
32.2 : Pollinisation et fertilisation
- Page ID
- 189396

Compétences à développer
- Décrire ce qui doit se produire pour la fertilisation des plantes
- Expliquer la pollinisation croisée et la manière dont elle se déroule
- Décrire le processus qui mène au développement d'une graine
- Définir la double fertilisation
Chez les angiospermes, la pollinisation est définie comme le placement ou le transfert du pollen de l'anthère vers le stigmate de la même fleur ou d'une autre fleur. Chez les gymnospermes, la pollinisation implique le transfert du pollen du cône mâle vers le cône femelle. Lors du transfert, le pollen germe pour former le tube pollinique et le sperme nécessaires à la fécondation de l'ovule. La pollinisation est bien étudiée depuis l'époque de Gregor Mendel. Mendel a réussi à pratiquer l'autofécondation et la pollinisation croisée des pois potagers tout en étudiant la transmission des caractéristiques d'une génération à l'autre. Les cultures d'aujourd'hui sont le résultat de la sélection végétale, qui utilise la sélection artificielle pour produire les cultivars actuels. C'est le cas du maïs d'aujourd'hui, qui est le résultat d'années de sélection qui ont débuté avec son ancêtre, la téosinte. La téosinte que les anciens Mayas ont commencé à cultiver contenait de minuscules graines, très différentes des épis de maïs relativement géants d'aujourd'hui. Fait intéressant, bien que ces deux plantes semblent totalement différentes, la différence génétique entre elles est minime.
La pollinisation prend deux formes : l'autopollinisation et la pollinisation croisée. L'autopollinisation se produit lorsque le pollen de l'anthère se dépose sur le stigmate de la même fleur ou d'une autre fleur de la même plante. La pollinisation croisée est le transfert du pollen de l'anthère d'une fleur au stigmate d'une autre fleur sur un individu différent de la même espèce. L'autopollinisation se produit dans les fleurs où l'étamine et le carpelle arrivent à maturité en même temps et sont positionnées de telle sorte que le pollen puisse se déposer sur le stigmate de la fleur. Cette méthode de pollinisation ne nécessite aucun investissement de la part de la plante pour fournir du nectar et du pollen comme nourriture aux pollinisateurs.
Lien vers l'apprentissage
Explorez ce site Web interactif pour examiner l'autopollinisation et la pollinisation croisée : http://passel.unl.edu/pages/animatio...erbreeding.swf
Les espèces vivantes sont conçues pour assurer la survie de leur progéniture ; celles qui échouent disparaissent. La diversité génétique est donc nécessaire pour que certains membres de la descendance puissent survivre dans des conditions environnementales ou de stress changeantes. L'autopollinisation conduit à la production de plantes présentant une moindre diversité génétique, car le matériel génétique de la même plante est utilisé pour former des gamètes et, finalement, le zygote. En revanche, la pollinisation croisée (ou croisement) entraîne une plus grande diversité génétique, car le microgamétophyte et le mégagamétophyte proviennent de plantes différentes.
Parce que la pollinisation croisée permet une plus grande diversité génétique, les plantes ont développé de nombreuses méthodes pour éviter l'autopollinisation. Chez certaines espèces, le pollen et l'ovaire arrivent à maturité à des moments différents. Ces fleurs rendent l'autopollinisation presque impossible. Lorsque le pollen arrive à maturité et est excrété, le stigmate de cette fleur est arrivé à maturité et ne peut être pollinisé que par le pollen d'une autre fleur. Certaines fleurs ont développé des caractéristiques physiques qui empêchent l'autopollinisation. La primevère est l'une de ces fleurs. Les primevères ont développé deux types de fleurs avec des différences de longueur des anthères et des stigmates : la fleur à yeux d'épingle possède des anthères positionnées à mi-chemin du tube pollinique, et le stigmate de la fleur aux yeux de thrum est également situé à mi-chemin. Les insectes se reproduisent facilement lorsqu'ils recherchent le nectar au fond du tube pollinique. Ce phénomène est également connu sous le nom d'hétérostylie. De nombreuses plantes, comme le concombre, ont des fleurs mâles et femelles situées sur différentes parties de la plante, ce qui rend l'autopollinisation difficile. Chez d'autres espèces encore, les fleurs mâles et femelles proviennent de plantes différentes (dioïques). Tous ces éléments constituent des obstacles à l'autopollinisation ; par conséquent, les plantes dépendent des pollinisateurs pour transférer le pollen. La majorité des pollinisateurs sont des agents biotiques tels que les insectes (comme les abeilles, les mouches et les papillons), les chauves-souris, les oiseaux et d'autres animaux. D'autres espèces végétales sont pollinisées par des agents abiotiques tels que le vent et l'eau.
