
8.6 : La photosynthèse et l'importance de la lumière
- Page ID
- 187719

Objectifs d'apprentissage
- Décrire la fonction et l'emplacement des pigments photosynthétiques chez les eucaryotes et les procaryotes
- Décrire les principaux produits des réactions dépendantes et indépendantes de la lumière
- Décrire les réactions qui produisent du glucose dans une cellule photosynthétique
- Comparer et contraster la photophosphorylation cyclique et non cyclique
Les organismes hétérotrophes, qu'il s'agisse d'E. coli ou d'humains, dépendent de l'énergie chimique présente principalement dans les molécules de glucides. Bon nombre de ces glucides sont produits par photosynthèse, le processus biochimique par lequel les organismes phototrophes convertissent l'énergie solaire (lumière du soleil) en énergie chimique. Bien que la photosynthèse soit le plus souvent associée aux plantes, la photosynthèse microbienne est également un important fournisseur d'énergie chimique, alimentant de nombreux écosystèmes divers. Dans cette section, nous allons nous concentrer sur la photosynthèse microbienne.
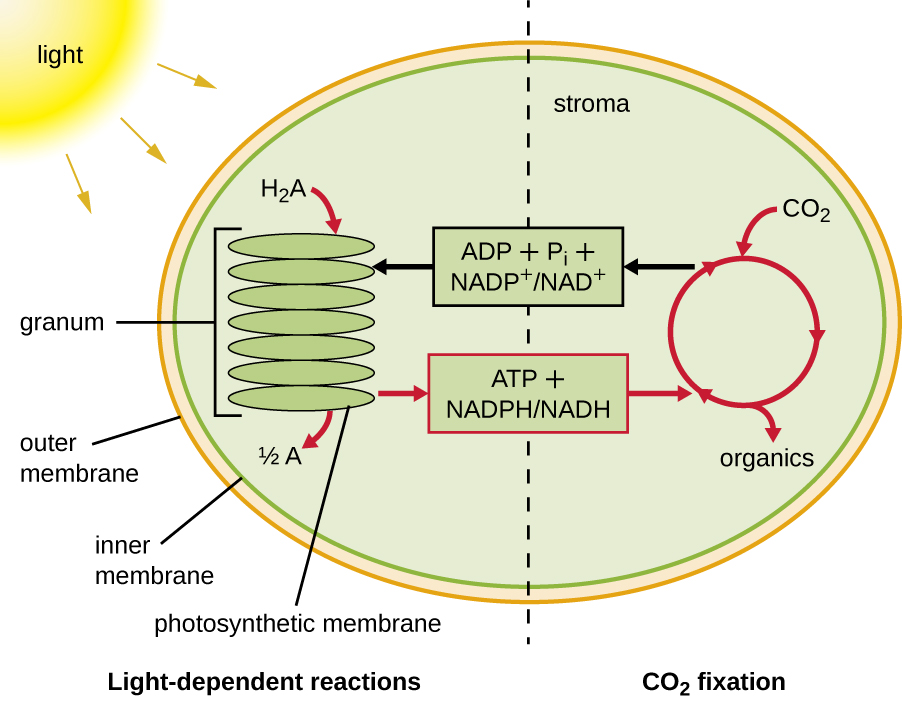
La photosynthèse se déroule en deux étapes séquentielles : les réactions dépendantes de la lumière et les réactions indépendantes de la lumière (Figure\(\PageIndex{1}\)). Dans les réactions dépendantes de la lumière, l'énergie du soleil est absorbée par les molécules pigmentaires des membranes photosynthétiques et convertie en énergie chimique stockée. Dans les réactions indépendantes de la lumière, l'énergie chimique produite par les réactions dépendantes de la lumière est utilisée pour entraîner l'assemblage de molécules de sucre à l'aide de CO 2 ; toutefois, ces réactions dépendent toujours de la lumière car les produits des réactions dépendantes de la lumière nécessaires à leur entraînement sont courts -vécu. Les réactions dépendantes de la lumière produisent de l'ATP et du NADPH ou du NADH pour stocker temporairement de l'énergie. Ces vecteurs énergétiques sont utilisés dans les réactions indépendantes de la lumière pour entraîner le processus défavorable sur le plan énergétique consistant à « fixer » le CO 2 inorganique sous une forme organique, le sucre.
