毒杆菌(导致肉毒中毒)。 诸如此类的病原体特别难以对抗,因为它们的内孢子很难被杀死。 《控制微生物生长》中讨论了内孢形成细菌的特殊灭菌方法。
练习\(\PageIndex{2}\)
- 什么是包容性?
- 内孢子的作用是什么?
质膜
围绕细胞质和细胞内部结构的结构统称为细胞包膜。 在原核细胞中,细胞包膜的结构因细胞和生物体的类型而异。 大多数(但不是全部)原核细胞都有细胞壁,但是这种细胞壁的构成各不相同。 所有细胞(原核细胞和真核细胞)都具有具有选择性渗透性的质膜(也称为细胞质膜或细胞膜),允许某些分子进入或离开细胞,同时限制其他分子的通过。
质膜的结构通常用流体镶嵌模型来描述,该模型是指膜成分在膜平面内流体移动的能力,以及成分的马赛克样成分,其中包括各种脂质和蛋白质成分(图\(\PageIndex{10}\))。 大多数细菌和真核细胞类型的质膜结构是双层,主要由由酯键和蛋白质形成的磷脂组成。 这些磷脂和蛋白质能够在膜平面内以及两个磷脂层之间横向移动。

古生物膜在几个重要方面与细菌和真核生物膜有根本的不同。 首先,古膜磷脂是通过乙醚键形成的,而细菌或真核细胞膜中发现的酯键形成鲜明对比。 其次,古磷脂有分支链,而细菌和真核细胞的磷脂是直链的。 最后,尽管一些古生物膜可以由双层形成,例如在细菌和真核生物中发现的双层,但其他古生物质膜是脂质单层。
细胞表面的蛋白质对多种功能都很重要,包括细胞间的通信、感知环境条件和致病毒力因子。 膜蛋白和磷脂可能含有与之相关的碳水化合物(糖),分别被称为糖蛋白或糖脂。 这些糖蛋白和糖脂复合物从细胞表面向外延伸,使细胞能够与外部环境相互作用(图\(\PageIndex{10}\))。 质膜中的糖蛋白和糖脂在古细菌、细菌和真核生物之间的化学成分可能有很大差异,这使科学家能够使用它们来表征独特的物种。
来自不同细胞类型的血浆膜还含有独特的磷脂,其中含有脂肪酸。 正如《使用生物化学识别微生物》中所述,磷脂衍生脂肪酸分析 (PLFA) 谱可用于根据脂肪酸的差异识别独特类型的细胞。 古细菌、细菌和真核生物都具有独特的 PFLA 特征。
膜传输机制
