
9.3 : Transcription
- Page ID
- 186315

Chez les procaryotes comme chez les eucaryotes, la deuxième fonction de l'ADN (la première était la réplication) est de fournir les informations nécessaires à la construction des protéines nécessaires pour que la cellule puisse remplir toutes ses fonctions. Pour ce faire, l'ADN est « lu » ou transcrit dans une molécule d'ARNm. L'ARNm fournit ensuite le code permettant de former une protéine par un processus appelé traduction. Grâce aux processus de transcription et de traduction, une protéine est construite avec une séquence spécifique d'acides aminés initialement codée dans l'ADN. Ce module aborde les détails de la transcription.
Le dogme central : l'ADN code pour l'ARN ; l'ARN code pour les protéines
Le flux d'informations génétiques dans les cellules, de l'ADN à l'ARNm en passant par les protéines, est décrit par le dogme central (Figure\(\PageIndex{1}\)), selon lequel les gènes spécifient les séquences des ARNm, qui à leur tour spécifient les séquences des protéines.
La copie de l'ADN en ARNm est relativement simple, un nucléotide étant ajouté au brin d'ARNm pour chaque nucléotide complémentaire lu dans le brin d'ADN. La traduction en protéine est plus complexe car des groupes de trois nucléotides d'ARNm correspondent à un acide aminé de la séquence protéique. Cependant, comme nous le verrons dans le module suivant, la traduction en protéine est toujours systématique, de sorte que les nucléotides 1 à 3 correspondent à l'acide aminé 1, les nucléotides 4 à 6 correspondent à l'acide aminé 2, etc.
Transcription : de l'ADN à l'ARNm
Les procaryotes et les eucaryotes effectuent fondamentalement le même processus de transcription, avec la différence importante du noyau lié à la membrane chez les eucaryotes. Lorsque les gènes sont liés au noyau, la transcription a lieu dans le noyau de la cellule et le transcrit de l'ARNm doit être transporté vers le cytoplasme. Les procaryotes, qui comprennent les bactéries et les archées, sont dépourvus de noyaux liés à la membrane et d'autres organites, et la transcription se produit dans le cytoplasme de la cellule. Chez les procaryotes comme chez les eucaryotes, la transcription se déroule en trois étapes principales : initiation, élongation et terminaison.
Initiation
La transcription nécessite que la double hélice de l'ADN se déroule partiellement dans la région de synthèse de l'ARNm. La zone de déroulement est appelée bulle de transcription. La séquence d'ADN sur laquelle les protéines et les enzymes impliquées dans la transcription se lient pour initier le processus est appelée promoteur. Dans la plupart des cas, les promoteurs existent en amont des gènes qu'ils régulent. La séquence spécifique d'un promoteur est très importante car elle détermine si le gène correspondant est transcrit tout le temps, de temps en temps ou presque (Figure\(\PageIndex{2}\)).
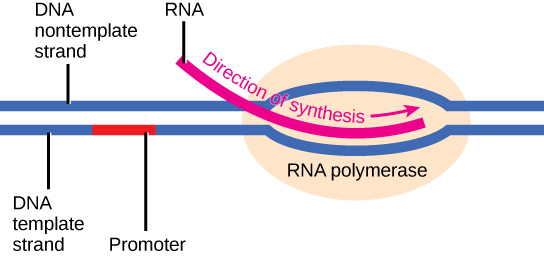
Allongement
La transcription se fait toujours à partir de l'un des deux brins d'ADN, appelé brin matrice. Le produit de l'ARNm est complémentaire du brin matrice et est presque identique à l'autre brin d'ADN, appelé brin non matrice, à l'exception du fait que l'ARN contient un uracile (U) à la place de la thymine (T) présente dans l'ADN. Au cours de l'élongation, une enzyme appelée ARN polymérase se déplace le long de la matrice d'ADN en ajoutant des nucléotides par appariement de bases avec la matrice d'ADN d'une manière similaire à la réplication de l'ADN, à la différence près qu'un brin d'ARN est synthétisé qui ne reste pas lié à la matrice d'ADN. Au fur et à mesure que l'élongation se poursuit, l'ADN est continuellement déroulé devant l'enzyme centrale et enroulé à nouveau derrière elle (Figure\(\PageIndex{3}\)).
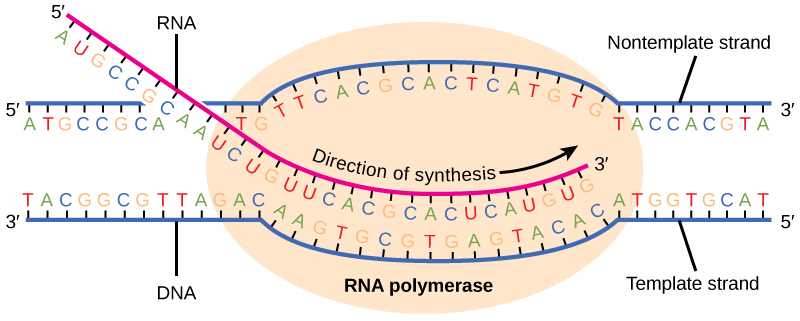
Résiliation
Une fois qu'un gène est transcrit, il faut demander à la polymérase procaryote de se dissocier de la matrice d'ADN et de libérer l'ARNm nouvellement créé. Selon le gène transcrit, il existe deux types de signaux de terminaison, mais les deux impliquent des séquences nucléotidiques répétées dans la matrice d'ADN qui entraînent le blocage de l'ARN polymérase, la sortie de la matrice d'ADN et la libération du transcrit de l'ARNm.