Structures photosynthétiques chez les eucaryotes et les procaryotes
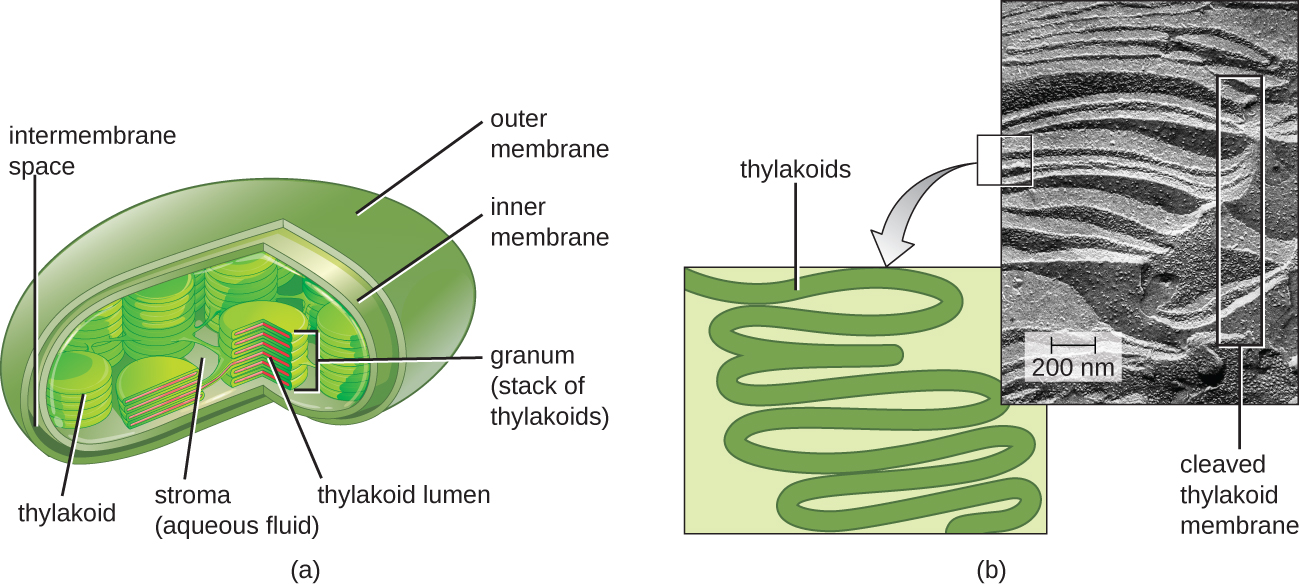
Chez tous les eucaryotes phototrophes, la photosynthèse a lieu à l'intérieur d'un chloroplaste, un organite créé chez les eucaryotes par endosymbiose d'une bactérie photosynthétique (voir Caractéristiques uniques des cellules eucaryotes). Ces chloroplastes sont entourés d'une double membrane avec des couches interne et externe. À l'intérieur du chloroplaste se trouve une troisième membrane qui forme des structures photosynthétiques empilées en forme de disque appelées thylakoïdes (Figure\(\PageIndex{2}\)). Un empilement de thylakoïdes est appelé granum, et l'espace entourant le granum dans le chloroplaste est appelé stroma.
Les membranes photosynthétiques des procaryotes, au contraire, ne sont pas organisées en organites distincts entourés de membranes ; il s'agit plutôt de régions repliées de la membrane plasmique. Dans les cyanobactéries, par exemple, ces régions repliées sont également appelées thylakoïdes. Dans les deux cas, les molécules de pigments photosynthétiques sont intégrées dans les membranes thylakoïdes ou dans d'autres membranes bactériennes photosynthétiques. Ces molécules sont organisées en un ou plusieurs photosystèmes, où l'énergie lumineuse est convertie en énergie chimique.