Connexion quotidienne : gènes d'incompatibilité chez les fleurs
Au cours des dernières décennies, des gènes d'incompatibilité, qui empêchent le pollen de germer ou de se développer dans le stigmate d'une fleur, ont été découverts chez de nombreuses espèces d'angiospermes. Si les plantes ne possèdent pas de gènes compatibles, le tube pollinique cesse de croître. L'auto-incompatibilité est contrôlée par le locus S (stérilité). Les tubes polliniques doivent traverser le tissu du stigmate et le coiffer avant de pouvoir pénétrer dans l'ovule. Le carpelle est sélectif quant au type de pollen qu'il laisse pousser à l'intérieur. L'interaction se produit principalement entre le pollen et les cellules épidermiques du stigmate. Chez certaines plantes, comme le chou, le pollen est rejeté à la surface du stigmate et le pollen indésirable ne germe pas. Chez d'autres plantes, la germination du tube pollinique est arrêtée après avoir atteint un tiers de la longueur du style, ce qui entraîne la mort du tube pollinique. La mort du tube pollinique est due soit à l'apoptose (mort cellulaire programmée), soit à la dégradation de l'ARN du tube pollinique. La dégradation résulte de l'activité d'une ribonucléase codée par le locus S. La ribonucléase est sécrétée par les cellules du style dans la matrice extracellulaire, qui se trouve le long du tube pollinique en croissance.
En résumé, l'auto-incompatibilité est un mécanisme qui empêche l'autofécondation chez de nombreuses espèces de plantes à fleurs. Le fonctionnement de ce mécanisme d'auto-incompatibilité a des conséquences importantes pour les sélectionneurs de plantes car il inhibe la production de plantes consanguines et hybrides.
Pollinisation par les insectes
Les abeilles sont peut-être les pollinisateurs les plus importants de nombreuses plantes de jardin et de la plupart des arbres fruitiers commerciaux (Figure\(\PageIndex{1}\)). Les espèces d'abeilles les plus communes sont les bourdons et les abeilles domestiques. Comme les abeilles ne peuvent pas voir la couleur rouge, les fleurs pollinisées par les abeilles ont généralement des nuances de bleu, de jaune ou d'autres couleurs. Les abeilles récoltent du pollen ou du nectar riches en énergie pour leur survie et leurs besoins énergétiques. Ils visitent les fleurs qui sont ouvertes pendant la journée, qui sont de couleurs vives, qui ont un arôme ou une odeur prononcé et qui ont une forme tubulaire, généralement en présence d'un guide à nectar. Un guide sur les nectars comprend des régions des pétales de fleurs qui ne sont visibles que par les abeilles, et non par les humains ; il aide à guider les abeilles vers le centre de la fleur, rendant ainsi le processus de pollinisation plus efficace. Le pollen adhère aux poils duveteux des abeilles, et lorsque l'abeille visite une autre fleur, une partie du pollen est transférée vers la deuxième fleur. Récemment, de nombreux rapports ont fait état du déclin de la population d'abeilles domestiques. De nombreuses fleurs resteront non pollinisées et ne porteront pas de graines si les abeilles domestiques disparaissent. L'impact sur les producteurs de fruits commerciaux pourrait être dévastateur.

De nombreuses mouches sont attirées par les fleurs qui dégagent une odeur pourrie ou une odeur de chair pourrie. Ces fleurs, qui produisent du nectar, ont généralement des couleurs ternes, comme le brun ou le violet. On les trouve sur la fleur cadavre ou le lis vaudou (Amorphophallus), l'arum du dragon (Dracunculus) et la fleur de charogne (Stapleia, Rafflesia). Le nectar fournit de l'énergie, tandis que le pollen fournit des protéines. Les guêpes sont également d'importants insectes pollinisateurs et pollinisent de nombreuses espèces de figues.
Les papillons, tels que le monarque, pollinisent de nombreuses fleurs de jardin et fleurs sauvages, qui se présentent généralement en grappes. Ces fleurs sont de couleurs vives, ont un parfum fort, sont ouvertes pendant la journée et sont dotées de guides sur le nectar pour faciliter l'accès au nectar. Le pollen est ramassé et transporté sur les membres du papillon. Les papillons, quant à eux, pollinisent les fleurs en fin d'après-midi et pendant la nuit. Les fleurs pollinisées par les papillons sont pâles ou blanches et sont plates, ce qui permet aux papillons de se poser. Un exemple bien étudié de plante pollinisée par les papillons est le yucca, qui est pollinisé par la teigne du yucca. La forme de la fleur et du papillon s'est adaptée de manière à permettre une pollinisation réussie. Le papillon dépose du pollen sur le stigmate collant pour que la fécondation ait lieu plus tard. La femelle dépose également des œufs dans l'ovaire. Au fur et à mesure que les œufs se transforment en larves, ils se nourrissent de la fleur et des graines en développement. Ainsi, l'insecte et la fleur bénéficient l'un de l'autre dans cette relation symbiotique. Le ver de l'épi du maïs et le plant Gaura entretiennent une relation similaire (Figure\(\PageIndex{2}\)).

Pollinisation par les chauv
Sous les tropiques et dans les déserts, les chauves-souris pollinisent souvent les fleurs nocturnes telles que l'agave, la goyave et la gloire matinale. Les fleurs sont généralement grandes et de couleur blanche ou pâle ; elles peuvent donc être distinguées de l'environnement sombre la nuit. Les fleurs ont un parfum puissant, fruité ou musqué et produisent de grandes quantités de nectar. Elles sont naturellement grandes et ont une bouche large pour accueillir la tête de la chauve-souris. Lorsque les chauves-souris cherchent le nectar, leur visage et leur tête se couvrent de pollen, qui est ensuite transféré sur la fleur suivante.
