
9.5 : Comment les gènes sont-ils régulés
- Page ID
- 186316

Pour qu'une cellule fonctionne correctement, les protéines nécessaires doivent être synthétisées au bon moment. Tous les organismes et cellules contrôlent ou régulent la transcription et la traduction de leur ADN en protéines. Le processus qui consiste à activer un gène pour produire de l'ARN et des protéines est appelé expression génique. Que ce soit dans un organisme unicellulaire simple ou dans un organisme multicellulaire complexe, chaque cellule contrôle quand et comment ses gènes sont exprimés. Pour que cela se produise, il doit y avoir un mécanisme permettant de contrôler le moment où un gène est exprimé pour produire de l'ARN et des protéines, la quantité de protéine produite et le moment où il est temps d'arrêter de fabriquer cette protéine parce qu'elle n'est plus nécessaire.
Les cellules des organismes multicellulaires sont spécialisées ; les cellules de différents tissus ont une apparence très différente et remplissent des fonctions différentes. Par exemple, une cellule musculaire est très différente d'une cellule du foie, qui est très différente d'une cellule de la peau. Ces différences sont la conséquence de l'expression de différents ensembles de gènes dans chacune de ces cellules. Toutes les cellules ont certaines fonctions de base qu'elles doivent remplir elles-mêmes, telles que la conversion de l'énergie des molécules de sucre en énergie de l'ATP. Chaque cellule possède également de nombreux gènes qui ne sont pas exprimés et en exprime de nombreux qui ne sont pas exprimés par d'autres cellules, de sorte qu'elle peut exercer ses fonctions spécialisées. De plus, les cellules activeront ou désactiveront certains gènes à différents moments en réponse à des changements dans l'environnement ou à différents moments du développement de l'organisme. Les organismes unicellulaires, qu'ils soient eucaryotes ou procaryotes, activent et désactivent également les gènes en réponse aux exigences de leur environnement afin de pouvoir répondre à des conditions particulières.
Le contrôle de l'expression des gènes est extrêmement complexe. Les dysfonctionnements de ce processus sont préjudiciables à la cellule et peuvent entraîner le développement de nombreuses maladies, dont le cancer.
Expression des gènes procaryotes ou eucaryotes
Pour comprendre comment l'expression des gènes est régulée, il faut d'abord comprendre comment un gène devient une protéine fonctionnelle dans une cellule. Le processus se produit à la fois dans les cellules procaryotes et eucaryotes, mais de manière légèrement différente.
Comme les organismes procaryotes n'ont pas de noyau cellulaire, les processus de transcription et de traduction se produisent presque simultanément. Lorsque la protéine n'est plus nécessaire, la transcription s'arrête. Par conséquent, la principale méthode pour contrôler le type et la quantité de protéines exprimées dans une cellule procaryote consiste à réguler la transcription de l'ADN en ARN. Toutes les étapes suivantes se déroulent automatiquement. Lorsque plus de protéines sont nécessaires, plus de transcription se produit. Par conséquent, dans les cellules procaryotes, le contrôle de l'expression des gènes se fait presque entièrement au niveau de la transcription.
Le premier exemple d'un tel contrôle a été découvert en utilisant E. coli dans les années 1950 et 1960 par des chercheurs français et est appelé opéron lac. L'opéron lac est un fragment d'ADN comportant trois gènes adjacents qui codent pour des protéines qui participent à l'absorption et au métabolisme du lactose, une source alimentaire de E. coli. Lorsque le lactose n'est pas présent dans l'environnement de la bactérie, les gènes lac sont transcrits en petites quantités. Lorsque du lactose est présent, les gènes sont transcrits et la bactérie est capable d'utiliser le lactose comme source alimentaire. L'opéron contient également une séquence promotrice à laquelle l'ARN polymérase se lie pour commencer la transcription ; entre le promoteur et les trois gènes se trouve une région appelée opérateur. En l'absence de lactose, une protéine appelée répresseur se lie à l'opérateur et empêche l'ARN polymérase de se lier au promoteur, sauf dans de rares cas. Ainsi, très peu de produits protéiques des trois gènes sont produits. Lorsque du lactose est présent, un produit final du métabolisme du lactose se lie à la protéine répressive et empêche celle-ci de se lier à l'opérateur. Cela permet à l'ARN polymérase de se lier au promoteur et de transcrire librement les trois gènes, permettant ainsi à l'organisme de métaboliser le lactose.