质膜最重要的功能之一是控制分子进出细胞的运输。 尽管外部环境发生任何变化,内部条件必须保持在一定范围内。 物质穿过质膜的运输允许细胞这样做。
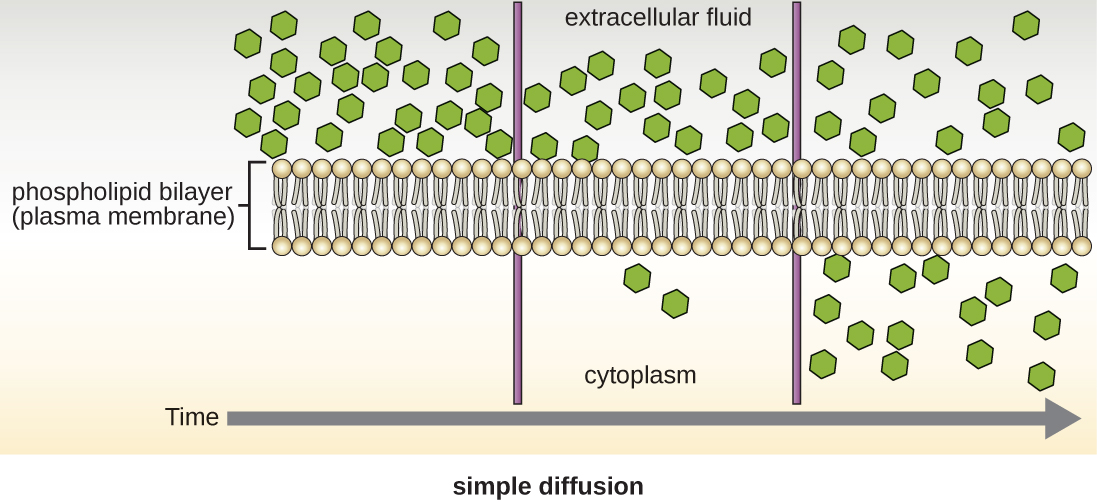
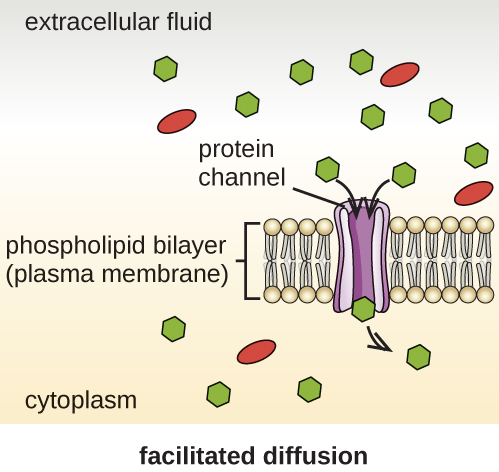
细胞在质膜上使用各种运输方式。 例如,分子通过浓度梯度从较高的浓度转移到较低的浓度,通过简单的扩散(也称为被动传输)进行传输(图\(\PageIndex{11}\))。 一些小分子,例如二氧化碳,可以通过简单的扩散直接穿过膜双层。 但是,带电分子以及大分子都需要膜中载体或通道的帮助。 这些结构将分子运送到膜上,这种过程被称为促进扩散(图\(\PageIndex{12}\))。
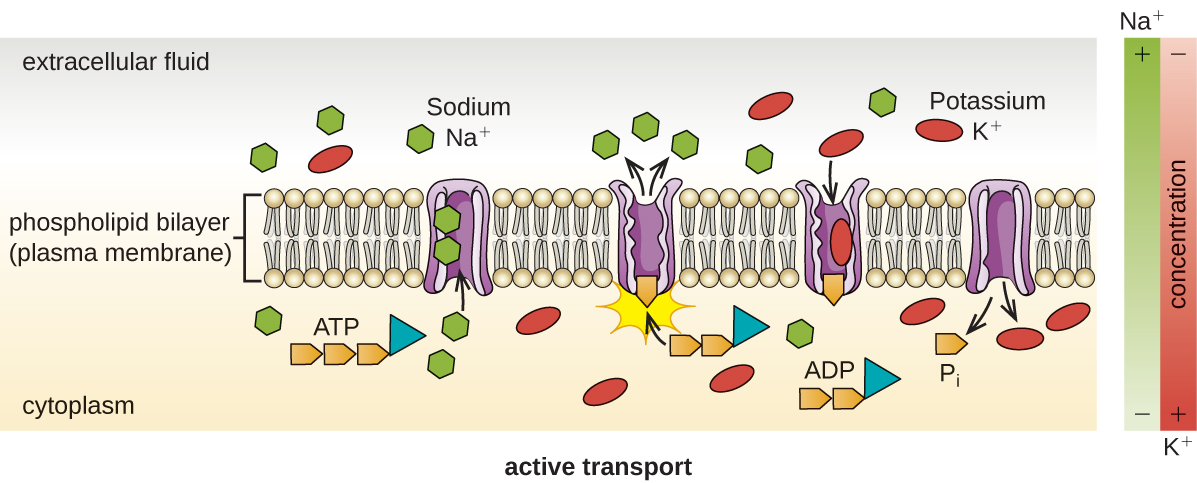
当细胞根据浓度梯度在膜上移动分子时,就会发生主动转运(图\(\PageIndex{13}\))。 被动转运和主动转运的主要区别在于,主动转运需要三磷酸腺苷(ATP)或其他形式的能量才能将分子 “上坡” 移动。 因此,主动运输结构通常被称为 “泵”。
群体转移还会将物质输送到细菌细胞中。 在这种情况下,当分子按照其浓度梯度进入细胞时,它会经过化学修改,因此不需要在不利的浓度梯度下运输。 这方面的一个常见例子是细菌磷酸转移酶系统,这是一系列在进入细胞时磷酸化(即向葡萄糖或其他糖添加磷酸离子)的载体。 由于糖代谢的早期阶段需要对糖进行磷酸化,因此磷酸转移酶系统被认为是一种能量中立系统。
光合膜结构
一些原核细胞,即蓝细菌和光合细菌,具有使它们能够进行光合作用的膜结构。 这些结构由质膜的折叠组成,质膜包围了绿色叶绿素和细菌叶绿素等光合色素。 在蓝细菌中,这些膜结构被称为 thylakoids;在光合细菌中,它们被称为色谱团、薄片或氯体。
细胞墙
细胞壁的主要功能是保护细胞免受外部环境恶劣条件的影响。 当存在时,古细菌、细菌和真核生物的细胞壁之间有明显的相似之处和不同之处。