Pollinisation par les oiseaux
De nombreuses espèces de petits oiseaux, comme le colibri (Figure\(\PageIndex{3}\)) et les oiseaux solaires, pollinisent des plantes telles que les orchidées et autres fleurs sauvages. Les fleurs visitées par les oiseaux sont généralement robustes et orientées de manière à permettre aux oiseaux de rester près de la fleur sans que leurs ailes ne s'emmêlent dans les fleurs voisines. La fleur a généralement une forme tubulaire incurvée, ce qui permet d'accéder au bec de l'oiseau. Les fleurs aux couleurs vives et inodores qui s'ouvrent pendant la journée sont pollinisées par les oiseaux. Lorsqu'un oiseau cherche un nectar riche en énergie, le pollen se dépose sur la tête et le cou de l'oiseau, puis est transféré à la fleur suivante qu'il visite. Les botanistes sont connus pour déterminer l'aire de répartition des plantes éteintes en collectant et en identifiant le pollen de spécimens d'oiseaux âgés de 200 ans provenant du même site.

Pollinisation par le vent
La plupart des espèces de conifères et de nombreux angiospermes, tels que les graminées, les érables et les chênes, sont pollinisés par le vent. Les pommes de pin sont brunes et non parfumées, tandis que les fleurs des espèces d'angiospermes pollinisées par le vent sont généralement vertes, petites, peuvent avoir de petits pétales ou pas de pétales et produisent de grandes quantités de pollen. Contrairement aux fleurs typiques pollinisées par les insectes, les fleurs adaptées à la pollinisation par le vent ne produisent ni nectar ni odeur. Chez les espèces pollinisées par le vent, les microsporanges se détachent de la fleur et, lorsque le vent souffle, le pollen léger est transporté avec elle (Figure\(\PageIndex{4}\)). Les fleurs apparaissent généralement au début du printemps, avant les feuilles, afin que celles-ci ne bloquent pas le mouvement du vent. Le pollen se dépose sur le stigmate plumeux exposé de la fleur (Figure\(\PageIndex{5}\)).


Pollinisation par l'eau
Certaines mauvaises herbes, comme les herbes marines australiennes et les mauvaises herbes des étangs, sont pollinisées par l'eau. Le pollen flotte sur l'eau et, lorsqu'il entre en contact avec la fleur, il se dépose à l'intérieur de la fleur.
Evolution Connection : Pollinisation par tromperie
Les orchidées sont des fleurs très appréciées, avec de nombreuses variétés rares (Figure\(\PageIndex{6}\)). Ils poussent dans divers habitats spécifiques, principalement sous les tropiques d'Asie, d'Amérique du Sud et d'Amérique centrale. Au moins 25 000 espèces d'orchidées ont été identifiées.

Les fleurs attirent souvent les pollinisateurs avec des récompenses alimentaires, sous forme de nectar. Cependant, certaines espèces d'orchidées font exception à cette norme : elles ont développé différentes méthodes pour attirer les pollinisateurs souhaités. Ils utilisent une méthode connue sous le nom de tromperie alimentaire, dans laquelle des couleurs vives et des parfums sont proposés, mais pas de nourriture. Anacamptis morio, communément appelée orchidée à ailes vertes, porte des fleurs d'un violet vif et dégage un fort parfum. Le bourdon, son principal pollinisateur, est attiré par la fleur en raison de sa forte odeur, qui indique généralement de la nourriture pour une abeille, et qu'il ramasse le pollen pour le transporter vers une autre fleur.
D'autres orchidées utilisent la tromperie sexuelle. Chiloglottis trapeziformis émet un composé qui a la même odeur que la phéromone émise par une guêpe femelle pour attirer les guêpes mâles. La guêpe mâle est attirée par l'odeur, atterrit sur la fleur d'orchidée et, ce faisant, transfère du pollen. Certaines orchidées, comme l'orchidée marteau australienne, utilisent l'odeur et la ruse visuelle dans une autre stratégie de tromperie sexuelle pour attirer les guêpes. La fleur de cette orchidée imite l'apparence d'une guêpe femelle et émet une phéromone. La guêpe mâle tente de s'accoupler avec ce qui semble être une guêpe femelle et, ce faisant, ramasse du pollen, qu'elle transfère ensuite au partenaire contrefait suivant.
