
22.2 : Structure des procaryotes
- Page ID
- 189946

Compétences à développer
- Décrire la structure de base d'un procaryote typique
- Décrire les différences structurelles importantes entre les archées et les bactéries
Il existe de nombreuses différences entre les cellules procaryotes et les cellules eucaryotes. Cependant, toutes les cellules ont quatre structures communes : la membrane plasmique, qui agit comme une barrière pour la cellule et la sépare de son environnement ; le cytoplasme, une substance gélatineuse à l'intérieur de la cellule ; les acides nucléiques, le matériel génétique de la cellule ; et les ribosomes, où se produit la synthèse des protéines. Les procaryotes se présentent sous différentes formes, mais beaucoup se répartissent en trois catégories : les cocci (sphériques), les bacilles (en forme de bâtonnet) et les spirilles (en forme de spirale) (Figure\(\PageIndex{1}\)).

La cellule procaryote
Rappelons que les procaryotes (Figure\(\PageIndex{2}\)) sont des organismes unicellulaires dépourvus d'organites ou d'autres structures liées à la membrane interne. Ils n'ont donc pas de noyau, mais possèdent généralement un seul chromosome, un morceau d'ADN double brin circulaire situé dans une zone de la cellule appelée nucléoïde. La plupart des procaryotes ont une paroi cellulaire à l'extérieur de la membrane plasmique.
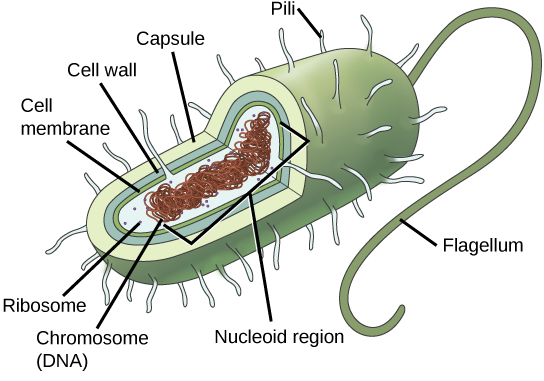
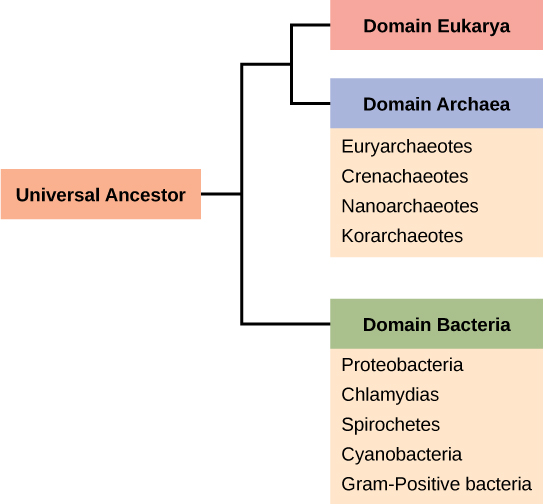
Rappelons que les procaryotes sont divisés en deux domaines différents, les bactéries et les archées, qui, avec les Eucaryas, constituent les trois domaines de la vie (Figure\(\PageIndex{3}\)).
La composition de la paroi cellulaire diffère significativement entre les domaines Bactéries et Archées. La composition de leurs parois cellulaires diffère également de celle des parois cellulaires eucaryotes que l'on trouve chez les plantes (cellulose) ou chez les champignons et les insectes (chitine). La paroi cellulaire fonctionne comme une couche protectrice et est responsable de la forme de l'organisme. Certaines bactéries ont une capsule externe à l'extérieur de la paroi cellulaire. D'autres structures sont présentes chez certaines espèces procaryotes, mais pas chez d'autres (Tableau\(\PageIndex{1}\)). Par exemple, la capsule présente chez certaines espèces permet à l'organisme de se fixer aux surfaces, le protège de la déshydratation et des attaques des cellules phagocytaires, et rend les agents pathogènes plus résistants à nos réponses immunitaires. Certaines espèces possèdent également des flagelles (singulier, flagelle) utilisés pour la locomotion et des pili (singulier, pilus) utilisés pour la fixation aux surfaces. Les plasmides, qui sont constitués d'ADN extra-chromosomique, sont également présents chez de nombreuses espèces de bactéries et d'archées.
