
21.1 : Évolution, morphologie et classification des virus
- Page ID
- 189906

Compétences à développer
- Décrire comment les virus ont été découverts pour la première fois et comment ils sont détectés
- Discutez de trois hypothèses concernant l'évolution des virus
- Reconnaître les formes de base des virus
- Comprendre les systèmes de classification anciens et émergents des virus
Les virus sont des entités diverses. Leur structure, leurs méthodes de réplication et leurs hôtes cibles varient. Presque toutes les formes de vie, des bactéries aux archées, en passant par les eucaryotes tels que les plantes, les animaux et les champignons, sont infectées par des virus. Bien que la majeure partie de la diversité biologique puisse être comprise à travers l'histoire de l'évolution, comme la façon dont les espèces se sont adaptées aux conditions et à l'environnement, beaucoup de choses sur l'origine et l'évolution des virus
Découverte et détection
Les virus ont été découverts pour la première fois après la mise au point d'un filtre en porcelaine, appelé filtre Chamberland-Pasteur, qui pouvait éliminer toutes les bactéries visibles au microscope de tout échantillon liquide. En 1886, Adolph Meyer a démontré qu'une maladie des plants de tabac, la maladie de la mosaïque du tabac, pouvait être transmise d'une plante malade à une plante saine par le biais d'extraits liquides de plantes. En 1892, Dmitri Ivanowski a montré que cette maladie pouvait être transmise de cette manière même après que le filtre Chamberland-Pasteur ait éliminé toutes les bactéries viables de l'extrait. Pourtant, il a fallu de nombreuses années avant qu'il soit prouvé que ces agents infectieux « filtrables » n'étaient pas simplement de très petites bactéries, mais qu'il s'agissait d'un nouveau type de très petite particule causant des maladies.
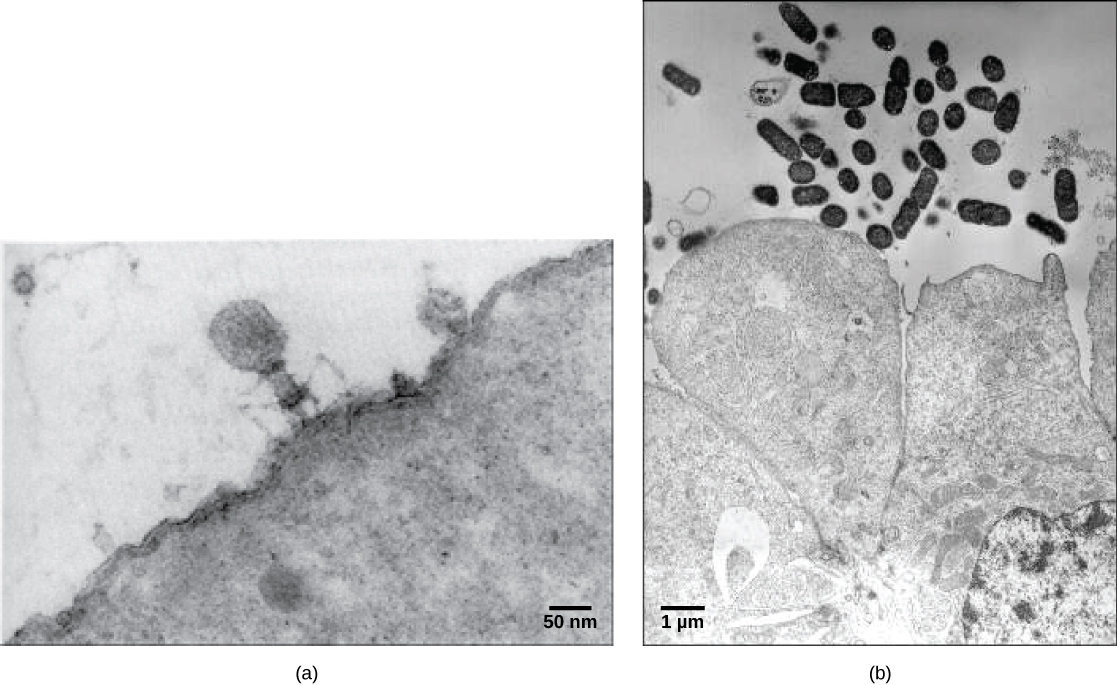
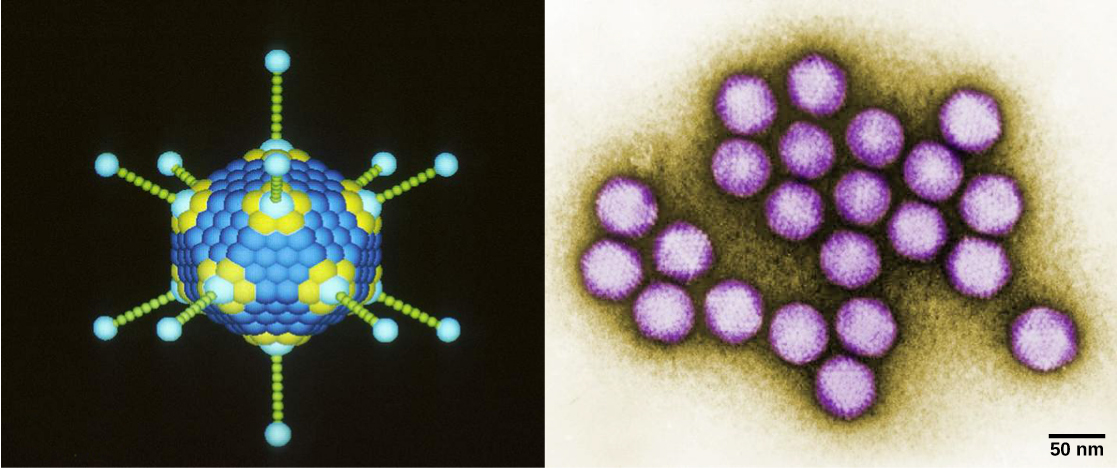
Les virions, des particules virales uniques, sont très petits, d'environ 20 à 250 nanomètres de diamètre. Ces particules virales individuelles constituent la forme infectieuse d'un virus situé à l'extérieur de la cellule hôte. Contrairement aux bactéries (qui sont environ 100 fois plus grosses), nous ne pouvons pas détecter les virus au microscope optique, à l'exception de certains grands virions de la famille des poxvirus. Ce n'est que lors de la mise au point du microscope électronique à la fin des années 1930 que les scientifiques ont eu une première bonne idée de la structure du virus de la mosaïque du tabac (TMV) (Figure\(\PageIndex{1}\)) et d'autres virus (Figure\(\PageIndex{1}\)). La structure de surface des virions peut être observée à la fois par microscopie électronique