Double fertilisation
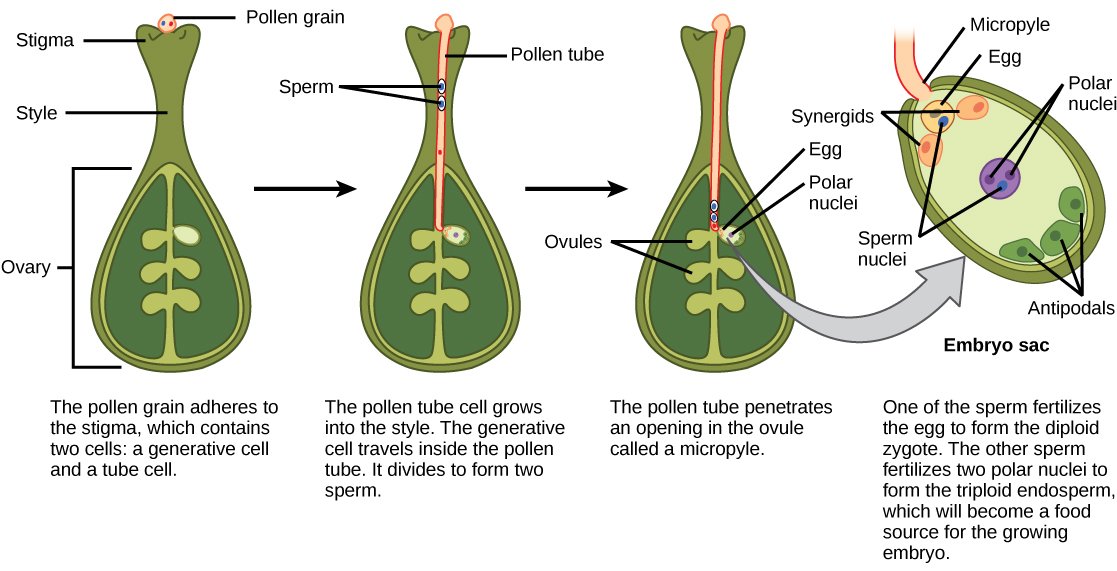
Une fois que le pollen s'est déposé sur le stigmate, il doit germer et se développer selon le style pour atteindre l'ovule. Les microspores, ou pollen, contiennent deux cellules : la cellule du tube pollinique et la cellule générative. La cellule du tube pollinique se développe dans un tube pollinique à travers lequel se déplace la cellule génératrice. La germination du tube pollinique nécessite de l'eau, de l'oxygène et certains signaux chimiques. Au fur et à mesure qu'il parcourt le style pour atteindre le sac embryonnaire, la croissance du tube pollinique est soutenue par les tissus du style. Entre-temps, si la cellule générative ne s'est pas déjà divisée en deux cellules, elle se divise maintenant pour former deux spermatozoïdes. Le tube pollinique est guidé par les substances chimiques sécrétées par les synergides présentes dans le sac embryonnaire et pénètre dans le sac ovulaire par le micropyle. Parmi les deux spermatozoïdes, l'un des spermatozoïdes féconde l'ovule pour former un zygote diploïde ; l'autre fusionne avec les deux noyaux polaires pour former une cellule triploïde qui se développe dans l'endosperme. Ensemble, ces deux événements de fécondation chez les angiospermes sont connus sous le nom de double fécondation (Figure\(\PageIndex{7}\)). Une fois la fécondation terminée, aucun autre sperme ne peut entrer. L'ovule fécondé forme la graine, tandis que les tissus de l'ovaire deviennent le fruit, enveloppant généralement la graine.
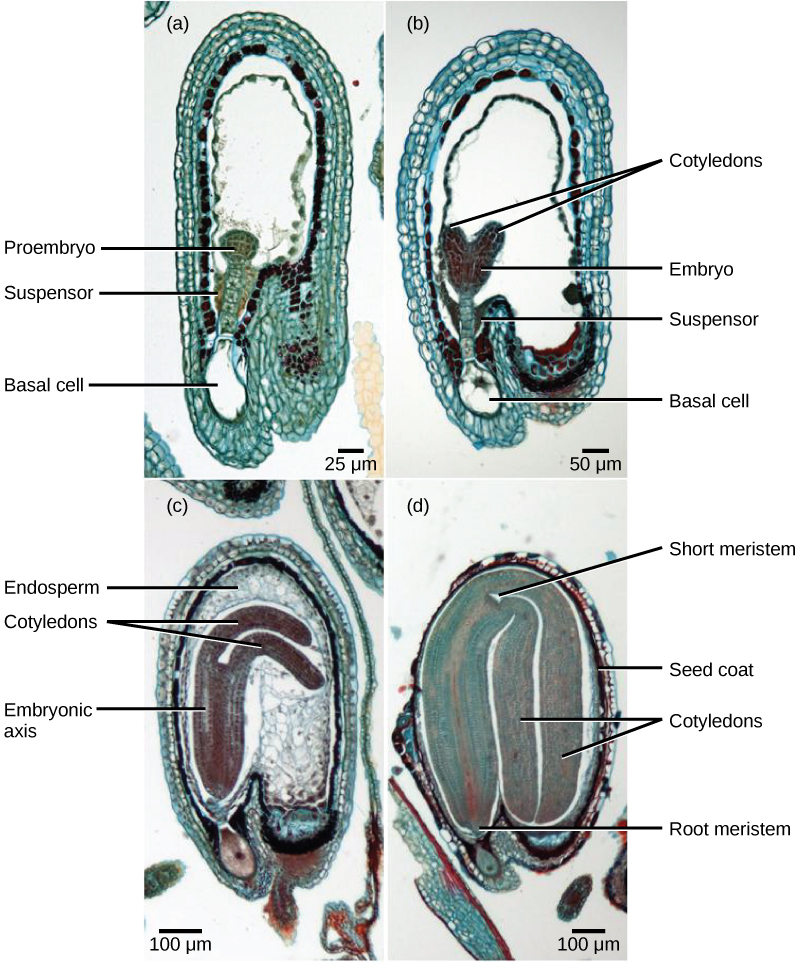
Après la fécondation, le zygote se divise pour former deux cellules : la cellule supérieure, ou cellule terminale, et la cellule inférieure, ou cellule basale. La division de la cellule basale donne naissance au suspenseur, qui finit par se connecter au tissu maternel. Le dispositif de suspension permet de transporter les nutriments de la plante mère vers l'embryon en croissance. La cellule terminale se divise également, donnant naissance à un proembryon de forme globulaire (Figure\(\PageIndex{8}\) a). Chez les dicotylédons (eudicots), l'embryon en développement a une forme de cœur, en raison de la présence des deux cotylédons rudimentaires (Figure\(\PageIndex{8}\) b). Chez les dicotylédons non endospermiques, tels que Capsella bursa, l'endosperme se développe initialement, puis est digéré et les réserves alimentaires sont transférées dans les deux cotylédons. À mesure que l'embryon et les cotylédons grossissent, ils perdent de la place à l'intérieur de la graine en développement et sont forcés de se plier (Figure\(\PageIndex{8}\) c). Finalement, l'embryon et les cotylédons remplissent la graine (Figure\(\PageIndex{8}\) d), et la graine est prête à être dispersée. Le développement embryonnaire est suspendu après un certain temps et la croissance ne reprend que lorsque la graine germe. Les plantules en développement dépendront des réserves alimentaires stockées dans les cotylédons jusqu'à ce que la première série de feuilles commence la photosynthèse.