细菌细胞壁的主要成分被称为 peptidoglycan(或 murein);它仅存在于细菌中。 从结构上讲,peptidoglycan 类似于一层网状或织物(图\(\PageIndex{14}\))。 每层由 N-乙酰氨基葡萄糖 (NAG) 和 N-乙酰氨酸 (NAM) 的长链交替分子组成。 长链的结构具有显著的二维抗拉强度,这是因为在每个肽聚糖层中形成了连接 NAG 和 NAM 的肽桥。 在革兰氏阴性细菌中,从每个 NAM 单位延伸的四肽链直接交联,而在革兰氏阳性细菌中,这些四肽链通过五氨酸交叉桥连接。 Peptidoglycan 亚基在细菌细胞内部制成,然后输出并分层组装,从而形成细胞的形状。
由于肽聚糖是细菌所独有的,因此许多抗生素药物旨在干扰肽聚糖的合成,削弱细胞壁并使细菌细胞更容易受到渗透压的影响(参见抗菌药物的机制)。 此外,人体免疫系统的某些细胞能够通过检测细菌细胞表面的肽聚糖来 “识别” 细菌病原体;然后,这些细胞使用溶菌酶等酶吞没并摧毁细菌细胞,溶菌酶会分解和消化细胞壁中的肽聚糖(参见病原体识别和吞噬作用)。
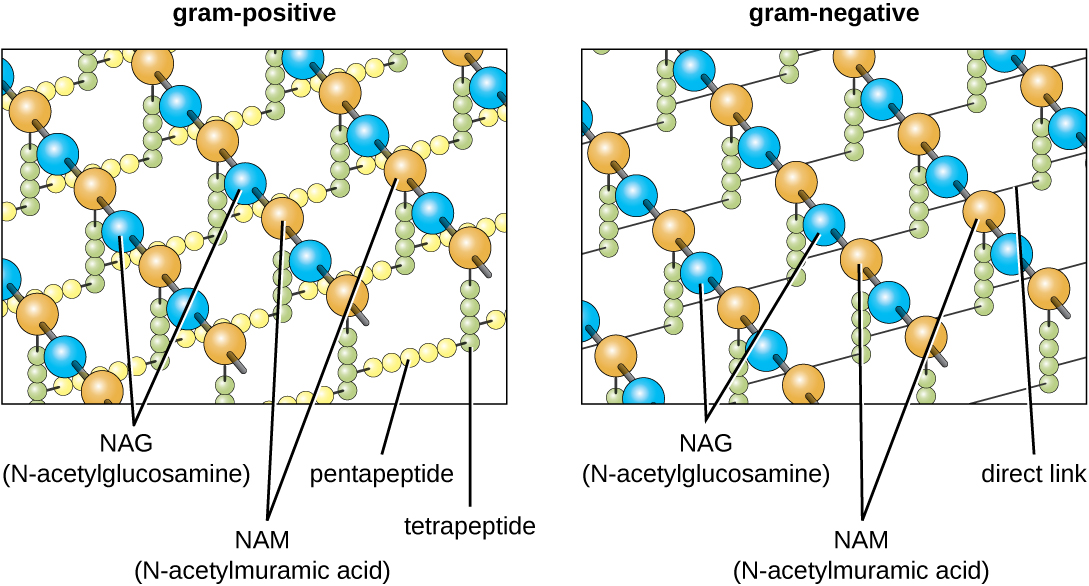
革兰染色方案(参见显微标本染色)用于区分两种常见的细胞壁结构(图\(\PageIndex{15}\))。 革兰阳性细胞的细胞壁由多层肽聚糖组成,总厚度为30—100 nm。 这些 peptidoglycan 层通常嵌入了 teichoic acids (TA),即延伸到肽聚糖层之外的碳水化合物链。 4 人们认为 TA 可以通过增加肽聚糖的硬度来稳定肽聚糖。 TA 还在致病革兰氏阳性细菌(例如链球菌)与宿主细胞表面的某些蛋白质结合的能力中发挥作用,从而增强它们引起感染的能力。 除了 peptidoglycan 和 TA 外,Mycobacteriaceae 家族细菌的细胞壁中还有一层外层的蜡质霉酸;如染色显微标本中所述,这些细菌被称为耐酸,因为必须使用耐酸污渍才能穿透霉菌用于显微镜检查的酸层(图\(\PageIndex{16}\))。

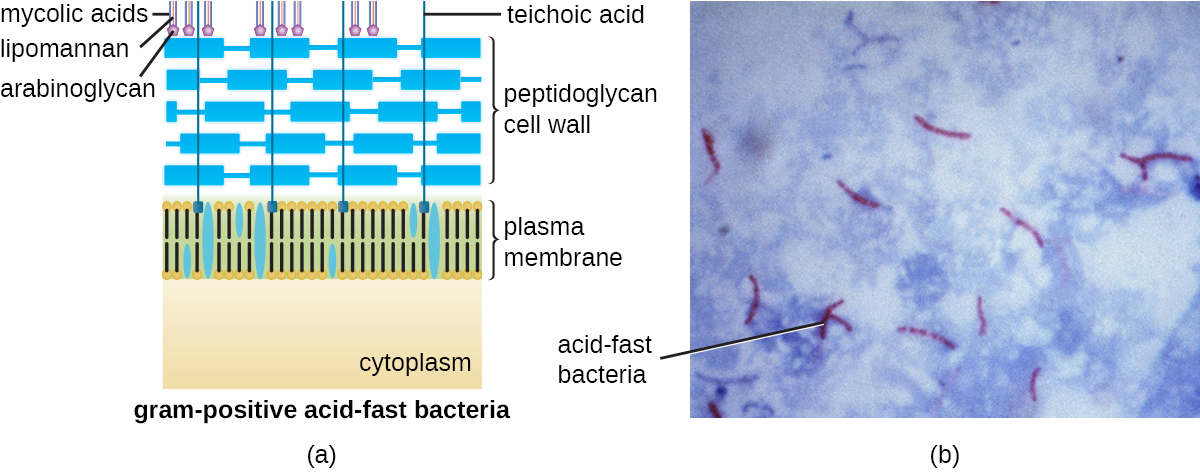
