
23.3 : Groupes de protistes
- Page ID
- 189864

Compétences à développer
- Décrire les organismes protistes représentatifs de chacun des six supergroupes d'eucaryotes actuellement reconnus
- Identifier les relations évolutives entre les plantes, les animaux et les champignons au sein des six supergroupes d'eucaryotes actuellement reconnus
En l'espace de plusieurs décennies, le Royaume de Protista a été démantelé car des analyses de séquences ont révélé de nouvelles relations génétiques (et donc évolutives) entre ces eucaryotes. De plus, les protistes qui présentent des caractéristiques morphologiques similaires peuvent avoir développé des structures analogues en raison de pressions sélectives similaires, plutôt qu'en raison d'une ascendance commune récente. Ce phénomène, appelé évolution convergente, est l'une des raisons pour lesquelles la classification des protistes est si difficile. Le schéma de classification émergent regroupe l'ensemble du domaine Eucaryote en six « supergroupes » qui contiennent tous les protistes ainsi que les animaux, les plantes et les champignons issus d'un ancêtre commun (Figure\(\PageIndex{1}\)). Les supergroupes sont considérés comme monophylétiques, ce qui signifie que tous les organismes de chaque supergroupe sont censés avoir évolué à partir d'un seul ancêtre commun, et que tous les membres sont donc plus étroitement liés les uns aux autres qu'aux organismes extérieurs à ce groupe. Il n'existe toujours pas de preuves de la monophylie de certains groupes.
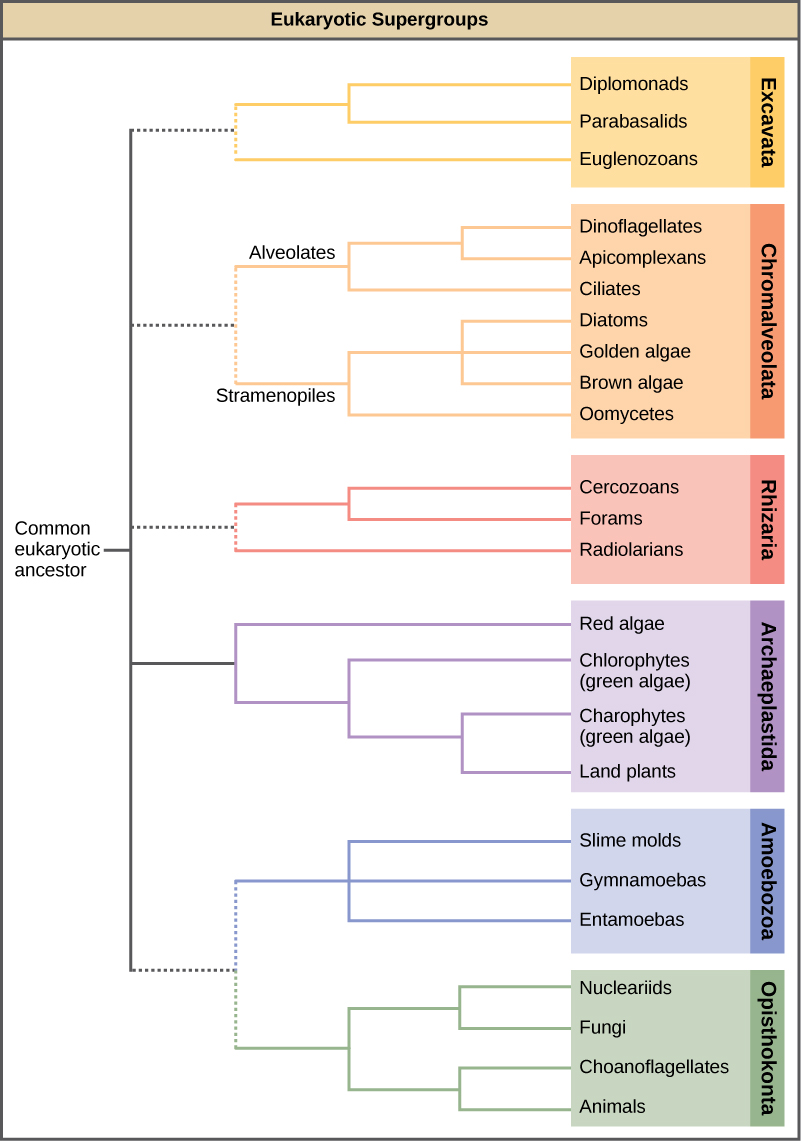
La classification des eucaryotes est toujours changeante et les six supergroupes pourraient être modifiés ou remplacés par une hiérarchie plus appropriée au fur et à mesure que les données génétiques, morphologiques et écologiques s'accumulent. Gardez à l'esprit que le schéma de classification présenté ici n'est qu'une des nombreuses hypothèses et que les véritables relations évolutives restent à déterminer. Lorsque vous découvrez les protistes, il est utile de se concentrer moins sur la nomenclature et davantage sur les points communs et les différences qui définissent les groupes eux-mêmes.
Excavata
De nombreuses espèces de protistes classées dans le supergroupe Excavata sont des organismes unicellulaires asymétriques dotés d'un canal d'alimentation « creusé » d'un côté. Ce supergroupe comprend des prédateurs hétérotrophes, des espèces photosynthétiques et des parasites. Ses sous-groupes sont les diplomonades, les parabasalides et les euglénozoaires.
Diplomas et publicités
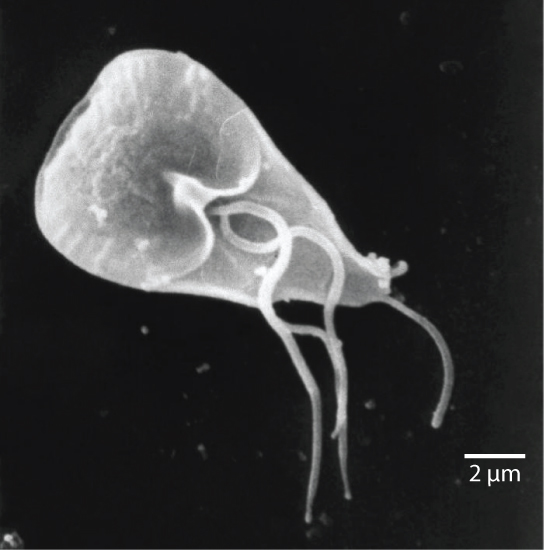
Parmi les Excavata se trouvent les diplomonades, dont le parasite intestinal Giardia lamblia (Figure\(\PageIndex{2}\)). Jusqu'à récemment, on pensait que ces protistes étaient dépourvus de mitochondries. Des organites mitochondriaux résiduels, appelés mitosomes, ont depuis été identifiés dans des diplomonades, mais ces mitosomes sont essentiellement non fonctionnels. Les diplomonades existent dans des environnements anaérobies et utilisent des voies alternatives, telles que la glycolyse, pour générer de l'énergie. Chaque cellule diplomonade possède deux noyaux identiques et utilise plusieurs flagelles pour la locomotion.