Développement de la graine
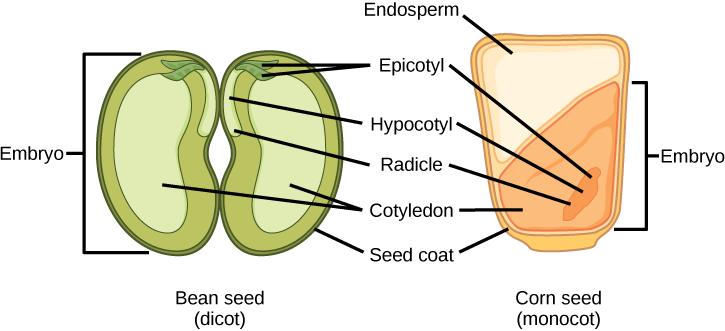
L'ovule mature se développe pour former la graine. Une graine typique contient un tégument, des cotylédons, un endosperme et un seul embryon (Figure\(\PageIndex{9}\)).
Art Connection
Laquelle des affirmations suivantes est vraie ?
- Les monocottes et les dicotylédones possèdent un endosperme.
- La radicule se développe jusqu'à la racine.
- Le plumule fait partie de l'épicotyle.
- L'endosperme fait partie de l'embryon.
Le stockage des réserves alimentaires dans les graines d'angiospermes diffère entre les monocottes et les dicotylédones. Chez les monocotylédons, comme le maïs et le blé, le cotylédon unique est appelé scutellum ; le scutellum est directement relié à l'embryon par le biais du tissu vasculaire (xylème et phloème). Les réserves alimentaires sont stockées dans le grand endosperme. Lors de la germination, les enzymes sont sécrétées par l'aleurone, une seule couche de cellules située juste à l'intérieur du tégument qui entoure l'endosperme et l'embryon. Les enzymes dégradent les glucides, les protéines et les lipides stockés, dont les produits sont absorbés par le scutellum et transportés par un canal vasculaire vers l'embryon en développement. Par conséquent, le scutellum peut être considéré comme un organe absorbant et non comme un organe de stockage.
Les deux cotylédons de la graine de dicot ont également des connexions vasculaires avec l'embryon. Dans les dicotys endospermiques, les réserves alimentaires sont stockées dans l'endosperme. Lors de la germination, les deux cotylédons agissent donc comme des organes absorbants pour absorber les réserves alimentaires libérées par voie enzymatique, un peu comme chez les monocotylédons (les monocotylédons, par définition, possèdent également des graines endospermiques). Le tabac (Nicotiana tabaccum), la tomate (Solanum lycopersicum) et le poivre (Capsicum annuum) sont des exemples de dicotylédones endospermiques. Chez les dicotylédons non endospermiques, l'endosperme triploïde se développe normalement après une double fécondation, mais les réserves alimentaires de l'endosperme sont rapidement remobilisées et transférées dans le cotylédon en développement pour y être stockées. Les deux moitiés d'une graine d'arachide (Arachis hypogaea) et des pois cassés (Pisum sativum) d'une soupe aux pois cassés sont des cotylédons individuels chargés de réserves alimentaires.
La graine, tout comme l'ovule, est protégée par un tégument formé à partir des téguments du sac ovulaire. Chez les dicotylédones, le tégument est ensuite divisé en une couche externe appelée testa et une couche interne connue sous le nom de tegmen.