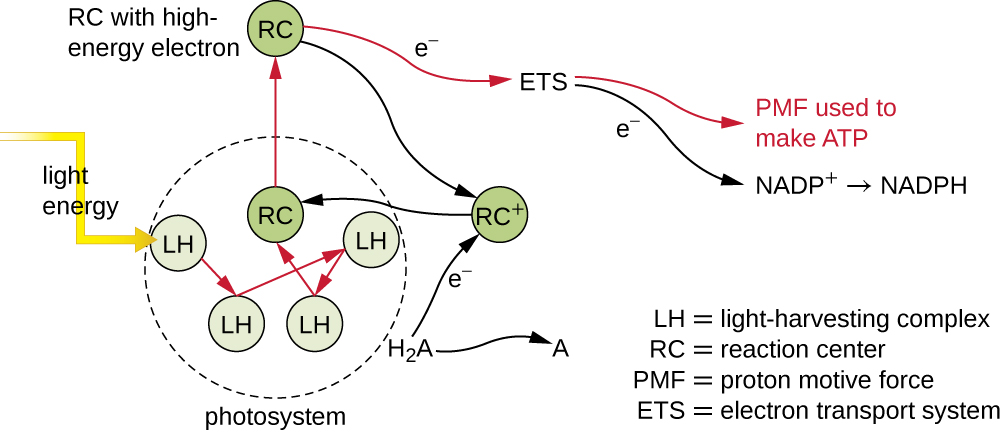
Les pigments photosynthétiques contenus dans les membranes photosynthétiques sont organisés en photosystèmes, chacun d'eux étant composé d'un complexe récupérateur de lumière (antennes) et d'un centre de réaction. Le complexe récupérateur de lumière est constitué de plusieurs protéines et de pigments associés qui peuvent chacun absorber l'énergie lumineuse et, par conséquent, s'exciter. Cette énergie est transférée d'une molécule de pigment à une autre jusqu'à ce qu'elle soit finalement (environ un millionième de seconde) envoyée au centre de réaction. Jusqu'à présent, seule l'énergie, et non les électrons, a été transférée entre les molécules. Le centre de réaction contient une molécule de pigment qui peut s'oxyder lors de l'excitation, libérant ainsi un électron. C'est à cette étape de la photosynthèse que l'énergie lumineuse est convertie en électron excité.
Différents types de pigments collecteurs de lumière absorbent des motifs uniques de longueurs d'onde (couleurs) de la lumière visible. Les pigments réfléchissent ou transmettent les longueurs d'onde qu'ils ne peuvent pas absorber, ce qui leur donne la couleur correspondante. Des exemples de pigments photosynthétiques (molécules utilisées pour absorber l'énergie solaire) sont les bactériochlorophylles (vertes, violettes ou rouges), les caroténoïdes (orange, rouge ou jaune), les chlorophylles (vertes), les phycocyanines (bleues) et les phycoérythrines (rouges). En ayant des mélanges de pigments, un organisme peut absorber de l'énergie provenant de plusieurs longueurs d'onde. Comme les bactéries photosynthétiques se développent généralement en compétition pour la lumière du soleil, chaque type de bactérie photosynthétique est optimisé pour récolter les longueurs d'onde de lumière auxquelles il est généralement exposé, ce qui entraîne une stratification des communautés microbiennes dans les écosystèmes aquatiques et pédologiques en fonction de la qualité de la lumière et de la pénétration.