Parabasalides
Un deuxième sous-groupe d'Excavata, les parabasalides, présente également des mitochondries semi-fonctionnelles. Chez les parabasalides, ces structures fonctionnent de manière anaérobie et sont appelées hydrogénosomes parce qu'elles produisent de l'hydrogène en tant que sous-produit. Les parabasalides se déplacent avec les flagelles et les ondulations de la membrane. Trichomonas vaginalis, un parabasalide responsable d'une maladie sexuellement transmissible chez l'homme, utilise ces mécanismes pour transiter par les voies urogénitales des hommes et des femmes. T. le vaginalis provoque la trichamonase, qui apparaît dans environ 180 millions de cas dans le monde chaque année. Alors que les hommes présentent rarement des symptômes lors d'une infection par ce protiste, les femmes infectées peuvent devenir plus vulnérables à une infection secondaire par le virus de l'immunodéficience humaine (VIH) et peuvent être plus susceptibles de développer un cancer du col de l'utérus. Femmes enceintes infectées par T. les vaginaux présentent un risque accru de complications graves, telles qu'un accouchement prématuré.
Euglénozoaires
Les euglénozoaires comprennent les parasites, les hétérotrophes, les autotrophes et les mixotrophes, dont la taille varie de 10 à 500 µm. Les euglénoïdes se déplacent dans leur habitat aquatique à l'aide de deux longs flagelles qui les guident vers des sources de lumière détectées par un organe oculaire primitif appelé tache oculaire. Le genre bien connu, Euglena, comprend certaines espèces mixotrophes qui ne présentent une capacité photosynthétique que lorsque la lumière est présente. Dans l'obscurité, les chloroplastes d'Euglena rétrécissent et cessent temporairement de fonctionner, et les cellules absorbent les nutriments organiques de leur environnement.
Le parasite humain, Trypanosoma brucei, appartient à un sous-groupe différent d'Euglénozoaires, les kinétoplastes. Le sous-groupe des kinétoplastes est nommé d'après le kinétoplaste, une masse d'ADN contenue dans la mitochondrie surdimensionnée unique que possède chacune de ces cellules. Ce sous-groupe comprend plusieurs parasites, appelés collectivement trypanosomes, qui provoquent des maladies humaines dévastatrices et infectent une espèce d'insecte pendant une partie de son cycle de vie. T. des brucei apparaissent dans l'intestin de la mouche tsé-tsé après que celle-ci a mordu un hôte humain ou un autre mammifère infecté. Le parasite se déplace ensuite vers les glandes salivaires de l'insecte pour être transmis à un autre humain ou à un autre mammifère lorsque la mouche tsé-tsé infectée consomme un autre repas de sang. T. Le brucei est fréquent en Afrique centrale et est l'agent causal de la maladie du sommeil en Afrique, une maladie associée à une fatigue chronique sévère, au coma, et qui peut être fatale si elle n'est pas traitée.
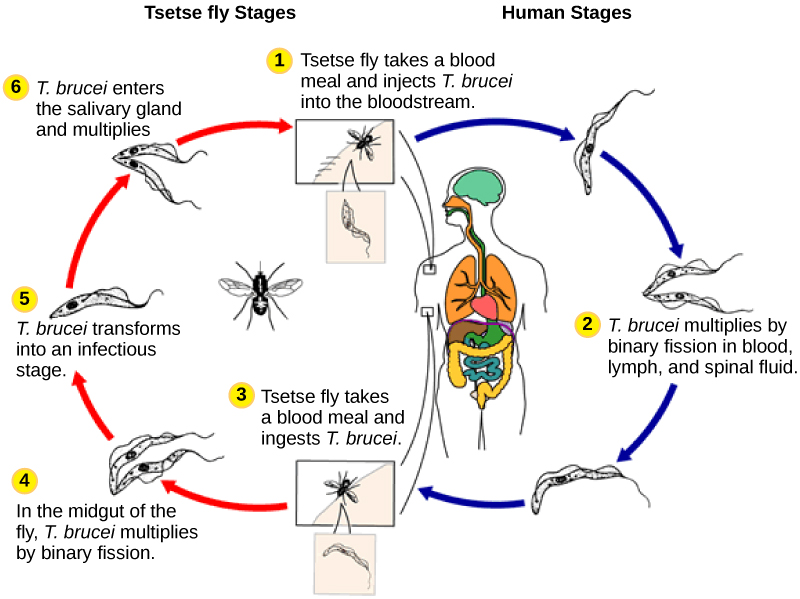
Trypanosoma brucei
Regardez cette vidéo pour voir T. brucei nager. https://youtu.be/EnsydwITLYk
Véolates chromatiques
Les données actuelles suggèrent que les espèces classées dans la catégorie des chromalvéolates sont dérivées d'un ancêtre commun qui a englouti une cellule d'algue rouge photosynthétique, qui avait elle-même déjà développé des chloroplastes à partir d'une relation endosymbiotique avec un procaryote photosynthétique. On pense donc que l'ancêtre des chromalvéolates est le résultat d'un événement endosymbiotique secondaire. Cependant, certains chromalvéolates semblent avoir perdu des organites plastidiques dérivés des algues rouges ou être totalement dépourvus de gènes plastidiques. Ce supergroupe doit donc être considéré comme un groupe de travail fondé sur des hypothèses et sujet à modification. Les chromalvéolates comprennent des organismes photosynthétiques très importants, tels que les diatomées, les algues brunes et d'importants agents pathogènes chez les animaux et les plantes. Les chromalvéolates peuvent être subdivisés en alvéolats et en stramenopiles.
Alvéolates : dinoflagellés, apicomplexiens et ciliés
De nombreuses données confirment que les alvéolés sont dérivés d'un ancêtre commun. Les alvéolats sont nommés d'après la présence d'une alvéole, ou d'un sac enveloppé d'une membrane, sous la membrane cellulaire. La fonction exacte de l'alvéole est inconnue, mais elle pourrait être impliquée dans l'osmorégulation. Les alvéolés sont ensuite classés selon certains des protistes les plus connus : les dinoflagellés, les apicomplexans et les ciliés.
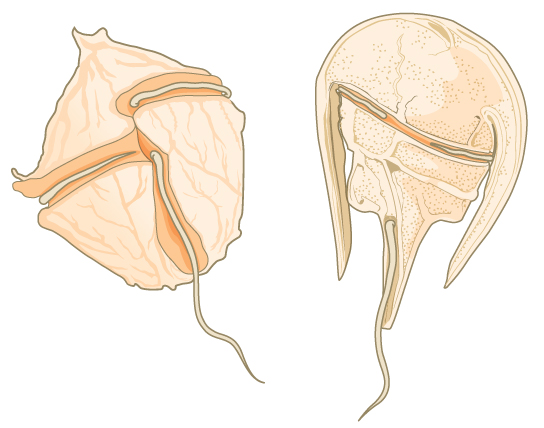
Les dinoflagellés présentent une grande diversité morphologique et peuvent être photosynthétiques, hétérotrophes ou mixotrophes. De nombreux dinoflagellés sont enfermés dans des plaques de cellulose imbriquées. Deux flagelles perpendiculaires s'insèrent dans les rainures entre les plaques de cellulose, un flagelle s'étendant longitudinalement et un second encerclant le dinoflagellé (Figure\(\PageIndex{4}\)). Ensemble, les flagelles contribuent au mouvement de rotation caractéristique des dinoflagellés. Ces protistes vivent dans des habitats d'eau douce et marins et font partie du plancton, des organismes typiquement microscopiques qui dérivent dans l'eau et constituent une source de nourriture cruciale pour les grands organismes aquatiques.