L'axe embryonnaire comprend trois parties : la plumule, la radicule et l'hypocotyle. La partie de l'embryon située entre le point d'attache du cotylédon et la radicule est appelée hypocotyle (hypocotyle signifie « sous les cotylédons »). L'axe embryonnaire se termine par une radicule (la racine embryonnaire), qui est la région à partir de laquelle la racine se développera. Chez les dicotyles, les hypocotyles s'étendent au-dessus du sol et donnent naissance à la tige de la plante. Chez les monocotyles, l'hypocotyle n'apparaît pas au-dessus du sol car les monocotyles ne présentent pas d'allongement de la tige. La partie de l'axe embryonnaire qui fait saillie au-dessus des cotylédons est connue sous le nom d'épicotyle. Le plumule est composé de l'épicotyle, de jeunes feuilles et du méristème apical des pousses.
Lors de la germination dans les graines de dicotyles, l'épicotyle prend la forme d'un crochet dont le plumule est dirigé vers le bas. Cette forme est appelée crochet à plumules, et elle persiste tant que la germination se fait dans l'obscurité. Par conséquent, lorsque l'épicotyle traverse le sol dur et abrasif, le plumule est protégé contre les dommages. Lorsqu'il est exposé à la lumière, le crochet hypocotyle se redresse, les jeunes feuilles du feuillage font face au soleil et se dilatent, et l'épicotyle continue de s'allonger. Pendant ce temps, la radicule se développe également et produit la racine primaire. À mesure qu'il pousse vers le bas pour former la racine pivotante, les racines latérales se ramifient sur tous les côtés, produisant le système racinaire typique du dicotylédone.
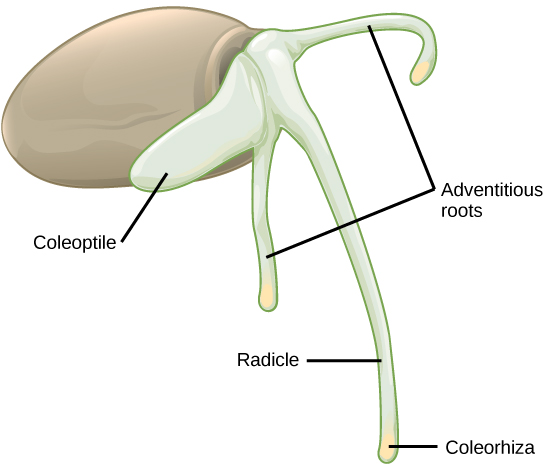
Dans les graines monocotylédones (Figure\(\PageIndex{10}\)), les testicules et les tegmen du tégument sont fusionnés. Au fur et à mesure que la graine germe, la racine primaire émerge, protégée par le revêtement de l'extrémité de la racine : la coléorhiza. Ensuite, la pousse primaire émerge, protégée par le coléoptile : le revêtement de l'extrémité de la pousse. Lors de l'exposition à la lumière (c'est-à-dire lorsque le plumule a quitté le sol et que le coléoptile protecteur n'est plus nécessaire), l'allongement du coléoptile cesse et les feuilles se dilatent et se déploient. À l'autre extrémité de l'axe embryonnaire, la racine primaire meurt rapidement, tandis que d'autres racines adventives (racines qui ne proviennent pas de l'endroit habituel, c'est-à-dire la racine) émergent de la base de la tige. Cela donne à la monocotylédone un système racinaire fibreux.
Germation des graines
De nombreuses graines mûres entrent dans une période d'inactivité ou d'activité métabolique extrêmement faible : un processus connu sous le nom de dormance, qui peut durer des mois, des années, voire des siècles. La dormance aide à maintenir la viabilité des graines dans des conditions défavorables. Lors du retour à des conditions favorables, la germination des graines a lieu. Les conditions favorables peuvent être aussi diverses que l'humidité, la lumière, le froid, le feu ou les traitements chimiques. Après de fortes pluies, de nombreux nouveaux plants apparaissent. Les feux de forêt entraînent également l'émergence de nouveaux plants. Certaines graines doivent être vernalisées (traitement au froid) avant de pouvoir germer. Cela garantit que les graines produites par les plantes des climats tempérés ne germeront pas avant le printemps. Les plantes qui poussent dans des climats chauds peuvent avoir des graines qui nécessitent un traitement thermique pour germer, afin d'éviter la germination pendant les étés chauds et secs. Dans de nombreuses graines, la présence d'un tégument épais retarde la capacité de germination. La scarification, qui comprend des processus mécaniques ou chimiques visant à ramollir le tégument, est souvent utilisée avant la germination. Le prétrempage dans de l'eau chaude ou le passage dans un environnement acide, tel que le tube digestif d'un animal, peut également être utilisé.