à balayage et par transmission, tandis que les structures internes du virus ne peuvent être observées que sur des images au microscope électronique à transmission. L'utilisation de ces technologies a permis de découvrir de nombreux virus de tous les types d'organismes vivants. Ils ont d'abord été regroupés selon une morphologie commune. Plus tard, les groupes de virus ont été classés selon le type d'acide nucléique qu'ils contenaient, ADN ou ARN, et selon que leur acide nucléique était simple ou double brin. Plus récemment, l'analyse moléculaire des cycles réplicatifs viraux a permis d'affiner leur classification.
Évolution des virus
Bien que les biologistes aient accumulé une quantité importante de connaissances sur l'évolution des virus actuels, on en sait beaucoup moins sur leur origine. Lorsqu'ils explorent l'histoire évolutive de la plupart des organismes, les scientifiques peuvent examiner les archives fossiles et les preuves historiques similaires. Cependant, les virus ne se fossilisent pas. Les chercheurs doivent donc faire des conjectures en étudiant l'évolution des virus actuels et en utilisant des informations biochimiques et génétiques pour créer des historiques spéculatifs sur les virus.
Bien que la plupart des résultats s'accordent sur le fait que les virus n'ont pas d'ancêtre commun, les chercheurs n'ont pas encore trouvé d'hypothèse sur l'origine des virus qui soit pleinement acceptée sur le terrain. L'une de ces hypothèses, appelée dévolution ou hypothèse régressive, propose d'expliquer l'origine des virus en suggérant que les virus ont évolué à partir de cellules libres. Cependant, de nombreux éléments expliquant comment ce processus a pu se produire restent un mystère. Une deuxième hypothèse (appelée hypothèse d'évasion ou hypothèse progressive) tient compte des virus possédant un génome d'ARN ou d'ADN et suggère que les virus proviennent de molécules d'ARN et d'ADN qui se sont échappées d'une cellule hôte. Une troisième hypothèse suppose l'existence d'un système d'autoréplication similaire à celui d'autres molécules autoréplicatives, évoluant probablement en même temps que les cellules dont elles dépendent en tant qu'hôtes ; des études sur certains agents pathogènes des plantes confirment cette hypothèse.
À mesure que la technologie progresse, les scientifiques peuvent développer et affiner d'autres hypothèses pour expliquer l'origine des virus. Le domaine émergent appelé systématique moléculaire des virus tente d'y parvenir en comparant du matériel génétique séquencé. Ces chercheurs espèrent un jour mieux comprendre l'origine des virus, une découverte qui pourrait mener à des avancées dans les traitements des maladies qu'ils provoquent.