Certains dinoflagellés produisent de la lumière, appelée bioluminescence, lorsqu'ils sont sectionnés ou stressés. Un grand nombre de dinoflagellés marins (milliards ou billions de cellules par vague) peuvent émettre de la lumière et provoquer le scintillement d'une vague déferlante entière ou prendre une couleur bleue brillante (Figure\(\PageIndex{5}\)). Pour environ 20 espèces de dinoflagellés marins, les explosions de population (également appelées fleurs) pendant les mois d'été peuvent teinter l'océan d'une couleur rouge boueuse. Ce phénomène, appelé marée rouge, est dû à l'abondance de pigments rouges présents dans les plastes dinoflagellés. En grande quantité, ces espèces de dinoflagellées sécrètent une toxine asphyxiante qui peut tuer les poissons, les oiseaux et les mammifères marins. Les marées rouges peuvent nuire considérablement à la pêche commerciale et les humains qui consomment ces protistes peuvent être empoisonnés.

Les protistes apicomplexans sont ainsi nommés parce que leurs microtubules, leur fibrine et leurs vacuoles sont répartis de manière asymétrique à une extrémité de la cellule dans une structure appelée complexe apical (Figure\(\PageIndex{6}\)). Le complexe apical est spécialisé dans l'entrée et l'infection des cellules hôtes. En effet, tous les apicomplexans sont parasitaires. Ce groupe comprend le genre Plasmodium, responsable du paludisme chez l'homme. Les cycles de vie de l'apicomplexan sont complexes et impliquent de multiples hôtes et stades de reproduction sexuée et asexuée.
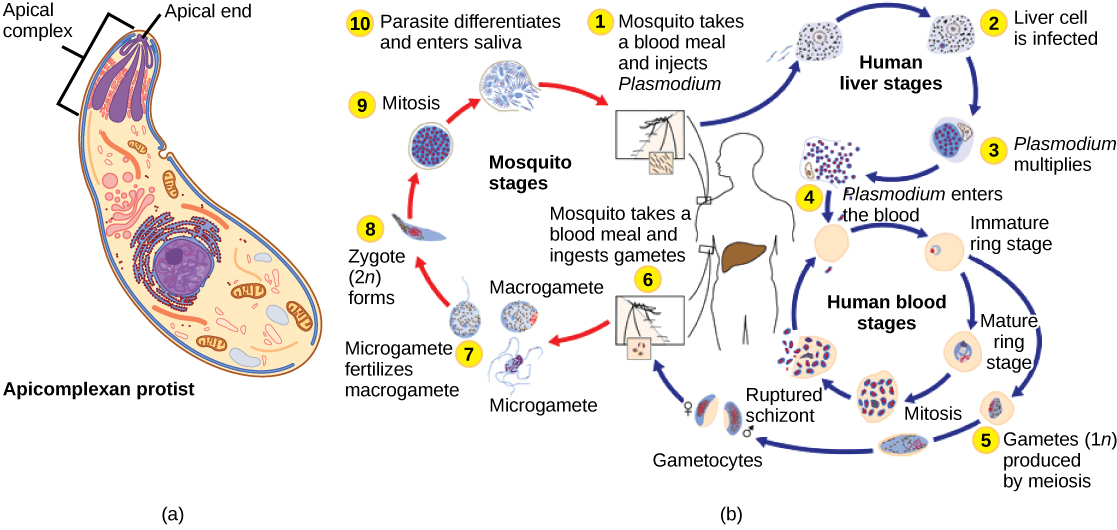
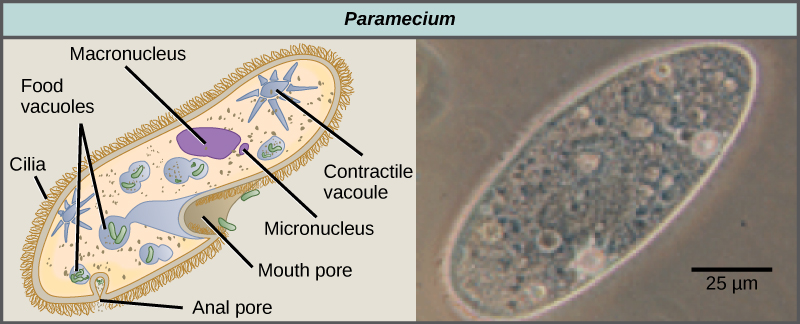
Les ciliés, qui comprennent Paramecium et Tetrahymena, sont un groupe de protistes de 10 à 3 000 micromètres de long recouverts de rangées, de touffes ou de spirales de minuscules cils. En battant leurs cils de manière synchrone ou par vagues, les ciliés peuvent coordonner leurs mouvements dirigés et ingérer des particules alimentaires. Certains ciliés possèdent des structures fusionnées à base de cils qui fonctionnent comme des pagaies, des entonnoirs ou des nageoires. Les ciliés sont également entourés d'une pellicule, offrant une protection sans compromettre l'agilité. Le genre Paramecium comprend des protistes qui ont organisé leurs cils en une bouche primitive en forme de plaque, appelée rainure buccale, utilisée pour capturer et digérer les bactéries (Figure\(\PageIndex{7}\)). Les aliments capturés dans le sillon buccal pénètrent dans une vacuole alimentaire, où ils se combinent aux enzymes digestives. Les particules de déchets sont expulsées par une vésicule exocytaire qui fusionne dans une région spécifique de la membrane cellulaire, appelée pore anal. En plus d'un système digestif basé sur des vacuoles, Paramecium utilise également des vacuoles contractiles, qui sont des vésicules osmorégulatrices qui se remplissent d'eau lorsqu'elle entre dans la cellule par osmose, puis se contractent pour extraire l'eau de la cellule.
Lien vers l'apprentissage
Regardez la vidéo de la vacuole contractile de Paramecium expulsant l'eau pour maintenir l'équilibre osmotique de la cellule.