Selon la taille des graines, le temps nécessaire à l'émergence d'un plant peut varier. Les espèces à grosses graines disposent de suffisamment de réserves alimentaires pour germer profondément sous terre tout en étendant leur épicotyle jusqu'à la surface du sol. Les graines des espèces à petites graines ont généralement besoin de lumière comme signal de germination. Cela garantit que les graines ne germent qu'à la surface du sol ou à proximité de celle-ci (là où la lumière est la plus intense). S'ils devaient germer trop loin sous la surface, les plantules en développement n'auraient pas suffisamment de réserves alimentaires pour atteindre la lumière du soleil.
Développement des fruits et des types de fruits
Après la fécondation, l'ovaire de la fleur se transforme généralement en fruit. Les fruits sont généralement associés à un goût sucré ; cependant, tous les fruits ne sont pas sucrés. Sur le plan botanique, le terme « fruit » est utilisé pour désigner un ovaire mûr. Dans la plupart des cas, les fleurs fécondées se transformeront en fruits, et les fleurs dont la fertilisation n'a pas eu lieu ne le seront pas. Certains fruits se développent à partir de l'ovaire et sont considérés comme de vrais fruits, tandis que d'autres se développent à partir d'autres parties du gamétophyte femelle et sont connus sous le nom de fruits accessoires. Le fruit renferme les graines et l'embryon en développement, le protégeant ainsi. Les fruits sont de nombreux types, selon leur origine et leur texture. Le tissu sucré de la mûre, la chair rouge de la tomate, la coque de l'arachide et la coque du maïs (la partie dure et fine qui se coince dans les dents lorsque vous mangez du pop-corn) sont tous des fruits. À mesure que le fruit mûrit, les graines mûrissent également.
Les fruits peuvent être classés comme simples, agrégés, multiples ou accessoires, selon leur origine (Figure\(\PageIndex{11}\)). Si le fruit se développe à partir d'un seul carpelle ou de carpelles fusionnés d'un seul ovaire, on parle de fruit simple, comme on le voit dans les noix et les haricots. Un fruit agrégé est un fruit qui se développe à partir de plusieurs carpelles, mais qui se trouvent tous dans la même fleur : les carpelles matures fusionnent pour former le fruit entier, comme on le voit sur la framboise. De multiples fruits se développent à partir d'une inflorescence ou d'une grappe de fleurs. Un exemple est l'ananas, où les fleurs fusionnent pour former le fruit. Les fruits accessoires (parfois appelés faux fruits) ne proviennent pas de l'ovaire, mais d'une autre partie de la fleur, comme le réceptacle (fraise) ou l'hypanthium (pommes et poires).

Les fruits se composent généralement de trois parties : l'exocarpe (la peau ou le revêtement externe), le mésocarpe (partie centrale du fruit) et l'endocarpe (partie interne du fruit). Ensemble, les trois sont connus sous le nom de péricarpe. Le mésocarpe est généralement la partie charnue et comestible du fruit ; cependant, dans certains fruits, comme l'amande, l'endocarpe est la partie comestible. Dans de nombreux fruits, deux ou les trois couches sont fusionnées et ne peuvent pas être distinguées à maturité. Les fruits peuvent être secs ou charnus. De plus, les fruits peuvent être divisés en types déhiscents ou indéhiscents. Les fruits déhiscents, tels que les pois, libèrent facilement leurs graines, tandis que les fruits indéhiscents, comme les pêches, dépendent de la pourriture pour libérer leurs graines.
Dispersion des fruits et des graines
Le fruit n'a qu'un seul but : disperser les graines. Les graines contenues dans les fruits doivent être dispersées loin de la plante mère afin qu'elles puissent trouver des conditions favorables et moins compétitives pour germer et pousser.
Certains fruits possèdent des mécanismes intégrés qui leur permettent de se disperser d'eux-mêmes, tandis que d'autres ont besoin de l'aide d'agents tels que le vent, l'eau et les animaux (Figure\(\PageIndex{12}\)). Les modifications de la structure, de la composition et de la taille des graines contribuent à la dispersion. Les fruits dispersés par le vent sont légers et peuvent avoir des appendices en forme d'ailes qui leur permettent d'être transportés par le vent. Certains ont une structure semblable à un parachute pour les maintenir à flot. Certains fruits, par exemple le pissenlit, ont une structure velue et légère qui se prête à la dispersion par le vent.
Les graines dispersées par l'eau sont contenues dans des fruits légers et flottants, ce qui leur donne la capacité de flotter. Les noix de coco sont bien connues pour leur capacité à flotter sur l'eau pour atteindre les terres où elles peuvent germer. De même, les saules et les bouleaux argentés produisent des fruits légers qui peuvent flotter sur l'eau.
Les animaux et les oiseaux mangent des fruits et les graines qui ne sont pas digérées sont excrétées dans leurs excréments à une certaine distance. Certains animaux, comme les écureuils, enfouissent les fruits contenant des graines pour une utilisation ultérieure ; si l'écureuil ne trouve pas sa réserve de fruits et si les conditions sont favorables, les graines germent. Certains fruits, comme le lambeau, possèdent des crochets ou des structures collantes qui adhèrent au pelage de l'animal et sont ensuite transportés vers un autre endroit. Les humains jouent également un rôle important dans la dispersion des graines lorsqu'ils transportent des fruits vers de nouveaux endroits et jettent la partie non comestible qui les contient.