À la fin, le processus de transcription est terminé. Dans une cellule procaryote, au moment de l'arrêt, le transcrit aurait déjà été utilisé pour synthétiser partiellement de nombreuses copies de la protéine codée, car ces processus peuvent se produire simultanément à l'aide de plusieurs ribosomes (polyribosomes) (Figure\(\PageIndex{4}\)). En revanche, la présence d'un noyau dans les cellules eucaryotes empêche la transcription et la traduction simultanées.
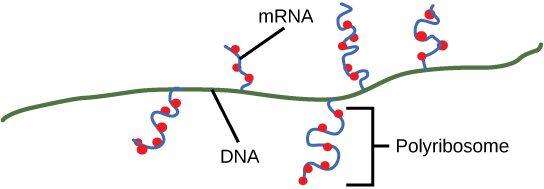
Traitement de l'ARN eucaryote
Les ARNm eucaryotes nouvellement transcrits doivent subir plusieurs étapes de traitement avant de pouvoir être transférés du noyau au cytoplasme et traduits en protéine. Les étapes supplémentaires impliquées dans la maturation de l'ARNm eucaryote créent une molécule beaucoup plus stable qu'un ARNm procaryote. Par exemple, les ARNm eucaryotes durent plusieurs heures, alors que l'ARNm procaryote typique ne dure pas plus de cinq secondes.
Le transcrit de l'ARNm est d'abord enrobé de protéines stabilisant l'ARN pour empêcher sa dégradation lors de sa transformation et de son exportation hors du noyau. Cela se produit alors que le pré-ARNm est encore en cours de synthèse en ajoutant un « capuchon » nucléotidique spécial à l'extrémité 5' du transcrit en croissance. En plus de prévenir la dégradation, les facteurs impliqués dans la synthèse des protéines reconnaissent le bouchon pour aider à initier la traduction par les ribosomes.
Une fois l'élongation terminée, une enzyme ajoute une chaîne d'environ 200 résidus d'adénine à l'extrémité 3', appelée queue poly-A. Cette modification protège davantage le pré-ARNm de la dégradation et indique aux facteurs cellulaires que le transcrit doit être exporté vers le cytoplasme.
Les gènes eucaryotes sont composés de séquences codant pour des protéines appelées exons (ex- on signifie qu'ils sont exprimés) et de séquences inertes appelées introns (int- ron indique leur rôle int ervenant). Les introns sont retirés du pré-ARNm pendant le traitement. Les séquences d'intron présentes dans l'ARNm ne codent pas pour les protéines fonctionnelles. Il est essentiel que tous les introns d'un pré-ARNm soient complètement et précisément éliminés avant la synthèse des protéines afin que les exons se rejoignent pour coder les bons acides aminés. Si le processus se trompe ne serait-ce que par un seul nucléotide, la séquence des exons réunis serait modifiée et la protéine résultante ne serait pas fonctionnelle. Le processus d'élimination des introns et de reconnexion des exons est appelé épissage (Figure\(\PageIndex{5}\)). Les introns sont retirés et dégradés alors que le pré-ARNm est encore dans le noyau.
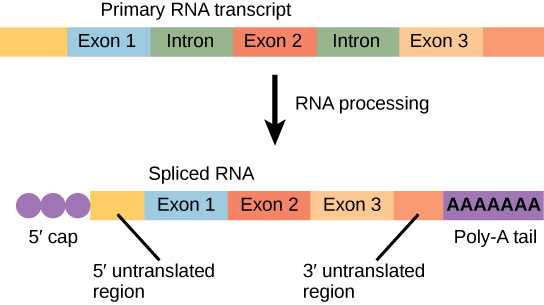
Résumé
Chez les procaryotes, la synthèse de l'ARNm est initiée au niveau d'une séquence promotrice sur la matrice d'ADN. L'élongation synthétise un nouvel ARNm. La terminaison libère l'ARNm et se produit par des mécanismes qui bloquent l'ARN polymérase et la font tomber de la matrice d'ADN. Les ARNm eucaryotes nouvellement transcrits sont modifiés par une calotte et une queue en poly-A. Ces structures protègent l'ARNm mature de la dégradation et aident à l'exporter du noyau. Les ARNm des eucaryotes subissent également un épissage, au cours duquel les introns sont retirés et les exons sont reconnectés avec une précision mononucléotidique. Seuls les ARNm finis sont exportés du noyau vers le cytoplasme.
Lexique
- exon
- une séquence présente dans l'ARNm codant pour les protéines après la fin de l'épissage avant l'ARNm
- intron
- séquences intermédiaires ne codant pas pour des protéines qui sont épissées à partir de l'ARNm pendant le traitement
- ARNm
- ARN messager ; forme d'ARN qui porte le code de séquence nucléotidique d'une séquence protéique traduite en une séquence polypeptidique
- brin non modèle
- le brin d'ADN qui n'est pas utilisé pour transcrire l'ARNm ; ce brin est identique à l'ARNm sauf que les nucléotides T de l'ADN sont remplacés par des nucléotides U dans l'ARNm
- promoteur
- une séquence de l'ADN à laquelle l'ARN polymérase et les facteurs associés se lient et initient la transcription
- ARN polymérase
- une enzyme qui synthétise un brin d'ARN à partir d'un brin matrice d'ADN
- épissage
- le processus d'élimination des introns et de reconnexion des exons dans un pré-ARNm
- brin de modèle
- le brin d'ADN qui spécifie la molécule d'ARNm complémentaire
- bulle de transcription
- la région de l'ADN déroulé localement qui permet la transcription de l'ARNm