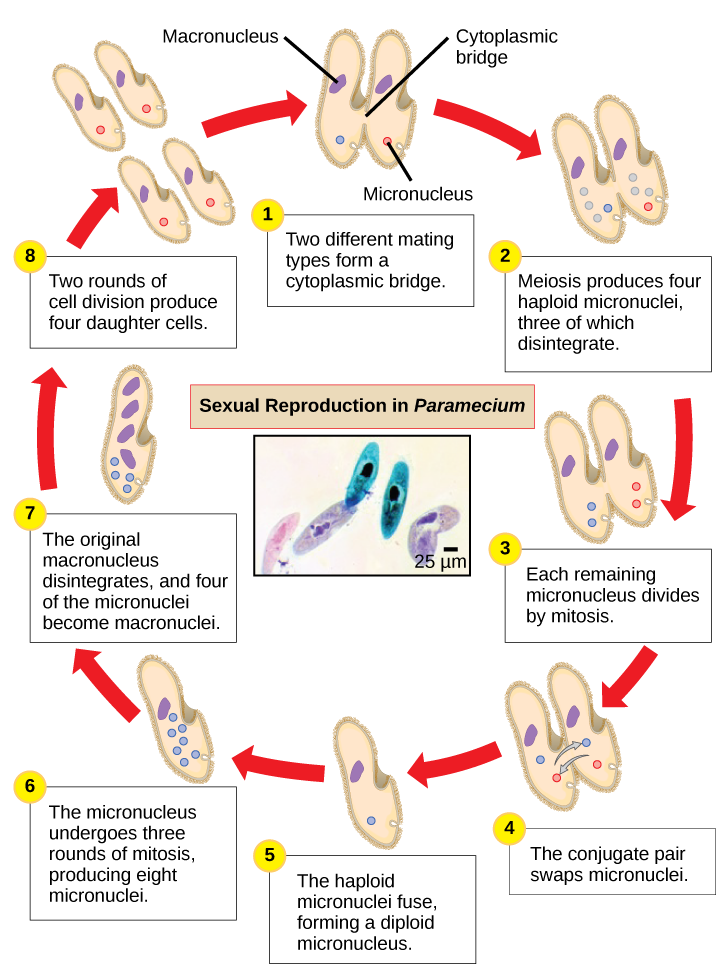
Le paramécium possède deux noyaux, un macronucleus et un micronoyau, dans chaque cellule. Le micronoyau est essentiel à la reproduction sexuée, tandis que le macronoyau dirige la fission binaire asexuée et toutes les autres fonctions biologiques. Le processus de reproduction sexuée chez Paramecium souligne l'importance du micronoyau pour ces protistes. Le paramécium et la plupart des autres ciliés se reproduisent sexuellement par conjugaison. Ce processus commence lorsque deux types d'accouplement différents de Paramecium entrent en contact physique et se rejoignent par un pont cytoplasmique (Figure\(\PageIndex{8}\)). Le micronoyau diploïde de chaque cellule subit ensuite une méiose pour produire quatre micronoyaux haploïdes. Trois d'entre elles dégénèrent dans chaque cellule, laissant un micronoyau qui subit ensuite une mitose, générant deux micronoyaux haploïdes. Les cellules échangent chacune l'un de ces noyaux haploïdes et s'éloignent l'une de l'autre. Un processus similaire se produit chez les bactéries qui possèdent des plasmides. La fusion des micronoyaux haploïdes génère un pré-micronoyau diploïde complètement nouveau dans chaque cellule conjuguée. Ce pré-micronoyau subit trois cycles de mitose pour produire huit copies, et le macronoyau d'origine se désintègre. Quatre des huit pré-micronoyaux deviennent des micronoyaux à part entière, tandis que les quatre autres effectuent plusieurs cycles de réplication de l'ADN pour devenir de nouveaux macronoyaux. Deux divisions cellulaires produisent ensuite quatre nouvelles paramécies à partir de chaque cellule conjugative d'origine.
Exercice
Laquelle des affirmations suivantes concernant la reproduction sexuée de Paramecium est fausse ?
- Les macronoyaux sont dérivés de micronoyaux.
- La mitose et la méiose se produisent pendant la reproduction sexuée.
- La paire conjuguée échange des macronucléii.
- Chaque parent produit quatre cellules filles.
Stramenopiles : diatomées, algues brunes, algues dorées et oomycètes
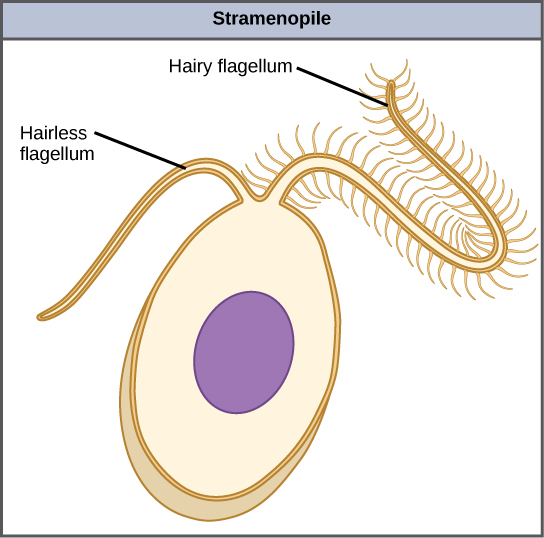
L'autre sous-groupe de chromalvéolates, les stramenopiles, comprend les algues marines photosynthétiques et les protistes hétérotrophes. La caractéristique unificatrice de ce groupe est la présence d'un flagelle texturé ou « velu ». De nombreux stramenopiles possèdent également un flagelle supplémentaire dépourvu de projections ressemblant à des cheveux (Figure\(\PageIndex{9}\)). La taille des membres de ce sous-groupe va des diatomées unicellulaires au varech massif et multicellulaire.
Les diatomées sont des protistes photosynthétiques unicellulaires qui s'enferment dans des parois cellulaires vitreuses aux motifs complexes composées de dioxyde de silicium dans une matrice de particules organiques (Figure\(\PageIndex{10}\)). Ces protistes font partie du plancton d'eau douce et marin. La plupart des espèces de diatomées se reproduisent de manière asexuée, bien que certains cas de reproduction sexuée et de sporulation existent également. Certaines diatomées présentent une fente dans leur coque de silice, appelée raphé. En expulsant un flux de mucopolysaccharides du raphé, la diatomée peut se fixer aux surfaces ou se propulser dans une direction.
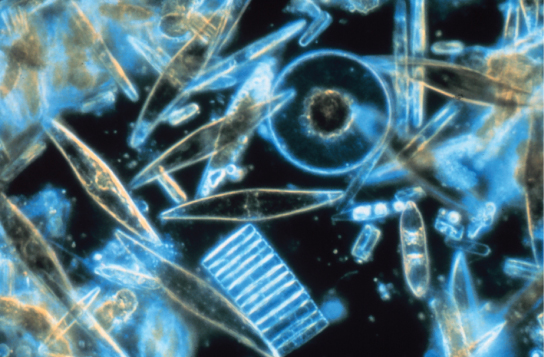
Pendant les périodes de disponibilité des nutriments, les populations de diatomées fleurissent en quantité supérieure à celle que peuvent consommer les organismes aquatiques. Les diatomées excédentaires meurent et s'enfoncent dans le fond marin, où elles ne sont pas facilement accessibles par les sasondes qui se nourrissent d'organismes morts. Par conséquent, le dioxyde de carbone que les diatomées avaient consommé et incorporé dans leurs cellules lors de la photosynthèse n'est pas renvoyé dans l'atmosphère. En général, ce processus par lequel le carbone est transporté profondément dans l'océan est décrit comme la pompe biologique au carbone, car le carbone est « pompé » vers les profondeurs de l'océan où il est inaccessible à l'atmosphère sous forme de dioxyde de carbone. La pompe à charbon biologique est un élément crucial du cycle du carbone qui permet de maintenir des niveaux de dioxyde de carbone atmosphériques plus faibles.
Comme les diatomées, les algues dorées sont en grande partie unicellulaires, bien que certaines espèces puissent former de grandes colonies. Leur couleur dorée caractéristique résulte de leur utilisation intensive de caroténoïdes, un groupe de pigments photosynthétiques généralement de couleur jaune ou orange. Les algues dorées se trouvent à la fois dans les environnements d'eau douce et marins, où elles constituent une partie importante de la communauté planctonique.