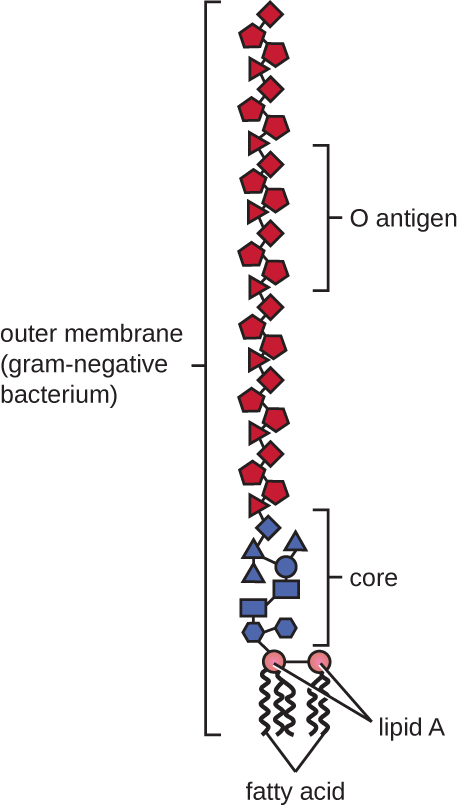
革兰氏阴性细胞的肽聚糖层比革兰阳性细胞薄得多(厚度不超过大约 4 nm 6),其细胞包膜的整体结构也更为复杂。 在革兰氏阴性细胞中,凝胶状基质占据细胞壁和质膜之间的周质空间,还有第二个脂质双层称为外膜,位于肽聚糖层的外部(图\(\PageIndex{15}\))。 这种外膜通过 murein 脂蛋白附着在肽聚糖上。 外膜的外叶含有脂多糖(LPS)分子,它在涉及革兰氏阴性细菌的感染中起到内毒素的作用,会导致发热、出血和感染性休克等症状。 每个 LPS 分子都由脂质 A(一种核心多糖)和一个 O 侧链组成,后者由构成脂多糖外表面的糖状分子组成(图\(\PageIndex{17}\))。 O 侧链的组成因细菌的不同种类和菌株而异。 O侧链中称为抗原的部分可以通过血清学或免疫学检测来鉴定特定的致病菌株,例如大肠杆菌O157:H7,这是一种导致血性腹泻和肾衰竭的致命细菌菌株。
古细胞壁结构与细菌的细胞壁结构在几个重要方面有所不同。 首先,古细胞壁不含肽聚糖;相反,它们含有一种叫做 pseudopeptidoglycan(pseudomurein)的类似聚合物,其中 NAM 被不同的亚基所取代。 其他古细菌可能有一层糖蛋白或多糖作为细胞壁,而不是伪肽聚糖。 最后,与某些细菌物种一样,有一些古细菌似乎完全缺乏细胞壁。
Glycocalyces 和 S 层
尽管大多数原核细胞都有细胞壁,但有些原核细胞在细胞壁外可能有额外的细胞包膜结构,例如糖酵素和S层。 glycocalyx 是一种糖衣,其中有两种重要的类型:胶囊和粘液层。 胶囊是位于细胞壁外的有组织层,通常由多糖或蛋白质组成(图\(\PageIndex{18}\))。 粘液层是一种组织不那么紧密的层,它只能松散地附着在细胞壁上,更容易被洗掉。 粘液层可能由多糖、糖蛋白或糖脂组成。
Glycocalyces 允许细胞粘附在表面,帮助形成生物膜(在表面层中形成的微生物菌落)。 在自然界中,大多数微生物生活在生物膜中的混合群落中,部分原因是生物膜为它们提供了一定程度的保护。 生物膜通常像海绵一样吸水,防止干燥。 它们还可以保护细胞免受捕食,阻碍抗生素和消毒剂的作用。 所有这些特性对生活在生物膜中的微生物都有利,但是它们在临床环境中带来了挑战,因为临床环境的目标通常是消灭微生物。