Tous les mécanismes ci-dessus permettent de disperser les graines dans l'espace, tout comme la progéniture d'un animal peut se déplacer vers un nouvel endroit. La dormance des graines, décrite précédemment, permet aux plantes de disperser leur progéniture dans le temps, ce que les animaux ne peuvent pas faire. Les graines dormantes peuvent attendre des mois, des années, voire des décennies pour que les conditions nécessaires à la germination et à la propagation de l'espèce soient réunies.



Résumé
Pour que la fécondation se produise chez les angiospermes, le pollen doit être transféré vers le stigmate de la fleur : un processus connu sous le nom de pollinisation. La pollinisation par les gymnospermatozoïdes implique le transfert du pollen d'un cône mâle vers un cône femelle. Lorsque le pollen de la fleur est transféré sur le stigmate de la même fleur, on parle d'autopollinisation. La pollinisation croisée se produit lorsque le pollen est transféré d'une fleur à une autre fleur de la même plante ou d'une autre plante. La pollinisation croisée nécessite des agents pollinisateurs tels que l'eau, le vent ou les animaux, et augmente la diversité génétique. Une fois que le pollen a atterri sur le stigmate, la cellule tubulaire donne naissance au tube pollinique, à travers lequel le noyau générateur migre. Le tube pollinique pénètre par le micropyle situé dans le sac ovulaire. La cellule générative se divise pour former deux spermatozoïdes : l'un fusionne avec l'ovule pour former le zygote diploïde, et l'autre fusionne avec les noyaux polaires pour former l'endosperme, qui est de nature triploïde. C'est ce que l'on appelle la double fertilisation. Après la fécondation, le zygote se divise pour former l'embryon et l'ovule fécondé forme la graine. Les parois de l'ovaire forment le fruit dans lequel se développent les graines. La graine, une fois arrivée à maturité, germera dans des conditions favorables et donnera naissance au sporophyte diploïde.
Connexions artistiques
Figure\(\PageIndex{9}\) : Quelle est la fonction du cotylédon ?
- Il se développe jusqu'à la racine.
- Il fournit de la nutrition à l'embryon.
- Il forme l'embryon.
- Elle protège l'embryon.
- Réponse
-
B
Lexique
- fruits accessoires
- fruit dérivé de tissus autres que l'ovaire
- fruits agrégés
- fruit qui se développe à partir de plusieurs carpelles d'une même fleur
- aleurone
- couche unique de cellules située juste à l'intérieur du tégument qui sécrète des enzymes lors de la germination
- coléoptile
- couverture de l'extrémité des pousses, présente dans les graines de monocotylédones en germination
- coléorhize
- couverture de l'extrémité des racines, présente dans les graines de monocotylédones en germination
- cotylédon
- partie charnue de la graine qui fournit de la nutrition à la graine
- pollinisation croisée
- transfert du pollen de l'anthère d'une fleur au stigmate d'une autre fleur
- dormance
- période d'absence de croissance et de processus métaboliques très lents
- double fertilisation
- deux événements de fécondation chez les angiospermes : l'un des spermatozoïdes fusionne avec l'ovule pour former le zygote, tandis que l'autre fusionne avec les noyaux polaires pour former l'endosperme
- endocarpe
- partie la plus interne du fruit
- endosperme
- structure triploïde résultant de la fusion d'un spermatozoïde avec des noyaux polaires, qui sert de tissu nutritif pour l'embryon
- dicot endospermique
- dicot qui stocke les réserves alimentaires dans l'endosperme
- exocarpe
- enveloppe extérieure d'un fruit
- épicotyle
- pousse embryonnaire au-dessus des cotylédons
- gravitropisme
- réponse de la croissance d'une plante dans le même sens que la gravité
- hypocotyle
- axe embryonnaire au-dessus des cotylédons
- mésocarpe
- partie centrale d'un fruit
- fruits multiples
- fruit qui se développe à partir de plusieurs fleurs sur une inflorescence
- guide du nectar
- motif pigmentaire sur une fleur qui guide un insecte vers les nectaires
- dicotylédone non endospermique
- dicot qui stocke les réserves alimentaires dans le cotylédon en développement
- péricarpe
- terme collectif décrivant l'exocarpe, le mésocarpe et l'endocarpe ; la structure qui entoure la graine et fait partie du fruit
- plumule
- pousse qui se développe à partir de la graine en germination
- pollinisation
- transfert du pollen vers le stigmate
- radicule
- racine originale qui se développe à partir de la graine en germination
- scarification
- procédés mécaniques ou chimiques visant à ramollir le tégument
- scutellum
- type de cotylédon présent dans les monocottes, comme dans les graines de graminées
- autopollinisation
- transfert du pollen de l'anthère au stigmate de la même fleur
- fruit simple
- fruit qui se développe à partir d'un seul carpelle ou de carpelles fusionnés
- suspenseur
- partie de l'embryon en croissance qui se connecte aux tissus maternels
- tegmen
- couche interne du tégument
- testa
- couche externe du tégument
- vernalisation
- exposition au froid requise par certaines graines avant qu'elles ne puissent germer