
14.1 : Le règne végétal
- Page ID
- 186408

Les plantes constituent un groupe d'organismes vaste et varié. Il existe près de 300 000 espèces de plantes répertoriées. 1 Parmi celles-ci, environ 260 000 sont des plantes qui produisent des graines. Les mousses, les fougères, les conifères et les plantes à fleurs font tous partie du règne végétal. Le règne végétal contient principalement des organismes photosynthétiques ; quelques formes parasitaires ont perdu leur capacité de photosynthèse. Le processus de photosynthèse utilise la chlorophylle, qui se trouve dans des organites appelés chloroplastes. Les plantes possèdent des parois cellulaires contenant de la cellulose. La plupart des plantes se reproduisent sexuellement, mais elles utilisent également diverses méthodes de reproduction asexuée. Les plantes présentent une croissance indéterminée, ce qui signifie qu'elles n'ont pas de forme corporelle définitive, mais qu'elles continuent à augmenter de la masse corporelle jusqu'à leur mort.
Adaptations des plantes à la vie terrestre
À mesure que les organismes s'adaptent à la vie sur terre, ils doivent faire face à plusieurs défis liés à l'environnement terrestre. L'eau a été décrite comme « la matière de la vie ». L'intérieur de la cellule, le milieu dans lequel la plupart des petites molécules se dissolvent et se diffusent, et dans lequel se produisent la majorité des réactions chimiques du métabolisme, est une soupe aqueuse. La dessiccation, ou dessèchement, constitue un danger constant pour un organisme exposé à l'air. Même lorsque les parties d'une plante se trouvent à proximité d'une source d'eau, leurs structures aériennes risquent de s'assécher. L'eau fournit de la flottabilité aux organismes qui vivent dans les habitats aquatiques. Sur terre, les plantes doivent développer un support structurel dans l'air, un milieu qui ne leur donne pas la même portance. De plus, les gamètes mâles doivent atteindre les gamètes femelles en utilisant de nouvelles stratégies, car la natation n'est plus possible. Enfin, les gamètes et les zygotes doivent être protégés contre le dessèchement. Les plantes terrestres qui ont réussi ont développé des stratégies pour relever tous ces défis, bien que toutes les adaptations ne soient pas apparues en même temps. Certaines espèces ne se sont pas éloignées du milieu aquatique, tandis que d'autres ont quitté l'eau pour conquérir les environnements les plus secs de la planète.
Pour pallier ces défis de survie, la vie sur terre offre plusieurs avantages. Tout d'abord, le soleil est abondant. Sur terre, la qualité spectrale de la lumière absorbée par le pigment photosynthétique, la chlorophylle, n'est pas filtrée par l'eau ou par les espèces photosynthétiques concurrentes présentes dans la colonne d'eau située au-dessus. Deuxièmement, le dioxyde de carbone est plus facilement disponible parce que sa concentration est plus élevée dans l'air que dans l'eau. De plus, les plantes terrestres ont évolué avant les animaux terrestres ; par conséquent, jusqu'à ce que les terres arides soient colonisées par les animaux, aucun prédateur ne menaçait le bien-être des plantes. Cette situation a changé lorsque les animaux sont sortis de l'eau et ont découvert d'abondantes sources de nutriments dans la flore établie. À leur tour, les plantes ont développé des stratégies pour empêcher la prédation : des épines et des épines aux produits chimiques toxiques.
Les premières plantes terrestres, comme les premiers animaux terrestres, ne vivaient pas loin d'une source d'eau abondante et ont développé des stratégies de survie pour lutter contre la sécheresse. L'une de ces stratégies est la tolérance à la sécheresse. Les mousses, par exemple, peuvent se dessécher et former un tapis brun et fragile, mais dès que la pluie met de l'eau à disposition, les mousses l'absorbent et reprennent leur aspect vert et sain. Une autre stratégie consiste à coloniser les environnements à forte humidité où les sécheresses sont rares. Les fougères, une des premières lignées de plantes, prospèrent dans les endroits humides et frais, tels que le sous-étage des forêts tempérées. Plus tard, les plantes se sont éloignées des milieux aquatiques en utilisant la résistance à la dessiccation plutôt que la tolérance. Ces plantes, comme les cactus, minimisent les pertes d'eau au point qu'elles peuvent survivre dans les environnements les plus secs de la planète.