生产胶囊的能力会增加微生物的致病性(致病能力),因为胶囊会使吞噬细胞(例如白细胞)更难吞没和杀死微生物。 例如,肺炎链球菌会产生一种胶囊,众所周知,这种胶囊有助于该细菌的致病性。 如显微镜标本染色中所述,胶囊很难在显微镜检查中染色;通常使用阴性染色技术。
S 层是另一种类型的细胞包膜结构;它由结构蛋白和糖蛋白的混合物组成。 在细菌中,S层存在于细胞壁外,但在某些古细菌中,S层充当细胞壁。 S 层的确切功能尚不完全清楚,也很难研究;但现有证据表明,它们可能在不同的原核细胞中发挥各种功能,例如帮助细胞承受渗透压,对于某些病原体,它们与宿主免疫相互作用系统。
临床重点:第 3 部分
在诊断出芭芭拉患有肺炎后,巴勒斯坦权力机构给她开了阿莫西林的处方,阿莫西林是一种常用的青霉素衍生物。 一个多星期后,尽管芭芭拉按照指示完成了全部课程,但她仍然感到虚弱,没有完全康复,尽管她仍然能够完成日常活动。 她回到健康中心进行后续检查。
许多类型的细菌、真菌和病毒都可能导致肺炎。 阿莫西林靶向细菌细胞壁的肽聚糖。 由于阿莫西林尚未解决芭芭拉的症状,巴勒斯坦权力机构得出结论,该病原体可能缺乏肽聚糖,这意味着病原体可能是病毒、真菌或缺乏肽聚糖的细菌。 另一种可能性是病原体是一种含有肽聚糖的细菌,但对阿莫西林产生了耐药性。
练习\(\PageIndex{3}\)
- 巴勒斯坦权力机构如何确定芭芭拉肺炎的病因?
- 鉴于阿莫西林无效,巴勒斯坦权力机构应该开什么形式的治疗方案?
丝状附属物
许多细菌细胞的细胞包膜内嵌有向外延伸的蛋白质附属物,允许与环境相互作用。 这些附属物可以附着在其他表面、转移 DNA 或提供运动。 丝状附属物包括纤维、绒毛和鞭毛。
Fimbriae 和 Pili