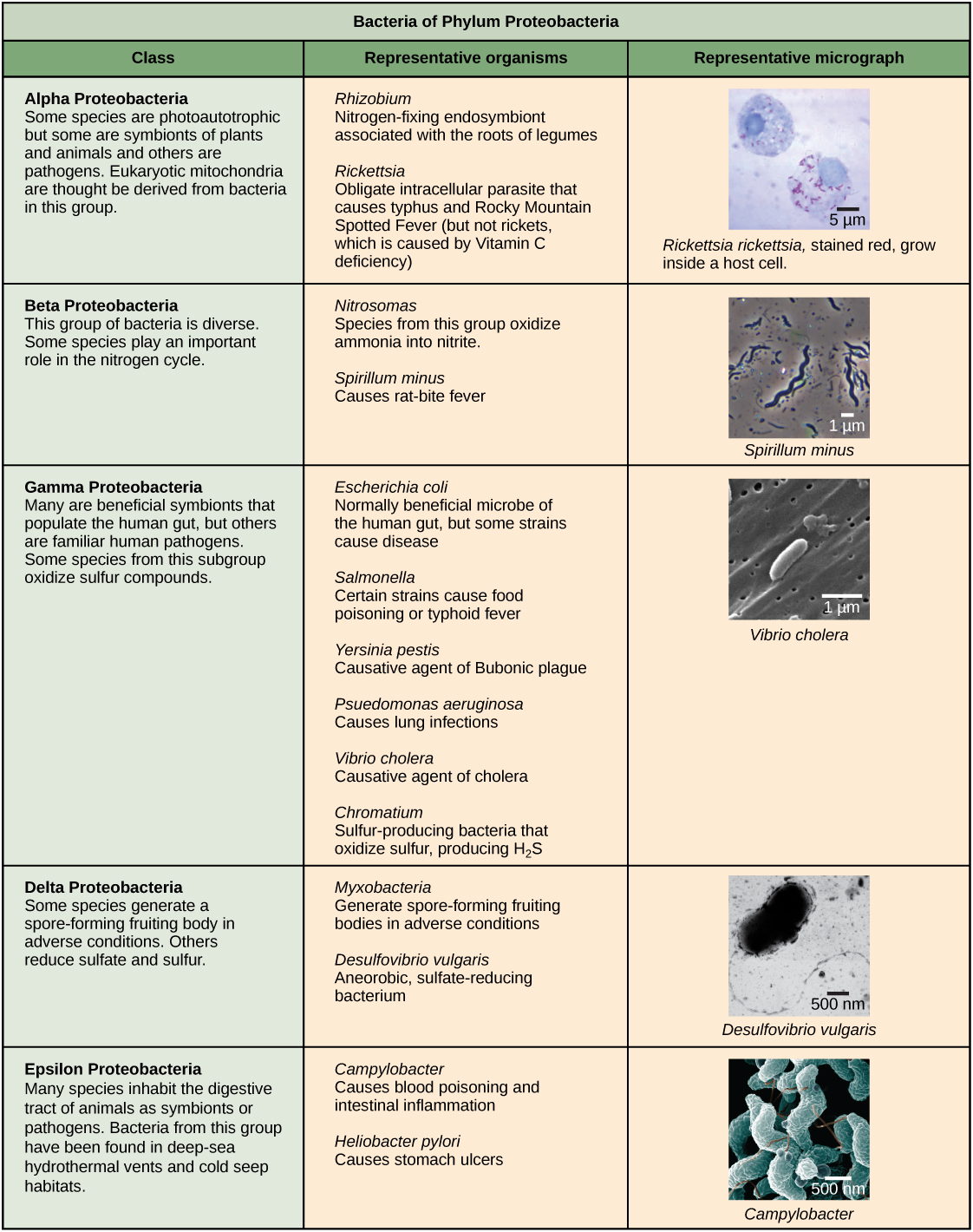
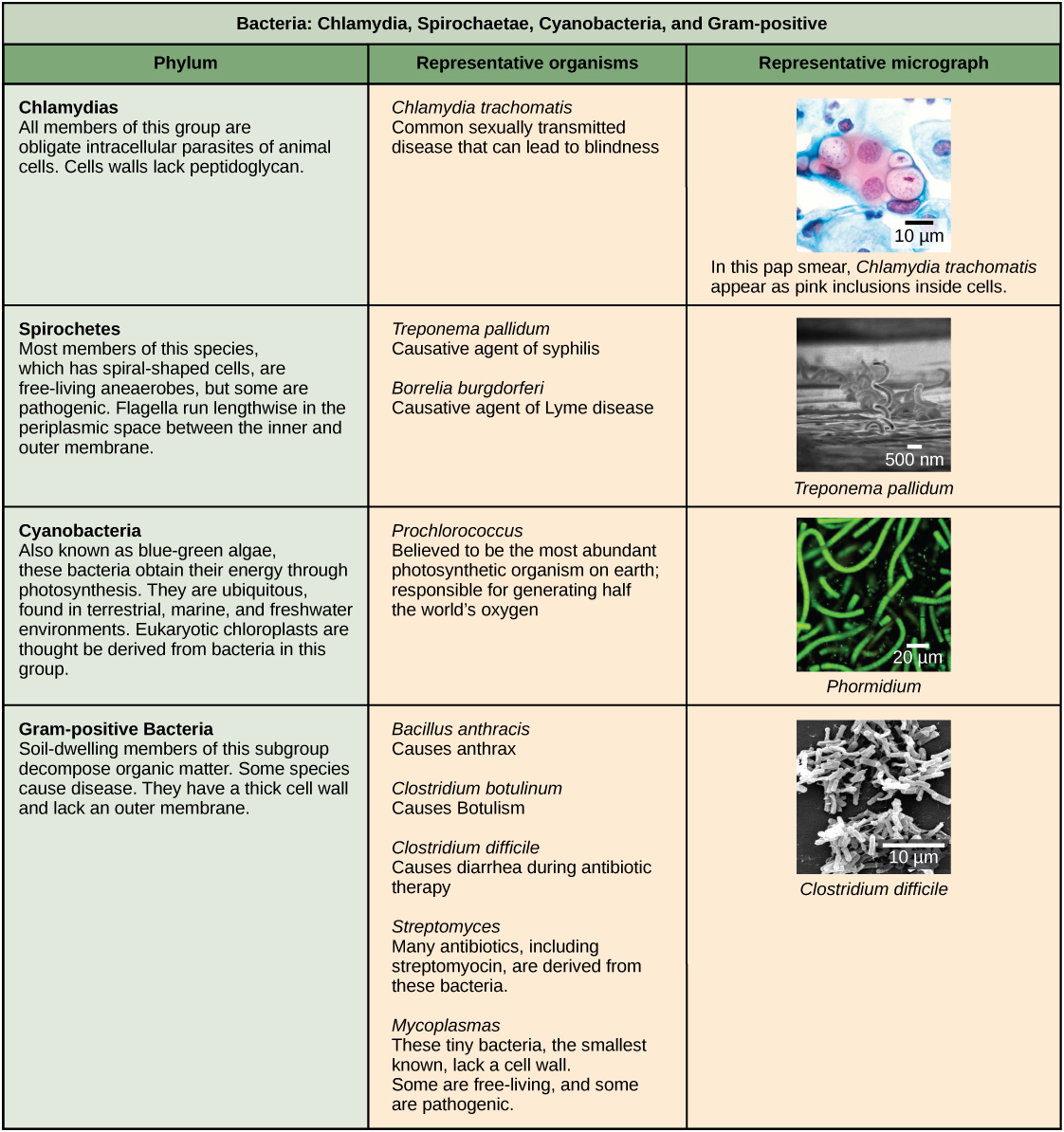
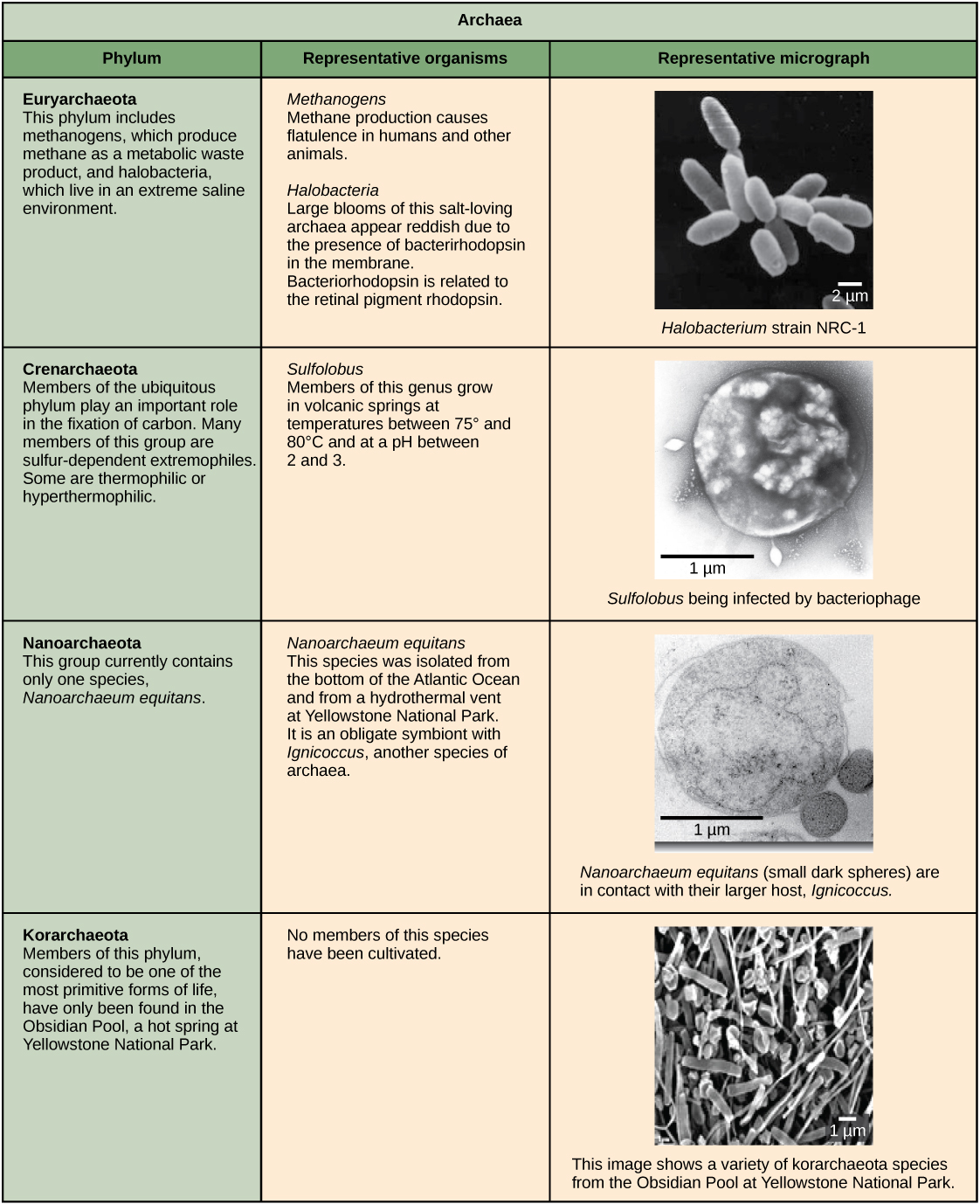
Les caractéristiques du phyla des bactéries sont décrites dans la figure\(\PageIndex{4}\) et la figure\(\PageIndex{5}\) ; les archées sont décrites dans la figure\(\PageIndex{6}\).
La membrane plasmique
La membrane plasmique est une mince bicouche lipidique (6 à 8 nanomètres) qui entoure complètement la cellule et sépare l'intérieur de l'extérieur. Sa nature sélectivement perméable maintient les ions, les protéines et les autres molécules à l'intérieur de la cellule et les empêche de se diffuser dans l'environnement extracellulaire, tandis que d'autres molécules peuvent se déplacer à travers la membrane. Rappelons que la structure générale d'une membrane cellulaire est une bicouche de phospholipides composée de deux couches de molécules lipidiques. Dans les membranes des cellules archéales, les chaînes d'isoprène (phytanyle) liées au glycérol remplacent les acides gras liés au glycérol dans les membranes bactériennes. Certaines membranes archéologiques sont des monocouches lipidiques au lieu de bicouches (Figure\(\PageIndex{7}\)).