Outre les adaptations spécifiques à la vie terrestre, les plantes terrestres présentent des adaptations qui sont responsables de leur diversité et de leur prédominance dans les écosystèmes terrestres. Quatre adaptations majeures sont observées chez de nombreuses plantes terrestres : l'alternance de générations, un sporange dans lequel se forment les spores, un gamétange qui produit des cellules haploïdes et, chez les plantes vasculaires, le tissu méristème apical des racines et des pousses.
Alternance de générations
L'alternance des générations décrit un cycle de vie au cours duquel un organisme possède à la fois des stades multicellulaires haploïdes et diploïdes (Figure\(\PageIndex{1}\)).
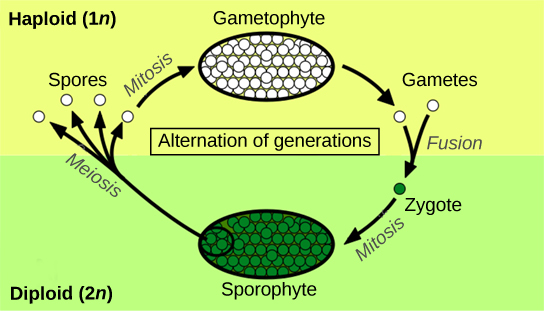
L'haplontique fait référence à un cycle de vie au cours duquel se produit un stade haploïde dominant. La diplontique fait référence à un cycle de vie dans lequel le stade diploïde est le stade dominant et où le nombre de chromosomes haploïdes n'est observé que pendant une brève période du cycle de vie pendant la reproduction sexuée. Les humains sont diplontiques, par exemple. La plupart des plantes présentent une alternance de générations, qualifiée d'haplodiplontique : la forme multicellulaire haploïde connue sous le nom de gamétophyte est suivie dans la séquence de développement par un organisme diploïde multicellulaire, le sporophyte. Le gamétophyte donne naissance aux gamètes, ou cellules reproductrices, par mitose. Il peut s'agir de la phase la plus évidente du cycle de vie de la plante, comme dans les mousses, ou elle peut se produire dans une structure microscopique, comme un grain de pollen chez les plantes supérieures (terme collectif désignant les plantes vasculaires). Le stade sporophyte est à peine perceptible chez les plantes inférieures (terme collectif désignant les groupes de plantes que sont les mousses, les hépatiques et les hornworts). Les arbres imposants constituent la phase diplontique du cycle de vie de plantes telles que les séquoias et les pins.