Fimbriae 和 pili 在结构上相似,由于两者之间的区别存在问题,因此这两个术语通常可以互换使用。 7 8 fimbriae 一词通常是指从细胞表面投射出数百个短的刷毛样蛋白。 Fimbriae 使细胞能够附着在表面和其他细胞上。 对于致病细菌,对宿主细胞的粘附对于定植、传染性和毒性很重要。 粘附在表面对生物膜的形成也很重要。
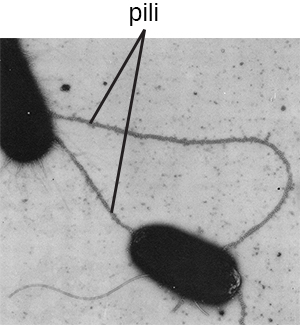
术语 pili(单数:pilus)通常是指有助于附着在表面的较长、数量较少的蛋白质附属物(图\(\PageIndex{19}\))。 一种特殊类型的 pilus,称为 F pilus 或 sex pilus,对于细菌细胞之间的 DNA 转移很重要。当两个细胞物理转移或交换各自基因组的一部分时,细菌细胞之间会发生这种转移(参见 Asexual Prokaryotes 如何实现遗传)多样性)。
在充分了解细菌细胞包膜各种成分的结构和功能之前,科学家们已经在使用细胞包膜特征对细菌进行分类。 1933年,丽贝卡·兰斯菲尔德(Rebecca Lancefield)提出了一种使用凝集测定法对链球菌种的各种β-溶血菌菌株进行血清分型的方法,该技术使用细菌团块来检测特定的细胞表面抗原。 在这样做的过程中,兰斯菲尔德发现,在A组中发现的一组化脓杆菌与各种人类疾病有关。 她确定,根据她命名为M蛋白的特定细胞表面蛋白的变异,可以将各种A组链球菌菌株相互区分。
如今,已经鉴定出基于M蛋白的80多种不同的A组链球菌菌株。 各种 A 组链球菌菌株与各种人类感染有关,包括链球菌咽炎(链球菌性喉炎)、脓疱病、中毒性休克综合征、猩红热、风湿热和坏死性筋膜炎。 M 蛋白是 A 组链球菌的重要毒力因子,可帮助这些菌株逃避免疫系统。 M 蛋白的变化似乎改变了特定 A 组链球菌菌株的传染性。
鞭毛
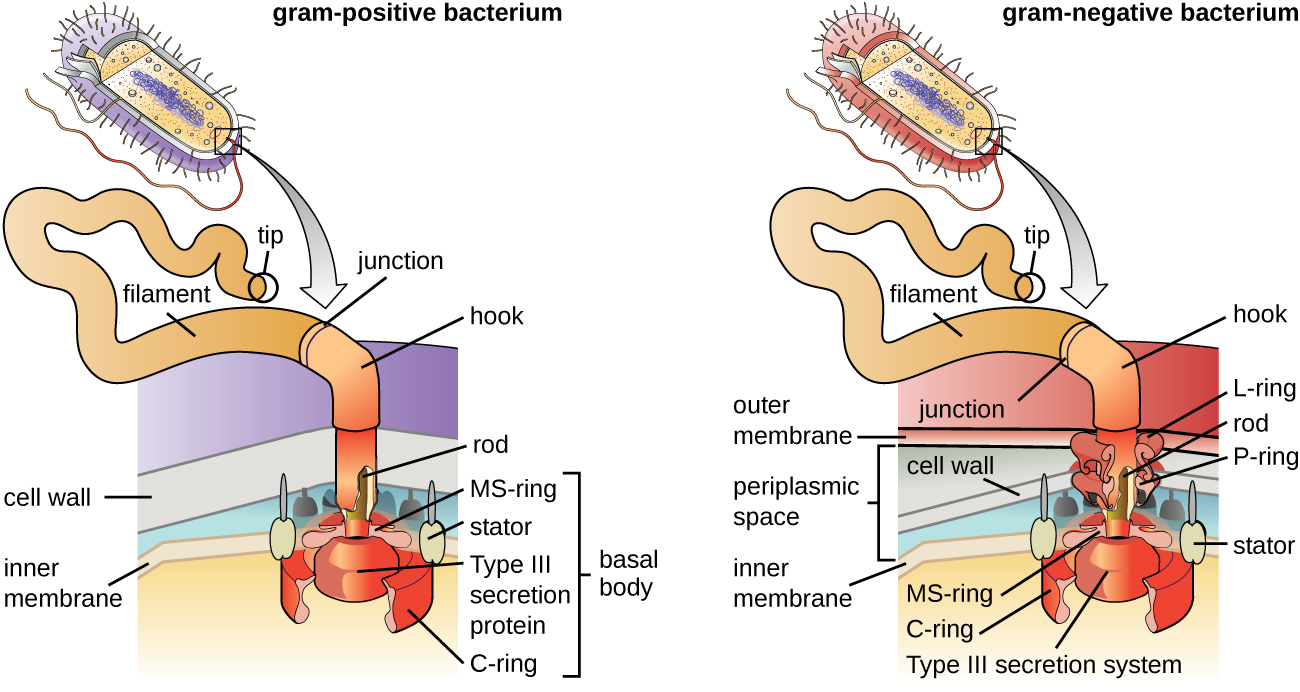
鞭毛是细胞用来在水环境中移动的结构。 细菌鞭毛就像螺旋桨一样。 它们是由鞭毛蛋白亚基组成的坚硬螺旋丝,从细胞向外延伸并在溶液中旋转。 基体是鞭毛的发动机,嵌入在质膜中(图\(\PageIndex{20}\))。 钩形区域将基体连接到灯丝。 由于细胞壁结构的不同,革兰氏阳性和革兰氏阴性细菌具有不同的基体构型。