La paroi cellulaire
Le cytoplasme des cellules procaryotes présente une forte concentration de solutés dissous. Par conséquent, la pression osmotique à l'intérieur de la cellule est relativement élevée. La paroi cellulaire est une couche protectrice qui entoure certaines cellules et leur donne forme et rigidité. Il est situé à l'extérieur de la membrane cellulaire et empêche la lyse osmotique (éclatement dû à l'augmentation du volume). La composition chimique des parois cellulaires varie entre les archées et les bactéries, ainsi qu'entre les espèces bactériennes.
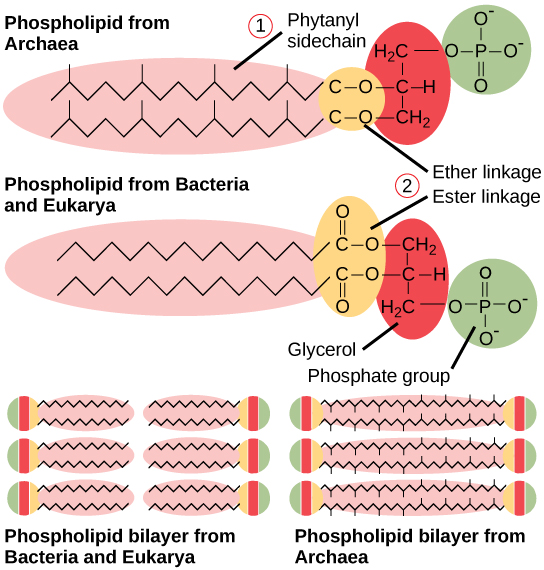
Les parois cellulaires bactériennes contiennent du peptidoglycane, composé de chaînes de polysaccharides réticulées par des peptides inhabituels contenant à la fois des acides aminés L et D, notamment de l'acide D-glutamique et de la D-alanine. Les protéines ne contiennent normalement que des acides aminés L. Par conséquent, bon nombre de nos antibiotiques agissent en imitant les acides aminés D et ont donc des effets spécifiques sur le développement de la paroi cellulaire bactérienne. Il existe plus de 100 formes différentes de peptidoglycane. Les protéines de la couche S (couche superficielle) sont également présentes à l'extérieur des parois cellulaires des archées et des bactéries.
Les bactéries sont divisées en deux grands groupes : Gram positif et Gram négatif, en fonction de leur réaction à la coloration à Gram. Notez que toutes les bactéries à Gram positif appartiennent à un phylum ; les bactéries de l'autre phyla (protéobactéries, chlamydias, spirochètes, cyanobactéries et autres) sont à Gram négatif. La méthode de coloration Gram doit son nom à son inventeur, le scientifique danois Hans Christian Gram (1853-1938). Les différentes réponses bactériennes à la procédure de coloration sont finalement dues à la structure de la paroi cellulaire. Les organismes à Gram positif n'ont généralement pas la membrane externe des organismes à Gram négatif (Figure\(\PageIndex{8}\)). Jusqu'à 90 pour cent de la paroi cellulaire des bactéries à Gram positif est composée de peptidoglycane, et la majeure partie du reste est composée de substances acides appelées acides teichoïques. Les acides teichoïques peuvent être liés par covalence aux lipides de la membrane plasmique pour former des acides lipotéichoïques. Les acides lipotéichoïques ancrent la paroi cellulaire à la membrane cellulaire. Les bactéries à Gram négatif possèdent une paroi cellulaire relativement mince composée de quelques couches de peptidoglycane (seulement 10 % de la paroi cellulaire totale), entourée d'une enveloppe extérieure contenant des lipopolysaccharides (LPS) et des lipoprotéines. Cette enveloppe extérieure est parfois appelée deuxième bicouche lipidique. La chimie de cette enveloppe externe est cependant très différente de celle de la bicouche lipidique typique qui forme les membranes plasmiques.