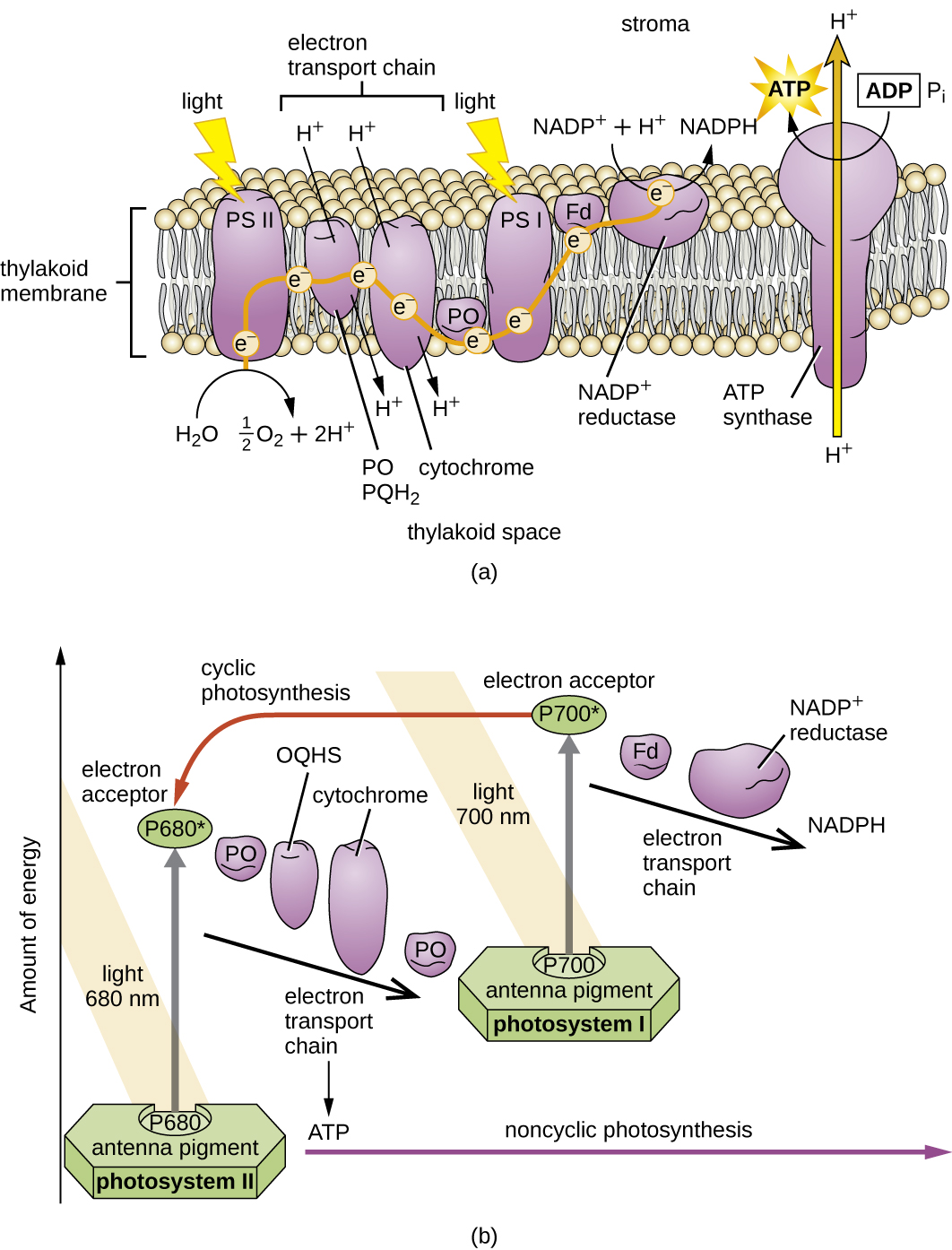
Une fois que le complexe de collecte de lumière transfère l'énergie au centre de réaction, le centre de réaction délivre ses électrons de haute énergie, un par un, à un porteur d'électrons dans un système de transport d'électrons, et le transfert d'électrons à travers l'ETS est initié. L'ETS est similaire à celui utilisé pour la respiration cellulaire et est intégré à la membrane photosynthétique. En fin de compte, l'électron est utilisé pour produire du NADH ou du NADPH. Le gradient électrochimique qui se forme à travers la membrane photosynthétique est utilisé pour générer de l'ATP par chimiosmose par le biais du processus de photophosphorylation, un autre exemple de phosphorylation oxydative (Figure\(\PageIndex{3}\)).
Exercice\(\PageIndex{1}\)
Où se déroule la photosynthèse chez un eucaryote phototrophe ?