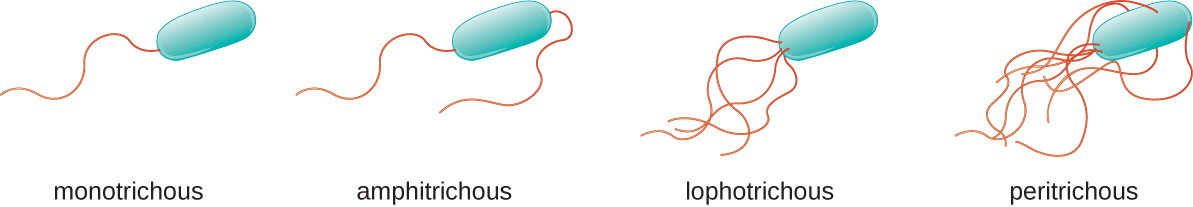
不同类型的运动细菌表现出不同的鞭毛排列(图\(\PageIndex{21}\))。 据说具有单一鞭毛的细菌通常位于细胞的一端(极性),具有单色鞭毛。 霍乱弧菌是导致霍乱的革兰氏阴性细菌,这是一种单丝鞭打的细菌病原体。 带有两栖鞭毛的细胞两端都有一根或一簇鞭毛。 例如 Spirillum minor,它是螺旋体(亚洲)老鼠咬伤热或 sodoku 的原因。 带有 lophotrichous 鞭毛的细胞在细胞的一端有一簇状物。 革兰氏阴性芽孢杆菌铜绿假单胞菌是一种机会主义病原体,以引起多种感染(包括 “游泳者的耳朵” 和烧伤感染)而闻名,它有环状鞭毛。 覆盖细菌细胞整个表面的鞭毛被称为 peritrichous 鞭毛。 革兰氏阴性细菌大肠杆菌呈现出周围的鞭毛排列。
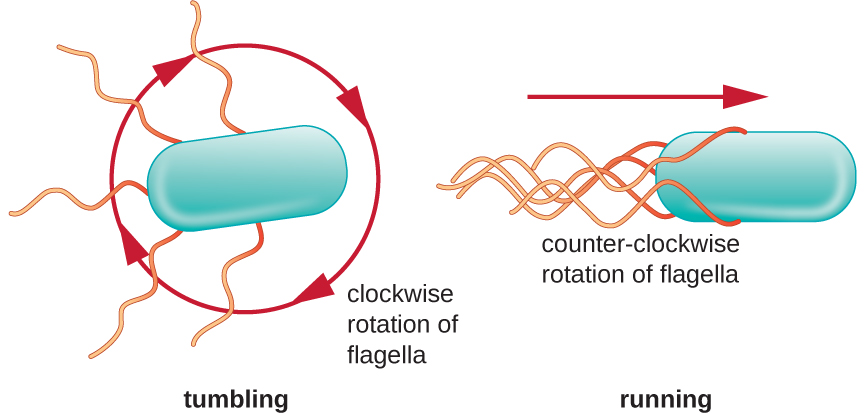
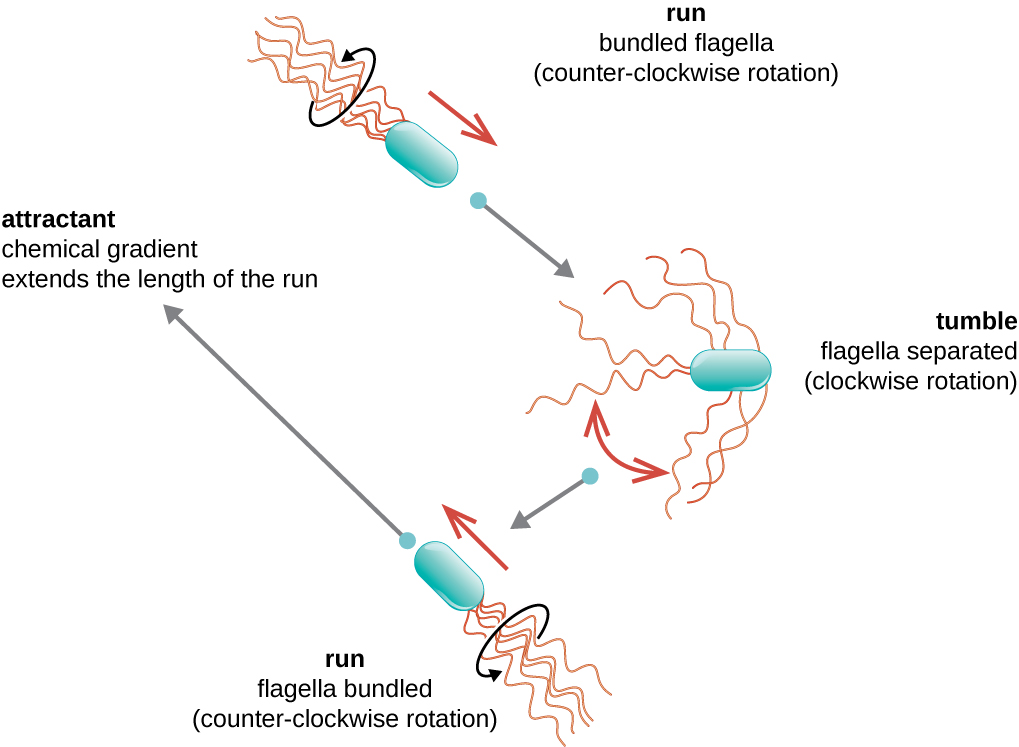
定向运动取决于鞭毛的配置。 细菌可以响应各种环境信号而移动,包括光(光性)、使用磁体的磁场(磁性)以及最常见的化学梯度(趋化性)。 有目的地向化学引诱剂(如食物来源)或驱虫剂(如有毒化学物质)移动,是通过增加跑步时间和缩短跌落时间来实现的。 跑步时,鞭毛沿逆时针方向旋转,使细菌细胞向前移动。 在有害细菌中,鞭毛以非常流线型的方式捆绑在一起(图\(\PageIndex{22}\)),可以高效移动。 翻滚时,鞭毛在顺时针旋转的同时会张开,从而产生循环运动,阻止有意义的向前移动,但会使细胞朝引物的方向重新定向。 当引诱剂存在时,仍然会发生奔跑和翻倒;但是,跑步时间会更长,而翻滚的长度会缩短,从而允许整体向更高浓度的引诱剂移动。 当不存在化学梯度时,跑步和翻滚的长度会更加相等,整体移动也更加随机(图\(\PageIndex{23}\))。
练习\(\PageIndex{4}\)
- 什么是肽聚糖层?革兰氏阳性和革兰氏阴性细菌之间有何区别?