Art Connection
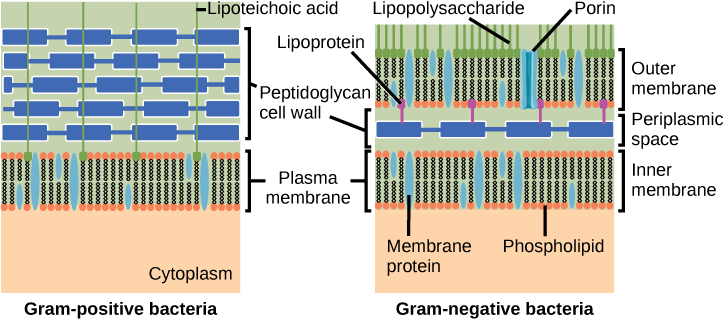
Laquelle des affirmations suivantes est vraie ?
- Les bactéries à Gram positif ont une seule paroi cellulaire ancrée à la membrane cellulaire par l'acide lipotéichoïque.
- Les porines permettent l'entrée de substances dans les bactéries Gram-positives et Gram-négatives.
- La paroi cellulaire des bactéries à Gram négatif est épaisse et la paroi cellulaire des bactéries à Gram positif est mince.
- Les bactéries à Gram négatif ont une paroi cellulaire constituée de peptidoglycane, tandis que les bactéries à Gram positif ont une paroi cellulaire constituée d'acide lipotéichoïque.
Les parois cellulaires archéennes ne contiennent pas de peptidoglycane. Il existe quatre types différents de parois cellulaires archéennes. L'un des types est composé de pseudopeptidoglycane, dont la morphologie est similaire au peptidoglycane mais qui contient différents sucres dans la chaîne polysaccharidique. Les trois autres types de parois cellulaires sont composés de polysaccharides, de glycoprotéines ou de protéines pures.
| Caractéristique structurelle | bactéries | Archaea |
|---|---|---|
| Type de cellule | Procaryote | Procaryote |
| Morphologie cellulaire | Variable | Variable |
| Paroi cellulaire | Contient du peptidoglycane | Ne contient pas de peptidoglycane |
| Type de membrane cellulaire | bicouche lipidique | Bicouche lipidique ou monocouche lipidique |
| Lipides des membranes plasmiques | Acides gras | Groupes phytanyles |
Reproduction
La reproduction chez les procaryotes est asexuée et se fait généralement par fission binaire. Rappelons que l'ADN d'un procaryote existe sous la forme d'un chromosome circulaire unique. Les procaryotes ne subissent pas de mitose. Le chromosome est plutôt répliqué et les deux copies obtenues se séparent l'une de l'autre, en raison de la croissance de la cellule. Le procaryote, maintenant agrandi, est pincé vers l'intérieur au niveau de son équateur et les deux cellules qui en résultent, qui sont des clones, se séparent. La fission binaire ne permet pas de recombinaison génétique ou de diversité génétique, mais les procaryotes peuvent partager des gènes par trois autres mécanismes.