Les sporanges chez les plantes sans pépins
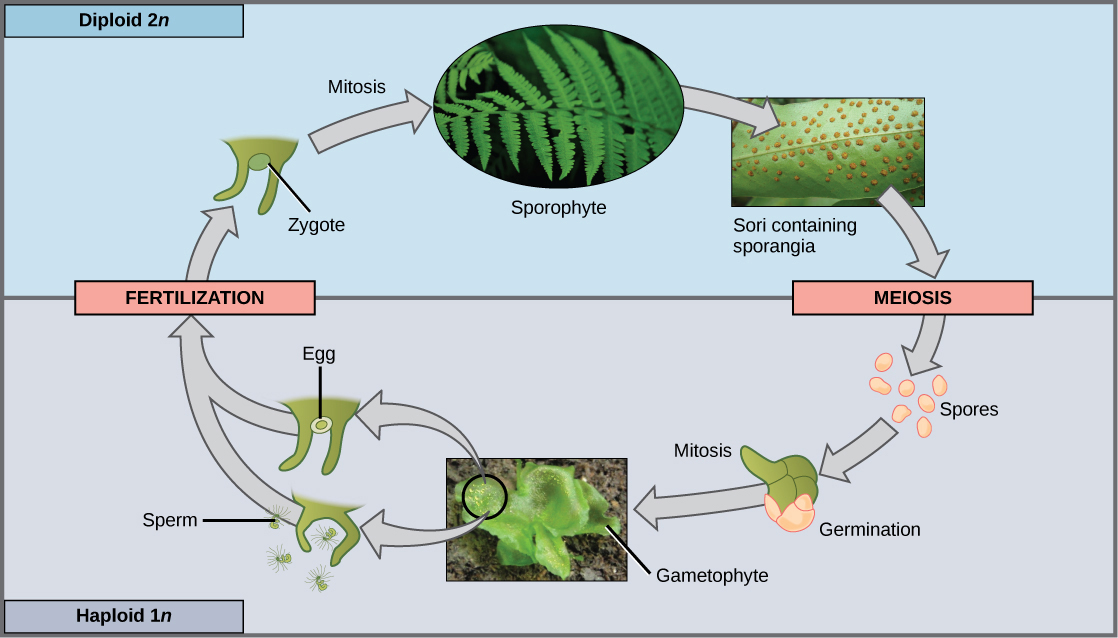
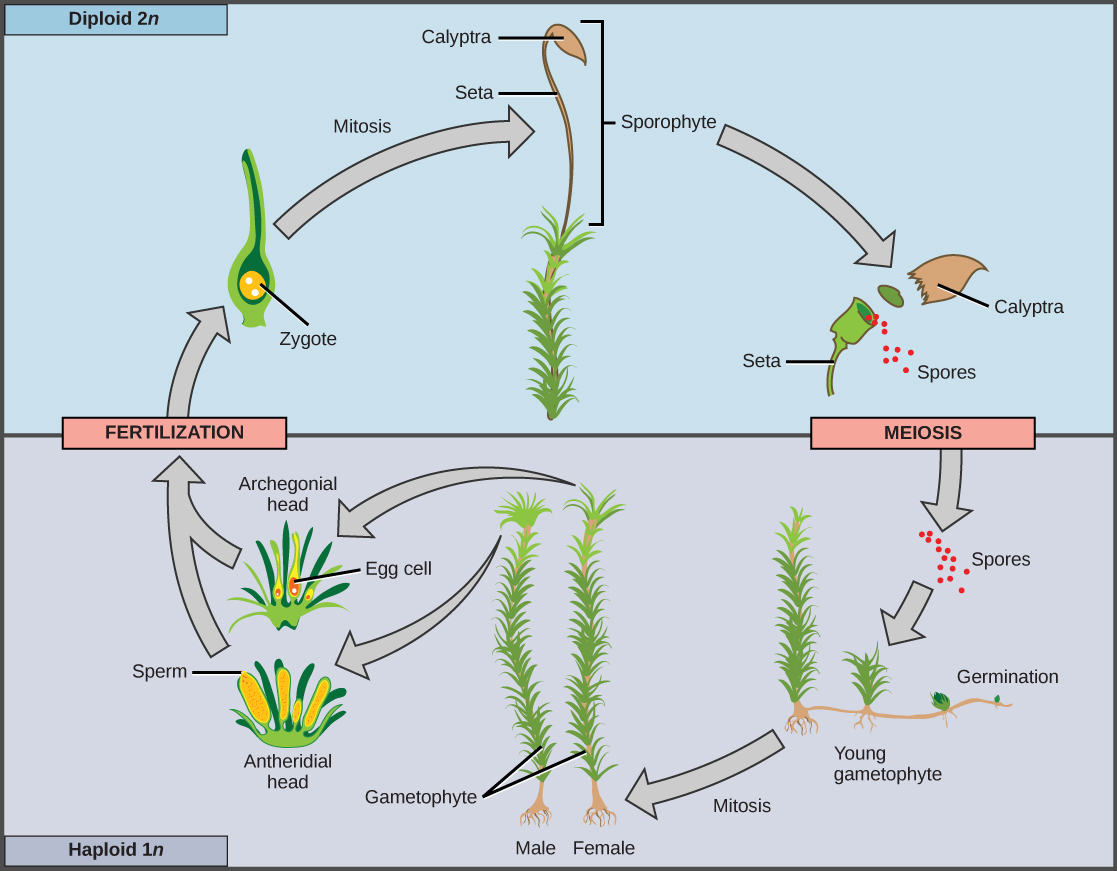
Le sporophyte des plantes sans pépins est diploïde et résulte de la syngamie ou de la fusion de deux gamètes (Figure\(\PageIndex{1}\)). Le sporophyte porte les sporanges (singulier, sporange), organes qui sont apparus pour la première fois chez les plantes terrestres. Le terme « sporange » signifie littéralement « spore dans un vaisseau », car il s'agit d'un sac reproducteur qui contient des spores. À l'intérieur des sporanges multicellulaires, les sporocytes diploïdes, ou cellules mères, produisent des spores haploïdes par méiose, ce qui réduit le nombre de chromosomes 2 n à 1 n. Les spores sont ensuite libérées par les sporanges et se dispersent dans l'environnement. Deux types de spores différents sont produits par les plantes terrestres, ce qui entraîne la séparation des sexes à différents moments du cycle de vie. Les plantes non vasculaires sans pépins (appelées plus précisément « plantes non vasculaires sans pépins avec une phase gamétophyte dominante ») ne produisent qu'un seul type de spores et sont dites homosexuelles. Après avoir germé à partir d'une spore, le gamétophyte produit des gamétangies mâles et femelles, généralement sur le même individu. En revanche, les plantes hétéroporeuses produisent deux types de spores morphologiquement différents. Les spores mâles sont appelées microspores en raison de leur plus petite taille ; les mégaspores, relativement plus grandes, se transformeront en gamétophyte femelle. L'hétérosporie est observée chez quelques plantes vasculaires sans pépins et chez toutes les plantes à graines.