Les cellules eucaryotes, en revanche, possèdent des organites intracellulaires et sont beaucoup plus complexes. Rappelons que dans les cellules eucaryotes, l'ADN est contenu à l'intérieur du noyau de la cellule et y est transcrit en ARNm. L'ARNm nouvellement synthétisé est ensuite transporté hors du noyau vers le cytoplasme, où les ribosomes traduisent l'ARNm en protéine. Les processus de transcription et de traduction sont physiquement séparés par la membrane nucléaire ; la transcription se produit uniquement à l'intérieur du noyau, et la traduction n'a lieu qu'à l'extérieur du noyau dans le cytoplasme. La régulation de l'expression génique peut intervenir à tous les stades du processus (Figure\(\PageIndex{1}\)). La régulation peut se produire lorsque l'ADN est déroulé et détaché des nucléosomes pour se lier aux facteurs de transcription (niveau épigénétique), lorsque l'ARN est transcrit (niveau transcriptionnel), lorsque l'ARN est traité et exporté vers le cytoplasme après sa transcription (niveau post-transcriptionnel), lorsque l'ARN est traduit en protéine (niveau translationnel), ou après la fabrication de la protéine (niveau post-traductionnel).
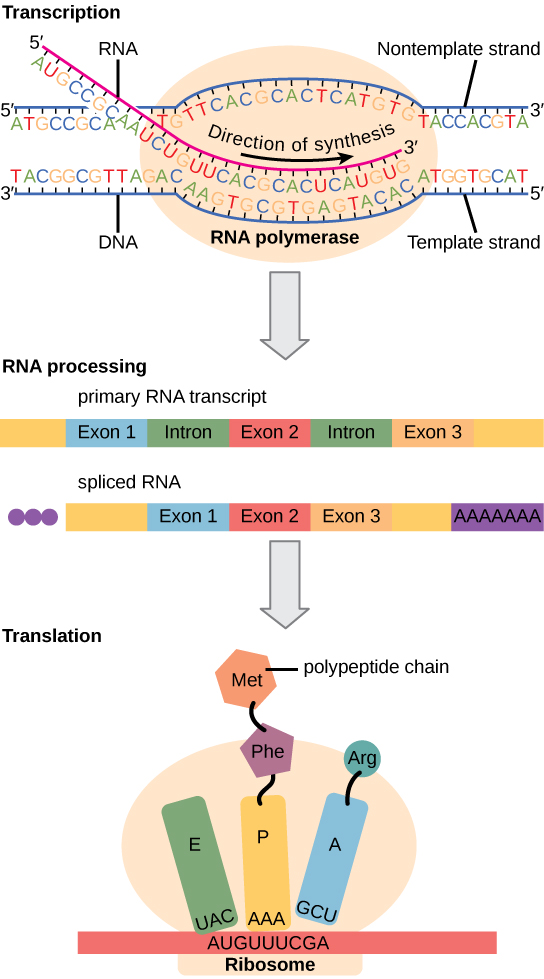
Les différences de régulation de l'expression génique entre les procaryotes et les eucaryotes sont résumées dans le tableau\(\PageIndex{1}\).
| Organismes procaryotes | Organismes eucaryotes |
|---|---|
| Noyau manquant | Contient un noyau |
| La transcription de l'ARN et la traduction des protéines se produisent presque |
|
| L'expression des gènes est régulée principalement au niveau transcriptionnel | L'expression des gènes est régulée à de nombreux niveaux (épigénétique, transcriptionnel, post-transcriptionnel, translationnel et post-traductionnel) |
L'ÉVOLUTION EN ACTION : épissage alternatif de l'ARN
Dans les années 1970, des gènes présentant un épissage alternatif de l'ARN ont été observés pour la première fois. L'épissage alternatif de l'ARN est un mécanisme qui permet de produire différents produits protéiques à partir d'un gène lorsque différentes combinaisons d'introns (et parfois d'exons) sont retirées du transcrit (Figure\(\PageIndex{2}\)). Cet épissage alternatif peut être aléatoire, mais le plus souvent, il est contrôlé et agit comme un mécanisme de régulation génique, la fréquence des différentes alternatives d'épissage étant contrôlée par la cellule afin de contrôler la production de différents produits protéiques dans différentes cellules, ou à différents stades de développement. L'épissage alternatif est maintenant considéré comme un mécanisme courant de régulation génique chez les eucaryotes ; selon une estimation, 70 % des gènes chez l'homme sont exprimés sous forme de protéines multiples par épissage alternatif.