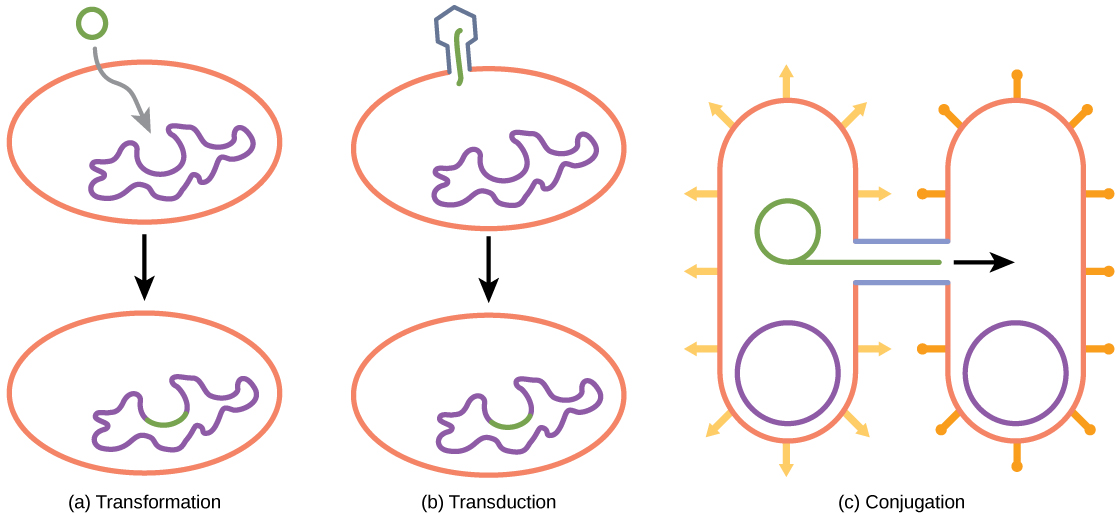
Lors de sa transformation, le procaryote absorbe l'ADN présent dans son environnement qui est excrété par d'autres procaryotes. Si une bactérie non pathogène absorbe l'ADN d'un gène de toxine provenant d'un pathogène et incorpore le nouvel ADN dans son propre chromosome, elle peut également devenir pathogène. Lors de la transduction, les bactériophages, les virus qui infectent les bactéries, transportent parfois de courts fragments d'ADN chromosomique d'une bactérie à l'autre. La transduction donne naissance à un organisme recombinant. Les archées ne sont pas affectées par les bactériophages, mais possèdent leurs propres virus qui transloquent le matériel génétique d'un individu à un autre. Lors de la conjugaison, l'ADN est transféré d'un procaryote à un autre au moyen d'un pilus, qui met les organismes en contact les uns avec les autres. L'ADN transféré peut se présenter sous la forme d'un plasmide ou d'un hybride, contenant à la fois un plasmide et un ADN chromosomique. Ces trois processus d'échange d'ADN sont illustrés sur la figure\(\PageIndex{9}\).
La reproduction peut être très rapide : quelques minutes pour certaines espèces. Cette courte durée de génération, associée à des mécanismes de recombinaison génétique et à des taux élevés de mutation, entraîne l'évolution rapide des procaryotes, leur permettant de répondre très rapidement aux changements environnementaux (tels que l'introduction d'un antibiotique).
Evolution Connection : L'évolution des procaryotes
Comment les scientifiques répondent-ils aux questions concernant l'évolution des procaryotes ? Contrairement aux animaux, les artefacts contenus dans les archives fossiles des procaryotes fournissent très peu d'informations. Les fossiles d'anciens procaryotes ressemblent à de minuscules bulles dans la roche. Certains scientifiques se tournent vers la génétique et vers le principe de l'horloge moléculaire, selon lequel plus les deux espèces divergeront récemment, plus leurs gènes (et donc leurs protéines) seront similaires. À l'inverse, les espèces qui ont divergé il y a longtemps auront davantage de gènes différents.
Des scientifiques de l'Institut d'astrobiologie de la NASA et du Laboratoire européen de biologie moléculaire ont collaboré pour analyser l'évolution moléculaire de 32 protéines spécifiques communes à 72 espèces de procaryotes. 1 Le modèle qu'ils ont dérivé de leurs données indique que trois groupes importants de bactéries — les actinobactéries, les déinocoques et les cyanobactéries (que les auteurs appellent les terrabactéries) — ont été les premiers à coloniser les terres. (Rappelons que Deinococcus est un genre de procaryote, une bactérie, hautement résistant aux rayonnements ionisants.) Les cyanobactéries sont des photosynthétiseurs, tandis que les actinobactéries sont un groupe de bactéries très communes qui incluent des espèces importantes dans la décomposition des déchets organiques.
Les chronologies de divergence suggèrent que les bactéries (membres du domaine Bactéries) ont divergé des espèces ancestrales communes il y a 2,5 à 3,2 milliards d'années, alors que les archées ont divergé plus tôt : entre 3,1 et 4,1 milliards d'années. Eukarya a ensuite divergé de la ligne archéenne. Les travaux suggèrent en outre que les stromatolites qui se sont formés avant l'apparition des cyanobactéries (il y a environ 2,6 milliards d'années) se sont photosynthétisés dans un environnement anoxique et qu'en raison des modifications des terrabactéries pour le sol (résistance au dessèchement et possession de composés qui protègent l'organisme) à cause de l'excès de lumière), la photosynthèse à l'aide d'oxygène peut être étroitement liée aux adaptations nécessaires à la survie sur terre.