Photosynthèse oxygénique et anoxygénique
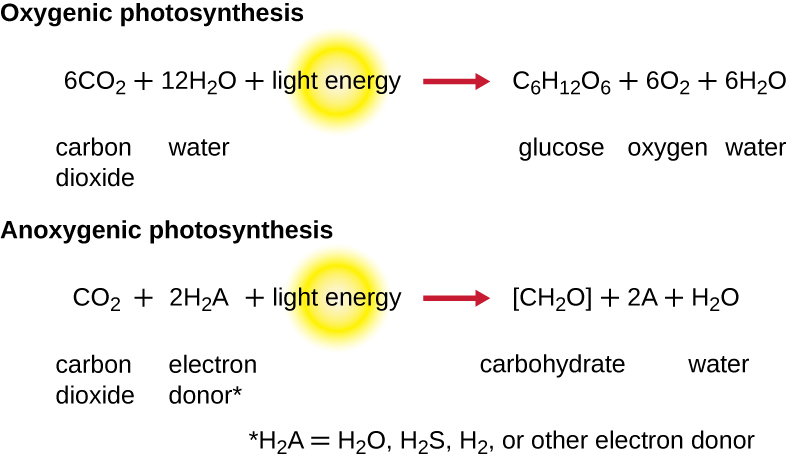
Pour que la photosynthèse se poursuive, l'électron perdu par le pigment du centre de réaction doit être remplacé. La source de cet électron (H 2 A) permet de différencier la photosynthèse oxygénée des plantes et des cyanobactéries de la photosynthèse anoxygénique réalisée par d'autres types de phototrophes bactériens (Figure\(\PageIndex{4}\)). Lors de la photosynthèse oxygénée, le H 2 O est divisé et fournit l'électron au centre de réaction. Comme l'oxygène est généré en tant que sous-produit et est libéré, ce type de photosynthèse est appelé photosynthèse oxygénée. Cependant, lorsque d'autres composés réduits servent de donneurs d'électrons, l'oxygène n'est pas généré ; ces types de photosynthèse sont appelés photosynthèse anoxygénique. Le sulfure d'hydrogène (H 2 S) ou le thiosulfate\(\ce{(S2O3^{2-})}\) peuvent servir de donneur d'électrons, générant ainsi\(\ce{(SO4^{2-})}\) des ions soufre et sulfate élémentaires, respectivement.
Les photosystèmes ont été classés en deux types : le photosystème I (PSI) et le photosystème II (PSII) (Figure\(\PageIndex{5}\)). Les cyanobactéries et les chloroplastes végétaux possèdent les deux photosystèmes, tandis que les bactéries photosynthétiques anoxygènes n'utilisent qu'un seul des photosystèmes. Les deux photosystèmes sont excités simultanément par l'énergie lumineuse. Si la cellule a besoin à la fois d'ATP et de NADPH pour la biosynthèse, elle effectuera une photophosphorylation non cyclique. Lors du passage de l'électron du centre de réaction PSII à l'ETS qui relie PSII et PSI, l'électron perdu du centre de réaction PSII est remplacé par le fractionnement de l'eau. L'électron du centre de réaction PSI excité est utilisé pour réduire le NADP + en NADPH et est remplacé par l'électron sortant de l'ETS. Le flux d'électrons de cette manière est appelé schéma Z.
Si les besoins d'une cellule en ATP sont significativement supérieurs à ses besoins en NADPH, elle peut contourner la production de puissance réductrice par photophosphorylation cyclique. Seul le PSI est utilisé lors de la photophosphorylation cyclique ; l'électron de haute énergie du centre de réaction PSI est transmis à un support ETS, puis retourne finalement au pigment du centre de réaction PSI oxydé, le réduisant ainsi.
Exercice\(\PageIndex{2}\)
Pourquoi une bactérie photosynthétique aurait-elle des pigments différents ?
réactions indépendantes de la lumière
Une fois que l'énergie du soleil est convertie en énergie chimique et stockée temporairement dans des molécules d'ATP et de NADPH (dont la durée de vie est de millionièmes de seconde), les photoautotrophes disposent du carburant nécessaire pour construire des molécules de glucides multicarbonées, capables de survivre pendant des centaines de millions d'années, à long terme stockage d'énergie. Le carbone provient du CO 2, le gaz qui est un déchet de la respiration cellulaire.