Lorsque la spore haploïde germe, elle génère un gamétophyte multicellulaire par mitose. Le gamétophyte soutient le zygote formé à partir de la fusion des gamètes et de la jeune forme sporophyte ou végétative qui en résulte, et le cycle recommence (Figure\(\PageIndex{2}\) et Figure\(\PageIndex{3}\)).
Les spores des plantes sans pépins et le pollen des plants à graines sont entourés d'épaisses parois cellulaires contenant un polymère résistant appelé sporopollénine. Cette substance se caractérise par de longues chaînes de molécules organiques liées aux acides gras et aux caroténoïdes, et donne à la plupart des pollens leur couleur jaune. La sporopollénine est exceptionnellement résistante à la dégradation chimique et biologique. Sa ténacité explique l'existence de fossiles de pollen bien conservés. La sporopollénine était autrefois considérée comme une innovation des plantes terrestres ; toutefois, l'algue verte Coléochètes est maintenant connue pour former des spores contenant de la sporopollénine.
La protection de l'embryon est une exigence majeure pour les plantes terrestres. L'embryon vulnérable doit être protégé de la dessiccation et d'autres risques environnementaux. Tant chez les plantes sans pépins que chez les plantes à graines, le gamétophyte femelle fournit de la nourriture, et chez les plantes à graines, l'embryon est également protégé au fur et à mesure qu'il se développe pour former la nouvelle génération de sporophytes.
La gamétangie chez les plantes sans pépins
Les gamétanges (au singulier, gamétangium) sont des structures situées sur les gamétophytes des plantes sans pépins dans lesquelles les gamètes sont produits par mitose. Le gamétange mâle, l'anthéridie, libère du sperme. De nombreuses plantes sans pépins produisent des spermatozoïdes munis de flagelles qui leur permettent de nager dans un environnement humide jusqu'à l'archégonie, le gamétange femelle. L'embryon se développe à l'intérieur de l'archégonium sous forme de sporophyte.
Méristems apicaux
Les pousses et les racines des plantes augmentent en longueur grâce à une division cellulaire rapide au sein d'un tissu appelé méristème apical (Figure\(\PageIndex{4}\)). Le méristème apical est un chapeau de cellules à l'extrémité de la pousse ou de la racine constitué de cellules indifférenciées qui continuent de proliférer tout au long de la vie de la plante. Les cellules méristématiques donnent naissance à tous les tissus spécialisés de la plante. L'allongement des pousses et des racines permet à la plante d'accéder à de l'espace et à des ressources supplémentaires : lumière dans le cas de la pousse, eau et minéraux dans le cas des racines. Un méristème distinct, appelé méristème latéral, produit des cellules qui augmentent le diamètre des tiges et des troncs d'arbres. Les méristèmes apicaux sont une adaptation qui permet aux plantes vasculaires de se développer dans des directions essentielles à leur survie : vers le haut pour une plus grande disponibilité de la lumière solaire, et vers le bas dans le sol pour obtenir de l'eau et des minéraux essentiels.

Adaptations supplémentaires des plantes terrestres
Au fur et à mesure que les plantes s'adaptaient à la terre sèche et devenaient indépendantes de la présence constante d'eau dans les habitats humides, de nouveaux organes et structures ont fait leur apparition. Les premières plantes terrestres ne poussaient pas à plus de quelques centimètres du sol et, sur ces tapis bas, elles se disputaient la lumière. En faisant évoluer une pousse et en grandissant, les plantes individuelles captent plus de lumière. Comme l'air offre beaucoup moins de soutien que l'eau, les plantes terrestres ont incorporé des molécules plus rigides dans leurs tiges (et plus tard, dans les troncs d'arbres). L'évolution du tissu vasculaire pour la distribution de l'eau et des solutés était une condition préalable nécessaire pour que les plantes puissent développer des corps plus grands. Le système vasculaire contient des tissus du xylème et du phloème. Le xylème conduit l'eau et les minéraux extraits du sol jusqu'à la pousse ; le phloème transporte les aliments issus de la photosynthèse dans l'ensemble de la plante. Le système racinaire qui a évolué pour absorber l'eau et les minéraux a également ancré la pousse de plus en plus grande dans le sol.