Résumé
Les procaryotes (domaines Archaea et Bactéries) sont des organismes unicellulaires dépourvus de noyau. Ils ont un seul morceau d'ADN circulaire dans la zone nucléoïde de la cellule. La plupart des procaryotes ont une paroi cellulaire qui se trouve à l'extérieur de la limite de la membrane plasmique. Certains procaryotes peuvent avoir des structures supplémentaires telles qu'une capsule, des flagelles et des pili. Les bactéries et les archées diffèrent par la composition lipidique de leurs membranes cellulaires et par les caractéristiques de la paroi cellulaire. Dans les membranes archéales, les unités phytanyles, plutôt que les acides gras, sont liées au glycérol. Certaines membranes archéologiques sont des monocouches lipidiques au lieu de bicouches.
La paroi cellulaire est située à l'extérieur de la membrane cellulaire et empêche la lyse osmotique. La composition chimique des parois cellulaires varie selon les espèces. Les parois cellulaires bactériennes contiennent du peptidoglycane. Les parois cellulaires archéennes ne contiennent pas de peptidoglycane, mais elles peuvent contenir du pseudopeptidoglycane, des polysaccharides, des glycoprotéines ou des parois cellulaires à base de protéines. Les bactéries peuvent être divisées en deux grands groupes : Gram positif et Gram négatif, en fonction de la réaction de coloration de Gram. Les organismes à Gram positif ont une paroi cellulaire épaisse, ainsi que des acides teichoïques. Les organismes à Gram négatif ont une paroi cellulaire mince et une enveloppe externe contenant des lipopolysaccharides et des lipoprotéines.
Connexions artistiques
Figure\(\PageIndex{8}\) : Lequel des énoncés suivants est vrai ?
- Les bactéries à Gram positif ont une seule paroi cellulaire ancrée à la membrane cellulaire par l'acide lipotéichoïque.
- Les porines permettent l'entrée de substances dans les bactéries Gram-positives et Gram-négatives.
- La paroi cellulaire des bactéries à Gram négatif est épaisse et la paroi cellulaire des bactéries à Gram positif est mince.
- Les bactéries à Gram négatif ont une paroi cellulaire constituée de peptidoglycane, tandis que les bactéries à Gram positif ont une paroi cellulaire constituée d'acide lipotéichoïque.
- Réponse
-
UN
Notes
- 1 Battistuzzi, FU, Feijao, A, et Hedges, SB. Une échelle de temps génomique de l'évolution des procaryotes : aperçu de l'origine de la méthanogenèse, de la phototrophie et de la colonisation des terres. BioMed Central : Biologie évolutive 4 (2004) : 44, doi : 10.1186/1471-2148-4-44.
Lexique
- capsule
- structure externe qui permet à un procaryote de se fixer aux surfaces et le protège de la déshydratation
- conjugaison
- processus par lequel les procaryotes transfèrent l'ADN d'un individu à un autre à l'aide d'un pilus
- Gram négatif
- bactérie dont la paroi cellulaire contient peu de peptidoglycane mais possède une membrane externe
- Gram positif
- bactérie contenant principalement du peptidoglycane dans ses parois cellulaires
- peptidoglycane
- matériau composé de chaînes de polysaccharides réticulées à des peptides inhabituels
- pilus
- appendice superficiel de certains procaryotes utilisé pour la fixation à des surfaces, y compris d'autres procaryotes
- pseudopeptidoglycane
- composant des parois cellulaires des archées dont la morphologie est similaire à celle du peptidoglycane mais qui contient des sucres différents
- couche S
- protéine de la couche superficielle présente à l'extérieur des parois cellulaires des archées et des bactéries
- acide teichoïque
- polymère associé à la paroi cellulaire des bactéries à Gram positif
- transduction
- processus par lequel un bactériophage déplace l'ADN d'un procaryote à un autre
- transformation
- processus par lequel un procaryote absorbe l'ADN présent dans son environnement qui est excrété par d'autres procaryotes