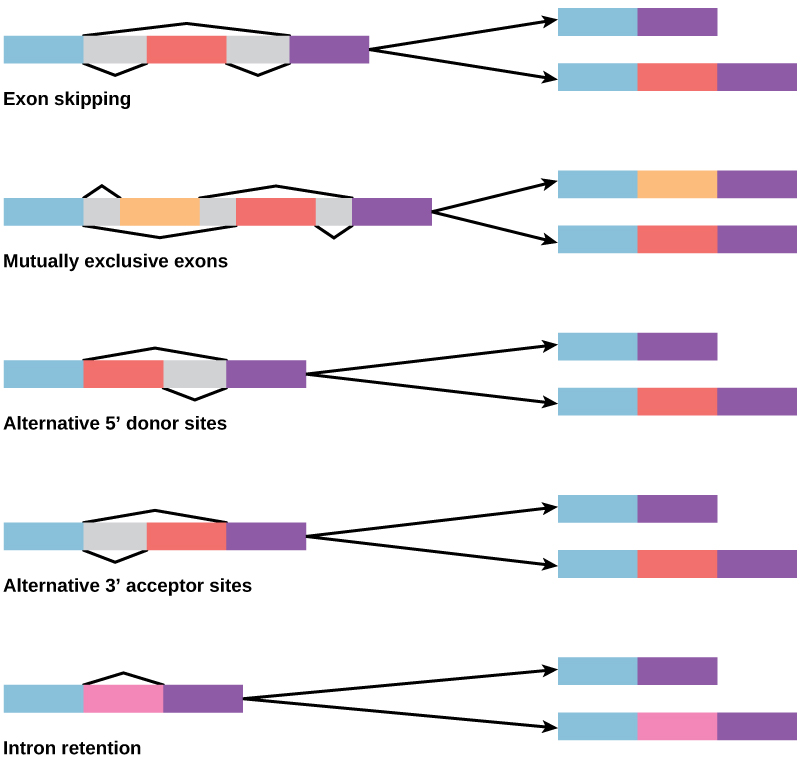
Comment l'épissage alternatif pourrait-il évoluer ? Les introns ont une séquence de reconnaissance de début et de fin, et il est facile d'imaginer l'échec du mécanisme d'épissage à identifier l'extrémité d'un intron et à trouver la fin de l'intron suivant, éliminant ainsi deux introns et l'exon intermédiaire. En fait, des mécanismes sont en place pour empêcher de tels sauts d'exons, mais des mutations sont susceptibles d'entraîner leur échec. De telles « erreurs » produiraient probablement une protéine non fonctionnelle. En effet, la cause de nombreuses maladies génétiques est l'épissage alternatif plutôt que les mutations d'une séquence. Cependant, un épissage alternatif créerait un variant protéique sans perte de la protéine d'origine, ouvrant ainsi des possibilités d'adaptation du nouveau variant à de nouvelles fonctions. La duplication de gènes a joué un rôle important dans l'évolution de nouvelles fonctions de la même manière, en fournissant des gènes qui peuvent évoluer sans éliminer la protéine fonctionnelle d'origine.
Résumé
Bien que toutes les cellules somatiques d'un organisme contiennent le même ADN, toutes les cellules de cet organisme n'expriment pas les mêmes protéines. Les organismes procaryotes expriment la totalité de l'ADN qu'ils codent dans chaque cellule, mais pas nécessairement tous en même temps. Les protéines ne sont exprimées que lorsqu'elles sont nécessaires. Les organismes eucaryotes expriment un sous-ensemble de l'ADN codé dans une cellule donnée. Dans chaque type de cellule, le type et la quantité de protéines sont régulés en contrôlant l'expression génique. Pour exprimer une protéine, l'ADN est d'abord transcrit en ARN, qui est ensuite traduit en protéines. Dans les cellules procaryotes, ces processus se produisent presque simultanément. Dans les cellules eucaryotes, la transcription se produit dans le noyau et est distincte de la traduction qui se produit dans le cytoplasme. L'expression génique chez les procaryotes est régulée uniquement au niveau transcriptionnel, tandis que dans les cellules eucaryotes, l'expression génique est régulée aux niveaux épigénétique, transcriptionnel, post-transcriptionnel, translationnel et post-traductionnel.
Lexique
- épissage alternatif de l'ARN
- un mécanisme de régulation génique post-transcriptionnel chez les eucaryotes dans lequel de multiples produits protéiques sont produits par un seul gène au moyen de combinaisons alternatives d'épissage du transcrit d'ARN
- épigénétique
- décrire les facteurs de régulation non génétiques, tels que les modifications apportées aux protéines histones et à l'ADN qui contrôlent l'accessibilité aux gènes des chromosomes
- expression des gènes
- processus qui contrôlent si un gène est exprimé
- post-transcriptionnel
- contrôle de l'expression génique après la création de la molécule d'ARN mais avant qu'elle ne soit traduite en protéine
- post-traductionnel
- contrôle de l'expression génique après la création d'une protéine