Le cycle de Calvin-Benson (nommé en l'honneur de Melvin Calvin [1911—1997] et Andrew Benson [1917—2015]), la voie biochimique utilisée pour la fixation du CO 2, est localisé dans le cytoplasme des bactéries photosynthétiques et dans le stroma des chloroplastes eucaryotes. Les réactions indépendantes de la lumière du cycle de Calvin peuvent être organisées en trois étapes de base : fixation, réduction et régénération (voir l'annexe C pour une illustration détaillée du cycle de Calvin).
- Fixation : L'enzyme ribulose bisphosphate carboxylase (RuBisCO) catalyse l'ajout d'un CO 2 au ribulose bisphosphate (RuBP). Il en résulte la production de 3-phosphoglycérate (3-PGA).
- Réduction : Six molécules d'ATP et de NADPH (issues de réactions dépendantes de la lumière) sont utilisées pour convertir le 3-PGA en glycéraldéhyde 3-phosphate (G3P). Une partie du G3P est ensuite utilisée pour produire du glucose.
- Régénération : Le G3P restant non utilisé pour synthétiser le glucose est utilisé pour régénérer le RuBP, ce qui permet au système de poursuivre la fixation du CO 2. Trois autres molécules d'ATP sont utilisées dans ces réactions de régénération.
Le cycle de Calvin est largement utilisé par les plantes et les bactéries photoautotrophes, et l'enzyme RubisCO est considérée comme l'enzyme la plus abondante de la planète, constituant 30 à 50 % de la protéine soluble totale des chloroplastes végétaux. 1 Cependant, outre son utilisation courante dans les photoautotrophes, le cycle de Calvin est également utilisé par de nombreux chimioautotrophes non photosynthétiques pour fixer le CO 2. De plus, d'autres bactéries et archées utilisent des systèmes alternatifs pour la fixation du CO 2. Bien que la plupart des bactéries utilisant des alternatives du cycle de Calvin soient chimioautotrophes, il a également été démontré que certaines bactéries photoautotrophes au soufre vert utilisent une autre voie de fixation du CO 2.
Exercice\(\PageIndex{3}\)
Décrivez les trois étapes du cycle de Calvin.
Concepts clés et résumé
- Les hétérotrophes dépendent des glucides produits par les autotrophes, dont beaucoup sont photosynthétiques, convertissant l'énergie solaire en énergie chimique.
- Différents organismes photosynthétiques utilisent différents mélanges de pigments photosynthétiques, qui élargissent la gamme des longueurs d'onde de lumière qu'un organisme peut absorber.
- Les photosystèmes (PSI et PSII) contiennent chacun un complexe récupérateur de lumière, composé de plusieurs protéines et de pigments associés qui absorbent l'énergie lumineuse. Les réactions de photosynthèse dépendantes de la lumière convertissent l'énergie solaire en énergie chimique, produisant de l'ATP et du NADPH ou du NADH pour stocker temporairement cette énergie.
- Dans la photosynthèse oxygénée, H 2 O sert de donneur d'électrons pour remplacer l'électron central de la réaction, et de l'oxygène se forme en tant que sous-produit. Dans la photosynthèse anoxygène, d'autres molécules réduites telles que l'H 2 S ou le thiosulfate peuvent être utilisées comme donneurs d'électrons ; l'oxygène n'est donc pas formé en tant que sous-produit.
- La photophosphorylation non cyclique est utilisée dans la photosynthèse oxygénée lorsqu'il est nécessaire de produire à la fois de l'ATP et du NADPH. Si les besoins d'une cellule en ATP l'emportent sur ses besoins en NADPH, elle peut effectuer une photophosphorylation cyclique à la place, ne produisant que de l'ATP.
- Les réactions de photosynthèse indépendantes de la lumière utilisent l'ATP et le NADPH issus des réactions dépendantes de la lumière pour fixer le CO 2 dans les molécules de sucre organique.
Notes
- 1 A. Dhingra et coll. « La traduction améliorée d'un gène Rbc S exprimé par les chloroplastes rétablit les niveaux de petites sous-unités et la photosynthèse dans les plantes antisens nucléaires Rbc S. » Actes de l'Académie nationale des sciences des États-Unis d'Amérique 101 n° 16 (2004) :6315—6320.