Chez les plantes terrestres, une couverture cireuse et imperméable appelée cuticule recouvre les parties aériennes de la plante : les feuilles et les tiges. La cuticule empêche également l'apport de dioxyde de carbone nécessaire à la synthèse des glucides par photosynthèse. Des stomates, ou pores, qui s'ouvrent et se ferment pour réguler le trafic de gaz et de vapeur d'eau sont donc apparus chez les plantes lorsqu'elles se déplaçaient dans des habitats plus secs.
Les plantes ne peuvent pas éviter les animaux prédateurs. Au lieu de cela, ils synthétisent une large gamme de métabolites secondaires toxiques : des molécules organiques complexes telles que les alcaloïdes, dont les odeurs nocives et le goût désagréable dissuadent les animaux. Ces composés toxiques peuvent provoquer des maladies graves et même la mort.
De plus, au fur et à mesure que les plantes évoluaient avec les animaux, des métabolites sucrés et nutritifs ont été développés pour inciter les animaux à fournir une aide précieuse à la dispersion des grains de pollen, des fruits ou des graines. Les plantes coévoluent avec des animaux depuis des centaines de millions d'années (Figure\(\PageIndex{5}\)).

L'ÉVOLUTION EN ACTION : la paléobotanique
La manière dont les organismes ont acquis les caractéristiques qui leur permettent de coloniser de nouveaux environnements et comment l'écosystème contemporain est façonné sont des questions fondamentales de l'évolution. La paléobotanique répond à ces questions en se spécialisant dans l'étude des plantes éteintes. Les paléobotanistes analysent des spécimens extraits d'études de terrain, reconstituant ainsi la morphologie d'organismes disparus depuis longtemps. Ils retracent l'évolution des plantes en suivant les modifications de la morphologie des plantes et mettent en lumière le lien entre les plantes existantes en identifiant des ancêtres communs qui présentent les mêmes caractéristiques. Ce domaine cherche à trouver des espèces de transition qui comblent les lacunes dans le développement des organismes modernes. Les fossiles se forment lorsque des organismes sont piégés dans des sédiments ou des environnements où leurs formes sont préservées (Figure\(\PageIndex{6}\)). Les paléobotanistes déterminent l'âge géologique des spécimens et la nature de leur environnement en utilisant les sédiments géologiques et les organismes fossiles qui les entourent. L'activité nécessite un soin particulier pour préserver l'intégrité des fossiles délicats et des couches dans lesquelles ils se trouvent.
L'un des développements récents les plus intéressants de la paléobotanique est l'utilisation de la chimie analytique et de la biologie moléculaire pour étudier les fossiles. La préservation des structures moléculaires nécessite un environnement exempt d'oxygène, car l'oxydation et la dégradation de la matière par l'activité des microorganismes dépendent de la présence d'oxygène. Un exemple de l'utilisation de la chimie analytique et de la biologie moléculaire est l'identification de l'oléanane, un composé qui dissuade les organismes nuisibles et qui, jusqu'à présent, semble être propre aux plantes à fleurs. L'oléanane a été récupéré à partir de sédiments datant du Permien, bien avant les dates actuelles données pour l'apparition des premières plantes à fleurs. Les acides nucléiques fossilisés (ADN et ARN) fournissent le plus d'informations. Leurs séquences sont analysées et comparées à celles d'organismes vivants et apparentés. Grâce à cette analyse, des relations évolutives peuvent être établies pour les lignées végétales.
Certains paléobotanistes sont sceptiques quant aux conclusions tirées de l'analyse des fossiles moléculaires. D'une part, les substances chimiques d'intérêt se dégradent rapidement lors de l'isolement initial lorsqu'elles sont exposées à l'air, ainsi que lors de manipulations ultérieures. Il existe toujours un risque élevé de contamination des échantillons par des matières étrangères, provenant principalement de microorganismes. Néanmoins, à mesure que la technologie s'affine, l'analyse de l'ADN des plantes fossilisées fournira des informations précieuses sur l'évolution des plantes et leur adaptation à un environnement en constante évolution.