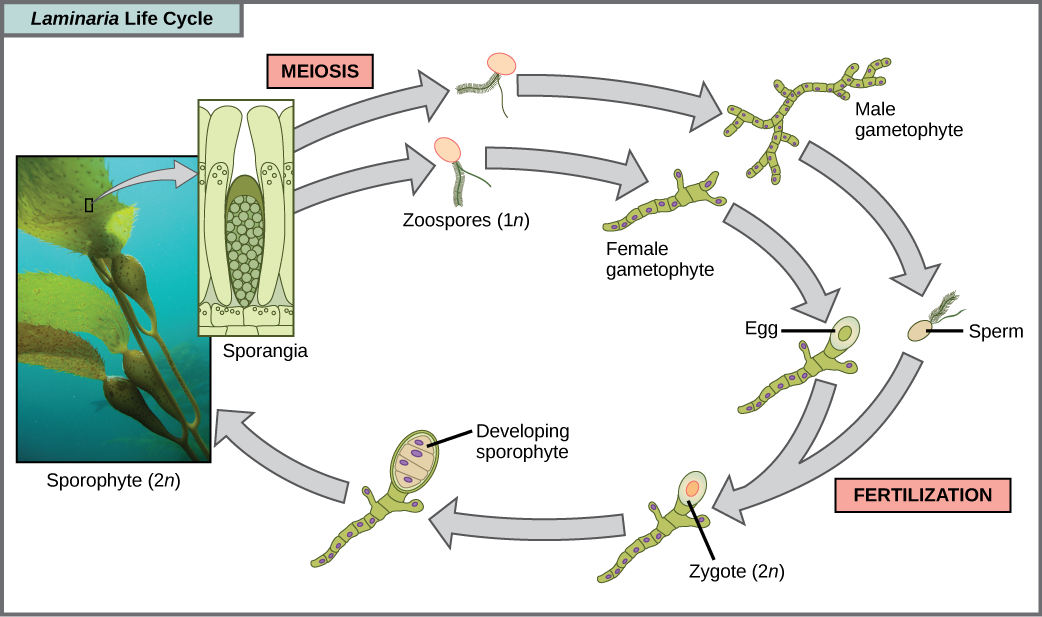
Les algues brunes sont principalement des organismes marins multicellulaires connus sous le nom d'algues marines. Le varech géant est un type d'algue brune. Certaines algues brunes ont développé des tissus spécialisés qui ressemblent à des plantes terrestres, avec des reliefs semblables à des racines, des stipes ressemblant à des tiges et des limbes foliaires capables de photosynthèse. Les raies de varech géants sont énormes et s'étendent dans certains cas sur 60 mètres. Les cycles de vie des algues sont variés, mais le plus complexe est l'alternance de générations, au cours de laquelle les stades haploïdes et diploïdes impliquent la multicellularité. Comparez ce cycle de vie à celui des humains, par exemple. Les gamètes haploïdes produits par la méiose (spermatozoïdes et ovules) se combinent lors de la fécondation pour générer un zygote diploïde qui subit de nombreuses mitoses pour produire un embryon multicellulaire puis un fœtus. Cependant, les spermatozoïdes et les ovules eux-mêmes ne deviennent jamais des êtres multicellulaires. Les plantes terrestres ont également évolué en alternance de générations. Dans le genre d'algues brunes Laminaria, les spores haploïdes se transforment en gamétophytes multicellulaires, qui produisent des gamètes haploïdes qui se combinent pour produire des organismes diploïdes qui deviennent ensuite des organismes multicellulaires ayant une structure différente de celle de la forme haploïde (Figure\(\PageIndex{11}\)). Certains autres organismes effectuent une alternance de générations dans lesquelles les formes haploïdes et diploïdes se ressemblent.
Exercice
Laquelle des affirmations suivantes concernant le cycle de vie des laminaires est fausse ?
- 1 n zoospores se forment dans les sporanges.
- Le sporophyte est la plante 2 n.
- Le gamétophyte est diploïde.
- Les stades gamétophyte et sporophyte sont tous deux multicellulaires.
Les moisissures aquatiques, les oomycètes (« champignons des œufs »), ont été nommées ainsi en raison de leur morphologie semblable à celle des champignons, mais des données moléculaires ont montré que les moisissures aquatiques ne sont pas étroitement liées aux champignons. Les oomycètes sont caractérisés par une paroi cellulaire à base de cellulose et un vaste réseau de filaments qui permettent l'absorption des nutriments. En tant que spores diploïdes, de nombreux oomycètes possèdent deux flagelles orientés de manière opposée (l'un poilu et l'autre lisse) pour la locomotion. Les oomycètes ne sont pas photosynthétiques et comprennent de nombreux sasondes et parasites. Les sasondes apparaissent sous forme d'excroissances duveteuses blanches sur les organismes morts (Figure\(\PageIndex{12}\)). La plupart des oomycètes sont aquatiques, mais certains parasitent les plantes terrestres. L'un des agents pathogènes des plantes est le Phytophthora infestans, l'agent responsable du mildiou de la pomme de terre, comme cela s'est produit lors de la famine de la pomme de terre en Irlande au XIXe siècle.

Rhizaria
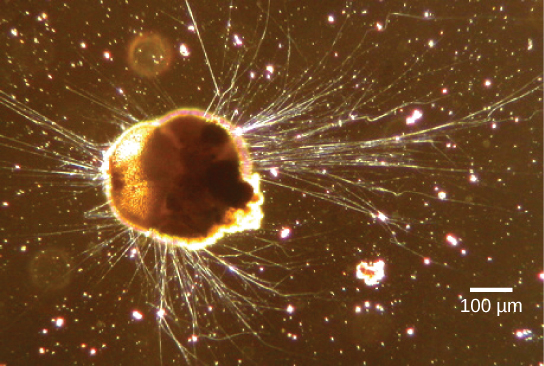
Le supergroupe des Rhizaria comprend de nombreuses amibes, dont la plupart possèdent des pseudopodes filiformes ou aiguilles (Figure\(\PageIndex{13}\)). Les pseudopodes ont pour fonction de piéger et d'engloutir les particules alimentaires et de diriger le mouvement des protistes rhizaires. Ces pseudopodes se projettent vers l'extérieur depuis n'importe quel endroit de la surface cellulaire et peuvent s'ancrer sur un substrat. Le protiste transporte ensuite son cytoplasme dans le pseudopode, déplaçant ainsi toute la cellule. Ce type de mouvement, appelé flux cytoplasmique, est utilisé par plusieurs groupes de protistes comme moyen de locomotion ou comme méthode de distribution des nutriments et de l'oxygène.
Lien vers l'apprentissage
Regardez cette vidéo pour voir le flux cytoplasmique dans une algue verte.
Forums
Les foraminifères, ou forams, sont des protistes hétérotrophes unicellulaires, mesurant entre environ 20 micromètres et plusieurs centimètres de long et ressemblant parfois à de minuscules escargots (Figure\(\PageIndex{14}\)). En tant que groupe, les forams présentent des coques poreuses, appelées tests, qui sont fabriqués à partir de divers matériaux organiques et généralement durcis avec du carbonate de calcium. Les tests peuvent contenir des algues photosynthétiques, que les forams peuvent récolter pour se nourrir. Les pseudopodes des forams s'étendent à travers les pores et permettent aux forams de se déplacer, de se nourrir et de recueillir des matériaux de construction supplémentaires. En général, les forams sont associés au sable ou à d'autres particules dans les habitats marins ou d'eau douce. Les foraminifères sont également utiles comme indicateurs de la pollution et de l'évolution des régimes météorologiques mondiaux.