- 比较和对比 monotrichous、amphitrichous、lophotrichous 和 peritrichous 鞭毛。
摘要
- 原核细胞与真核细胞的不同之处在于,它们的遗传物质包含在核样体中,而不是膜结合的细胞核中。 此外,原核细胞通常缺乏膜结合的细胞器。
- 相同物种的原核细胞通常具有相似的细胞形态和细胞排列。
- 大多数原核细胞都有细胞壁,可以帮助生物体维持细胞形态并保护其免受渗透压变化的影响。
- 在核样体之外,原核细胞在质粒中可能含有染色体外的 DNA。
- 在细胞质中发现的原核生物核糖体的大小为70S。
- 一些原核细胞含有内含物,可以储存营养物质或化学物质以用于其他用途。
- 一些原核细胞能够通过孢子形成形成内孢子,在条件不利时在休眠状态下存活。 Endospores 可以发芽,条件改善后会转化为营养细胞。
- 在原核细胞中,细胞包膜包括质膜,通常是细胞壁。
- 细菌膜由具有整体或外周蛋白的磷脂组成。 这些磷脂的脂肪酸成分是酯连接的,通常用于鉴定特定类型的细菌。 这些蛋白具有多种功能,包括运输、细胞间通信和感知环境条件。 古生物膜的不同之处在于它们由脂肪酸组成,这些脂肪酸与磷脂有乙醚连接。
- 有些分子可以通过简单的扩散在细菌膜上移动,但大多数大分子必须使用细胞能量通过膜结构主动运输。
- 原核细胞壁可能由肽聚糖(细菌)或伪肽聚糖(古细菌)组成。
- 革兰氏阳性细菌细胞的特征是肽聚糖层较厚,而革兰氏阴性细菌细胞的特征是被外膜包围的肽聚糖层很薄。
- 一些原核细胞会产生糖萼涂层,例如胶囊和粘液层,这有助于附着在表面和/或逃避宿主免疫系统。
- 一些原核细胞有 f imbriae 或 pili,即有助于附着在表面的丝状附属物。 Pili 还用于在细胞之间转移遗传物质。
- 一些原核细胞使用一根或多根鞭毛在水中移动。 Peritrichous 细菌有许多鞭毛,它们利用奔跑和翻滚故意朝化学引诱剂的方向移动。
脚注
- 1 Y.-H.M. 陈,W.F. Marshall。 “细胞和细胞器大小的缩放特性。” Organogenesis 6 no.2 (2010): 88—96。
- 2 F. Rothfuss、M Bender、R Conrad。 “深层老化湖泊沉积物(博登湖)中细菌的存活和活性。” 微生物生态学 33 第 1 期 (1997): 69—77。
- 3 R. Sinclair 等人。 “A 类精选代理在环境中的持久性。” 应用与环境微生物学 74 第 3 期 (2008): 555—563。
- 4 T.J. Silhavy、D. Kahne、S. Walker。 “细菌细胞包膜。” 《冷泉港生物学展望》第 2 期第 5 期 (2010): a000414。
- 5 B. Zuber 等人。 “玻璃体切片的低温电子显微镜揭示了革兰氏阳性细菌的周质空间中的颗粒层以及肠球菌 gallinarum 和 Streptoccus gordonii Septa 的精细结构。” 《细菌学杂志》 188 第 18 期 (2006): 6652—6660
- 6 L. Gana、S. Chena、G.J. Jensena。 “革兰氏阴性肽聚糖的分子组织。” 美利坚合众国国家科学院院刊 105 第 48 期 (2008): 18953—18957。
- 7 J.A. Garnetta 等人 “大肠杆菌 Comm on Pilus 对生物发生和生物膜形成的结构见解。” 《美利坚合众国国家科学院院刊》 109 第 10 期 (2012): 3950—3955。
- 8 T. Proft,E.N. Baker。 “革兰氏阴性和革兰氏阳性细菌中的Pili——结构、组装及其在疾病中的作用。” 细胞和分子生命科学 66 (2009): 613。