Les principales divisions des plantes terrestres
Les plantes terrestres sont classées en deux groupes principaux en fonction de l'absence ou de la présence de tissu vasculaire, comme détaillé dans la figure\(\PageIndex{7}\). Les plantes dépourvues de tissu vasculaire formé de cellules spécialisées pour le transport de l'eau et des nutriments sont appelées plantes non vasculaires. Les bryophytes, hépatiques, mousses et hornworts sont dépourvus de pépins et non vasculaires et sont probablement apparus au début de l'évolution des plantes terrestres. Les plantes vasculaires ont développé un réseau de cellules qui transportent l'eau et les solutés à travers le corps végétal. Les premières plantes vasculaires sont apparues à la fin de l'Ordovicien (il y a 461 à 444 millions d'années) et étaient probablement similaires aux lycophytes, qui comprennent les massues (à ne pas confondre avec les mousses) et les ptérophytes (fougères, prêles et fougères fouettées). Les lycophytes et les ptérophytes sont appelés plantes vasculaires sans pépins. Ils ne produisent pas de graines, qui sont des embryons dont les réserves alimentaires stockées sont protégées par une enveloppe rigide. Les plantes à graines constituent le plus grand groupe de toutes les plantes existantes et dominent donc le paysage. Les plantes à graines comprennent les gymnospermes, notamment les conifères, qui produisent des « graines nues », et les plantes les plus performantes, les plantes à fleurs ou angiospermes, qui protègent leurs graines dans des chambres situées au centre de la fleur. Les parois de ces chambres se transforment ensuite en fruits.

Résumé de la section
Les plantes terrestres ont développé des caractéristiques qui ont permis de coloniser les terres et de survivre hors de l'eau. Les adaptations à la vie terrestre comprennent les tissus vasculaires, les racines, les feuilles, les cuticules cireuses et une couche externe résistante qui protège les spores. Les plantes terrestres comprennent les plantes non vasculaires et les plantes vasculaires. Les plantes vasculaires, qui comprennent les plantes sans pépins et les plantes à graines, possèdent des méristèmes apicaux et des embryons dotés de réserves nutritives. Toutes les plantes terrestres partagent les caractéristiques suivantes : alternance de générations, la plante haploïde étant appelée gamétophyte et la plante diploïde appelée sporophyte ; formation de spores haploïdes dans un sporange ; formation de gamètes dans un gamétange.
Notes
- 1 A.D. Chapman (2009) Nombre d'espèces vivantes en Australie et dans le monde. 2e édition. Un rapport pour l'étude des ressources biologiques australiennes. Services australiens d'information sur la biodiversité, Toowoomba, Australie. Disponible en ligne à l'adresse http://www.environment.gov.au/biodiv...ps-plants.html.
Lexique
- méristème apical
- le point de croissance d'une plante vasculaire situé à l'extrémité d'une pousse ou d'une racine où se produit la division cellulaire
- diplontique
- décrit un cycle de vie dans lequel le stade diploïde est le stade dominant
- gamétangium
- (pluriel : gamétange) structure au sein de laquelle les gamètes sont produits
- gamétophyte
- la plante haploïde qui produit des gamètes
- haplodiplontique
- décrit un cycle de vie dans lequel les stades haploïde et diploïde alternent ; également connu sous le nom d'alternance du cycle de vie des générations
- haplontique
- décrit un cycle de vie dans lequel le stade haploïde est le stade dominant
- hétéroporeux
- ayant deux types de spores qui donnent naissance à des gamétophytes mâles et femelles
- homosporeux
- ayant un type de spore qui donne naissance à des gamétophytes qui donnent naissance à des gamètes mâles et femelles
- plante non vasculaire
- plante dépourvue de tissu vasculaire formé de cellules spécialisées pour le transport de l'eau et des nutriments
- sporange
- (pluriel : sporanges) organe dans lequel les spores sont produites
- sporophyte
- la plante diploïde qui produit des spores
- syngamie
- l'union de deux gamètes lors de la fécondation
- plante vasculaire
- une plante dans laquelle se trouve un réseau de cellules qui conduisent l'eau et les solutés à travers l'organisme