Radiolaires
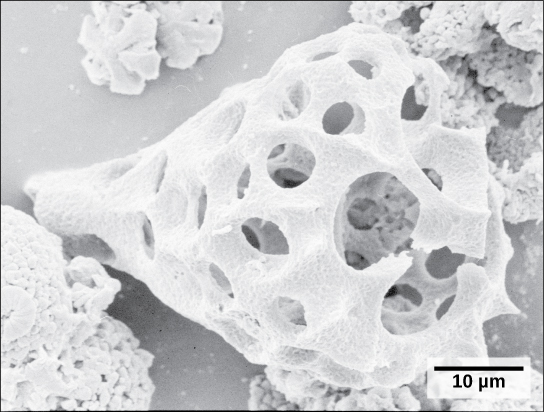
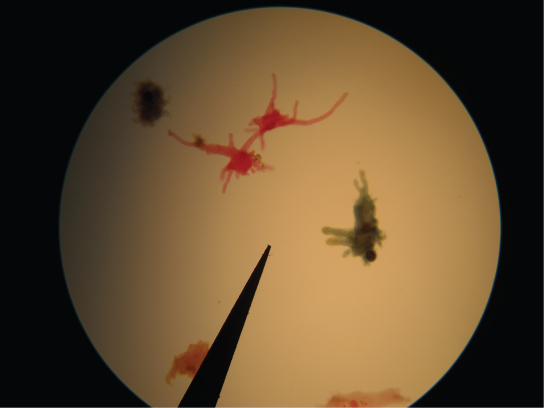
Un deuxième sous-type de Rhizaria, les radiolaires, présentent des surfaces extérieures complexes de silice vitreuse présentant une symétrie radiale ou bilatérale (Figure\(\PageIndex{15}\)). Des pseudopodes en forme d'aiguilles soutenus par des microtubules rayonnent vers l'extérieur à partir du corps cellulaire de ces protistes et ont pour fonction de capter les particules alimentaires. Les coquilles des radiolaires morts s'enfoncent au fond de l'océan, où elles peuvent s'accumuler à une profondeur de 100 mètres. Les radiolaires conservés et sédimentés sont très courants dans les archives fossiles.
Archéplastides
Les algues rouges et les algues vertes font partie du supergroupe Archaeplastida. C'est à partir d'un ancêtre commun de ces protistes que les plantes terrestres ont évolué, puisque leurs plus proches parents se trouvent dans ce groupe. Des preuves moléculaires soutiennent que tous les Archaeplastides sont les descendants d'une relation endosymbiotique entre un protiste hétérotrophe et une cyanobactérie. Les algues rouges et vertes comprennent des formes unicellulaires, multicellulaires et coloniales.
Algues rouges
Les algues rouges, ou rhodophytes, sont principalement multicellulaires, dépourvues de flagelles et varient en taille, allant de protistes unicellulaires microscopiques à de grandes formes multicellulaires regroupées dans la catégorie informelle des algues marines. Le cycle de vie des algues rouges est une alternance de générations. Certaines espèces d'algues rouges contiennent des phycoérythrines, des pigments accessoires photosynthétiques de couleur rouge qui surpassent la teinte verte de la chlorophylle, donnant à ces espèces une apparence de différentes nuances de rouge. D'autres protistes classés comme algues rouges sont dépourvus de phycoérythrines et sont des parasites. Les algues rouges sont communes dans les eaux tropicales où elles ont été détectées à 260 mètres de profondeur. D'autres algues rouges existent dans les milieux terrestres ou d'eau douce.
Algues vertes : chlorophytes et charophytes
Le groupe d'algues le plus abondant est celui des algues vertes. Les algues vertes présentent des caractéristiques similaires à celles des plantes terrestres, notamment en termes de structure chloroplastique. Le fait que ce groupe de protistes ait partagé un ancêtre commun relativement récent avec des plantes terrestres est bien étayé. Les algues vertes se subdivisent en chlorophytes et en charophytes. Les charophytes sont les plus proches parents vivants des plantes terrestres et leur ressemblent en termes de morphologie et de stratégies de reproduction. Les charophytes sont courants dans les milieux humides et leur présence indique souvent la santé de l'écosystème.
Les chlorophytes présentent une grande diversité de formes et de fonctions. Les chlorophytes vivent principalement dans les sols d'eau douce et humides et constituent une composante courante du plancton. Chlamydomonas est un chlorophyte unicellulaire simple présentant une morphologie en forme de poire et deux flagelles antérieurs opposés qui guident ce protiste vers la lumière captée par son œil. Les espèces de chlorophytes plus complexes présentent des gamètes et des spores haploïdes qui ressemblent à ceux de Chlamydomonas.
Le chlorophyte Volvox est l'un des rares exemples d'organisme colonial qui se comporte d'une certaine manière comme un ensemble de cellules individuelles, mais d'autres manières comme les cellules spécialisées d'un organisme multicellulaire (Figure\(\PageIndex{16}\)). Les colonies de Volvox contiennent de 500 à 60 000 cellules, chacune dotée de deux flagelles, contenues dans une matrice sphérique creuse composée d'une sécrétion gélatineuse de glycoprotéines. Les cellules individuelles de Volvox se déplacent de manière coordonnée et sont interconnectées par des ponts cytoplasmiques. Seules quelques cellules se reproduisent pour créer des colonies filles, un exemple de spécialisation cellulaire fondamentale de cet organisme.
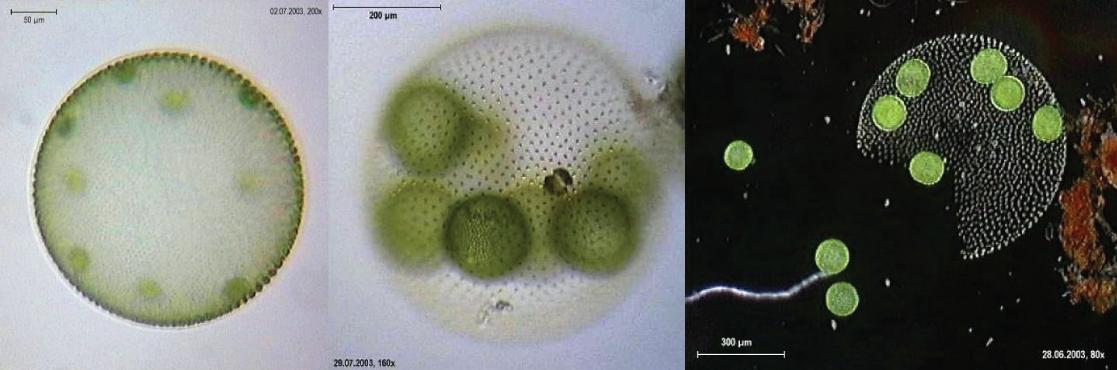
De véritables organismes multicellulaires, tels que la laitue de mer Ulva, sont représentés parmi les chlorophytes. De plus, certains chlorophytes existent sous la forme de grandes cellules uniques multinucléées. Les espèces du genre Caulerpa présentent un feuillage aplati semblable à une fougère et peuvent atteindre une longueur de 3 mètres (Figure\(\PageIndex{17}\)). Les espèces de Caulerpa subissent une division nucléaire, mais leurs cellules ne terminent pas la cytokinèse et restent au contraire des cellules uniques massives et élaborées.

Amibozoaires
Les amibozoaires présentent de façon caractéristique des pseudopodes qui s'étendent comme des tubes ou des lobes plats, plutôt que les pseudopodes ressemblant à des poils des amibes rhizaires (Figure\(\PageIndex{18}\)). Les amibozoaires comprennent plusieurs groupes d'organismes unicellulaires ressemblant à des amibes qui vivent en liberté ou qui sont des parasites.
Moules à slime
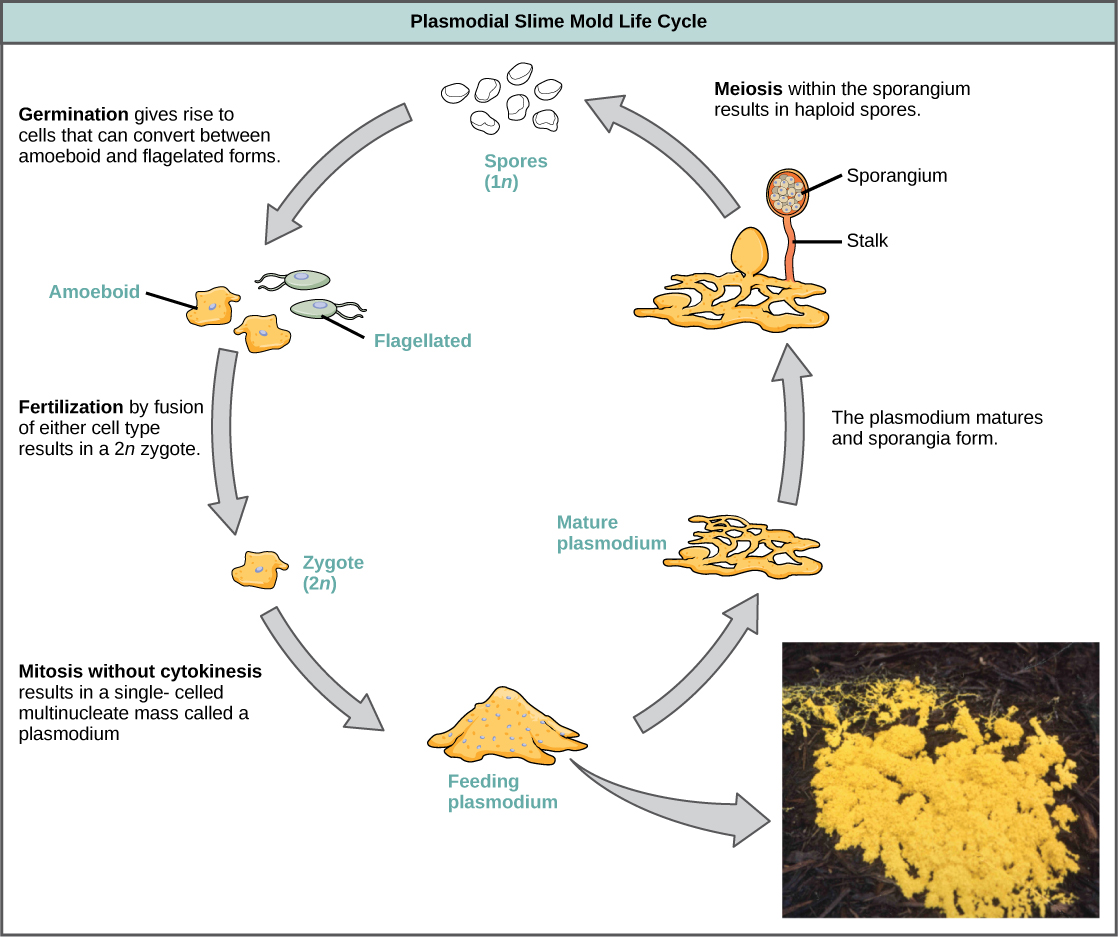
Un sous-ensemble des amibozoaires, les moisissures visqueuses, présente plusieurs similitudes morphologiques avec des champignons que l'on pense être le résultat d'une évolution convergente. Par exemple, en période de stress, certaines moisissures visqueuses se transforment en organes de fructification produisant des spores, un peu comme les champignons.
Les moisissures visqueuses sont classées en fonction de leur cycle de vie en types plasmodiaux ou cellulaires. Les moisissures plasmodiales sont composées de grandes cellules multinucléées et se déplacent le long des surfaces comme une goutte amorphe de boue pendant leur phase d'alimentation (Figure\(\PageIndex{19}\)). Les particules alimentaires sont soulevées et englouties dans la moisissure visqueuse pendant qu'elle glisse. À la maturation, le plasmodium prend l'apparence d'un filet et peut former des organes fructifères, ou sporanges, en période de stress. Les spores haploïdes sont produites par méiose à l'intérieur des sporanges, et les spores peuvent être disséminées dans l'air ou l'eau pour éventuellement atterrir dans des environnements plus favorables. Dans ce cas, les spores germent pour former des cellules haploïdes amiboïdes ou flagellées qui peuvent se combiner les unes aux autres et produire une moisissure visqueuse zygotique diploïde qui complète le cycle de vie.
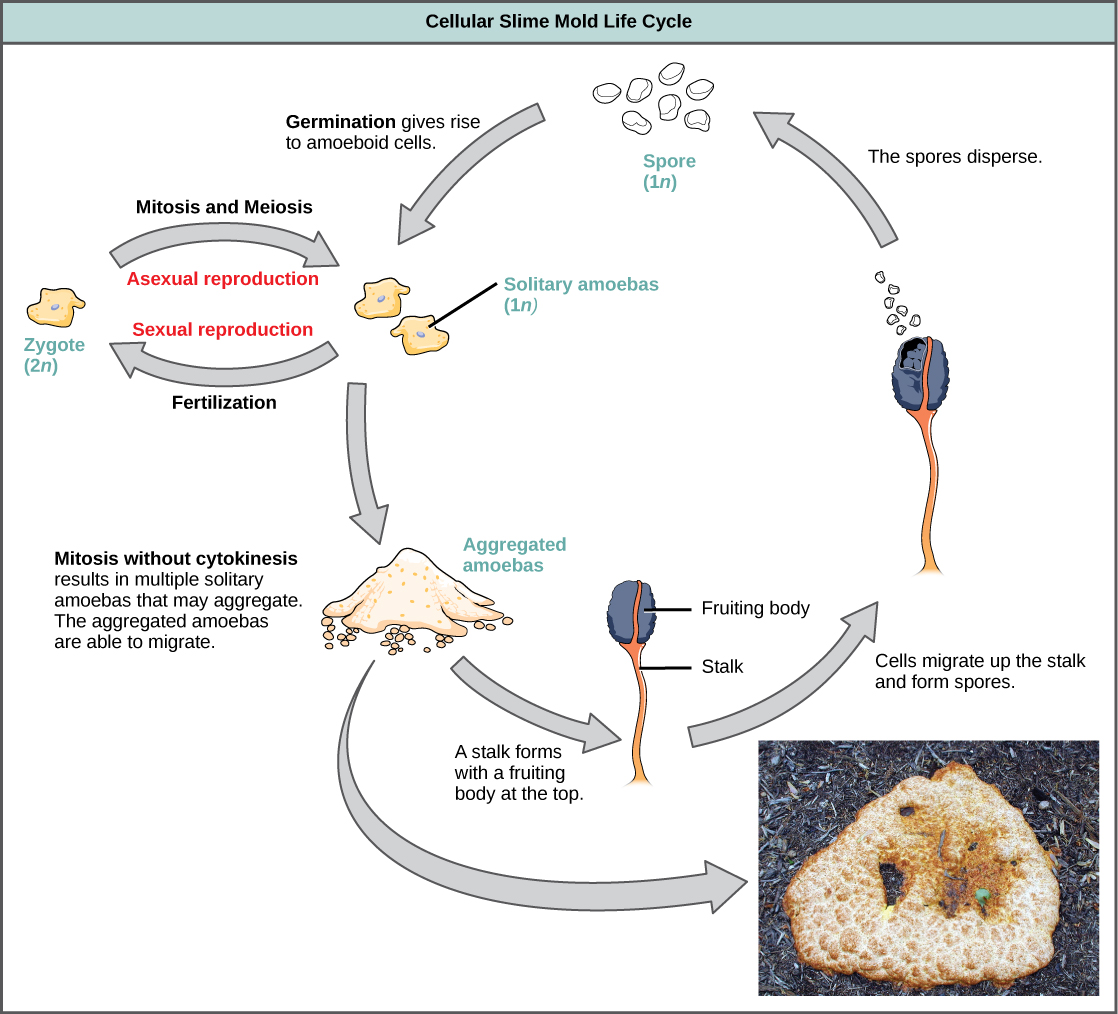
Les moisissures visqueuses cellulaires fonctionnent comme des cellules amiboïdes indépendantes lorsque les nutriments sont abondants (Figure\(\PageIndex{20}\)). Lorsque la nourriture est épuisée, les moisissures visqueuses cellulaires s'empilent les unes sur les autres pour former une masse de cellules qui se comportent comme une seule unité, appelée limace. Certaines cellules de la limace contribuent à former une tige de 2 à 3 millimètres, qui s'assèche et meurt au cours du processus. Les cellules situées au sommet de la tige forment un corps fructifère asexué qui contient des spores haploïdes. Comme pour les moisissures plasmodiales, les spores sont disséminées et peuvent germer si elles atterrissent dans un environnement humide. Un genre représentatif des moisissures visqueuses cellulaires est le Dictyostelium, que l'on trouve couramment dans les sols humides des forêts.
Lien vers l'apprentissage
Consultez ce site pour observer la formation d'un organe fructifère par une moisissure visqueuse cellulaire.
Opisthokonta
Les opisthokonts comprennent les choanoflagellés ressemblant à des animaux, dont on pense qu'ils ressemblent à l'ancêtre commun des éponges et, en fait, à tous les animaux. Les choanoflagellés comprennent des formes unicellulaires et coloniales et comptent environ 244 espèces décrites. Ces organismes présentent un flagelle apical unique entouré d'un collier contractile composé de microvillosités. Le collier utilise un mécanisme similaire à celui des éponges pour filtrer les bactéries destinées à être ingérées par le protiste. La morphologie des choanoflagellés a été reconnue très tôt comme ressemblant aux cellules du col des éponges et suggérant une relation possible avec les animaux.
Les mésomycétozoaires forment un petit groupe de parasites, principalement des poissons, et au moins une forme qui peut parasiter les humains. Leurs cycles de vie sont mal connus. Ces organismes présentent un intérêt particulier, car ils semblent être très proches des animaux. Dans le passé, ils étaient regroupés avec des champignons et d'autres protistes en fonction de leur morphologie.
Résumé
Le processus de classification des protistes en groupes significatifs se poursuit, mais les données génétiques des 20 dernières années ont permis de clarifier de nombreuses relations qui n'étaient pas claires ou erronées auparavant. L'opinion majoritaire à l'heure actuelle est de classer tous les eucaryotes en six supergroupes : Excavata, Chromalveolata, Rhizaria, Archaeplastida, Amoebozoa et Opisthokonta. L'objectif de ce système de classification est de créer des groupes d'espèces qui sont toutes issues d'un ancêtre commun. À l'heure actuelle, la monophylie de certains supergroupes est mieux étayée par des données génétiques que d'autres. Bien qu'il existe d'énormes variations au sein des supergroupes, des points communs aux niveaux morphologique, physiologique et écologique peuvent être identifiés.
Connexions artistiques
Figure\(\PageIndex{8}\) : Laquelle des affirmations suivantes concernant la reproduction sexuée de Paramecium est fausse ?
- Les macronoyaux sont dérivés de micronoyaux.
- La mitose et la méiose se produisent pendant la reproduction sexuée.
- La paire conjuguée échange des macronuclei.
- Chaque parent produit quatre cellules filles.
- Réponse
-
C
Figure\(\PageIndex{11}\) : Parmi les affirmations suivantes concernant le cycle de vie des laminaires, laquelle est fausse ?
- 1 n zoospores se forment dans les sporanges.
- Le sporophyte est la plante 2 n.
- Le gamétophyte est diploïde.
- Les stades gamétophyte et sporophyte sont tous deux multicellulaires.
- Réponse
-
C
Lexique
- pompe à charbon biologique
- processus par lequel le carbone inorganique est fixé par des espèces photosynthétiques qui meurent et tombent ensuite sur le fond marin, où les sasondes ne peuvent pas les atteindre et où leur consommation de dioxyde de carbone ne peut pas être renvoyée dans l'atmosphère
- bioluminescence
- génération et émission de lumière par un organisme, comme dans les dinoflagellés
- vacuole contractile
- vésicule qui se remplit d'eau (lorsqu'elle entre dans la cellule par osmose) puis se contracte pour en extraire l'eau ; vésicule osmorégulatrice
- streaming cytoplasmique
- mouvement du cytoplasme dans un pseudopode étendu de telle sorte que la cellule entière soit transportée vers le site du pseudopode
- hydrogénosome
- organite transporté par les parabasalides (Excavata) qui fonctionne de manière anaérobie et produit de l'hydrogène comme sous-produit ; probablement issu des mitochondries
- kinétoplast
- masse d'ADN contenue dans l'unique mitochondrie surdimensionnée, caractéristique des kinétoplastes (phylum : Euglenozoa)
- mitosome
- organite non fonctionnel présent dans les cellules des diplomonades (Excavata) qui ont probablement évolué à partir d'une mitochondrie
- plancton
- groupe diversifié d'organismes principalement microscopiques qui dérivent dans les systèmes marins et d'eau douce et servent de source de nourriture aux grands organismes aquatiques
- raphé
- fente dans la coque de silice des diatomées à travers laquelle le protiste sécrète un flux de mucopolysaccharides pour la locomotion et la fixation aux substrats
- test
- enveloppe poreuse d'un foram fabriqué à partir de divers matériaux organiques et généralement durci avec du carbonate de calcium