Morphologie virale
Les virus sont acellulaires, c'est-à-dire qu'ils sont des entités biologiques qui n'ont pas de structure cellulaire. Ils sont donc dépourvus de la plupart des composants des cellules, tels que les organites, les ribosomes et la membrane plasmique. Un virion est constitué d'un noyau d'acide nucléique, d'un enrobage protéique externe ou capside, et parfois d'une enveloppe extérieure constituée de membranes protéiques et phospholipidiques dérivées de la cellule hôte. Les virus peuvent également contenir des protéines supplémentaires, telles que des enzymes. La différence la plus évidente entre les membres des familles virales est leur morphologie, qui est très diverse. Une caractéristique intéressante de la complexité virale est que la complexité de l'hôte n'est pas corrélée à la complexité du virion. Certaines des structures viriennes les plus complexes sont observées dans les bactériophages, des virus qui infectent les organismes vivants les plus simples, les bactéries.
Morphologie
Les virus se présentent sous de nombreuses formes et tailles, mais elles sont cohérentes et distinctes pour chaque famille virale. Tous les virions possèdent un génome d'acide nucléique recouvert d'une couche protectrice de protéines, appelée capside. La capside est composée de sous-unités protéiques appelées capsomères. Certaines capsides virales sont de simples « sphères » polyédriques, tandis que d'autres ont une structure assez complexe.
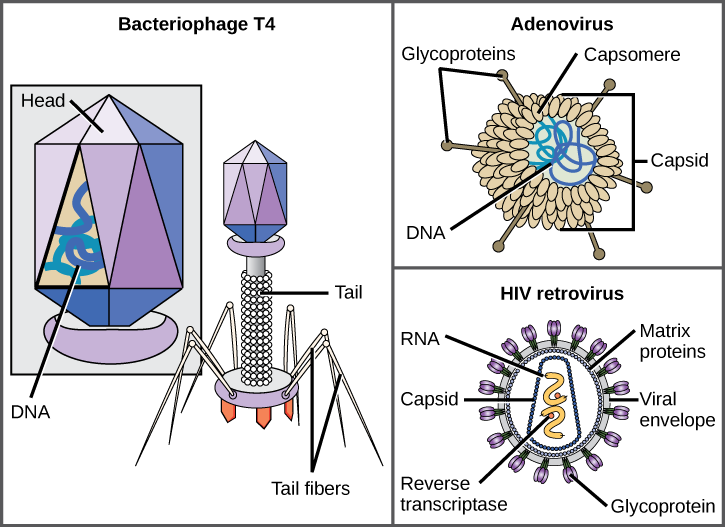
En général, les formes des virus sont classées en quatre groupes : filamenteux, isométrique (ou icosaédrique), enveloppé et tête et queue. Les virus filamenteux sont longs et cylindriques. De nombreux virus végétaux sont filamenteux, y compris le TMV. Les virus isométriques ont des formes à peu près sphériques, comme le poliovirus ou l'herpèsvirus. Les virus enveloppés ont des membranes entourant les capsides. Les virus animaux, tels que le VIH, sont souvent enveloppés. Les virus de la tête et de la queue infectent les bactéries et ont une tête similaire à celle des virus icosaédriques et une forme de queue semblable à celle des virus filamenteux.
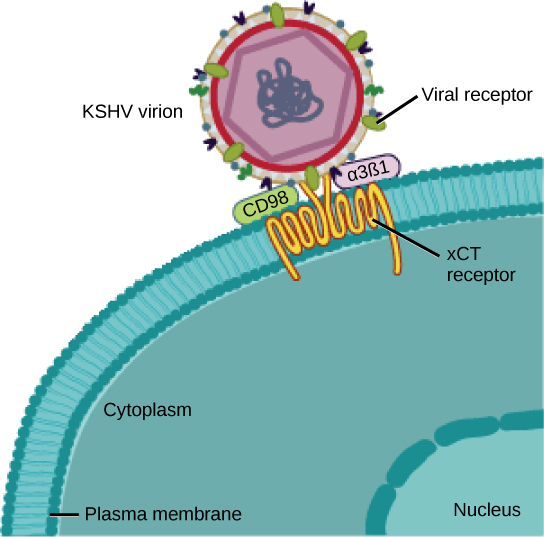
De nombreux virus utilisent une sorte de glycoprotéine pour se fixer à leurs cellules hôtes via des molécules présentes sur la cellule appelées récepteurs viraux (Figure\(\PageIndex{2}\)). Pour ces virus, l'attachement est nécessaire pour pénétrer ultérieurement dans la membrane cellulaire, afin qu'ils puissent terminer leur réplication à l'intérieur de la cellule. Les récepteurs utilisés par les virus sont des molécules qui se trouvent normalement à la surface des cellules et qui ont leurs propres fonctions physiologiques. Les virus ont simplement évolué pour utiliser ces molécules pour leur propre réplication. Par exemple, le VIH utilise la molécule CD4 sur les lymphocytes T comme l'un de ses récepteurs. Le CD4 est un type de molécule appelé molécule d'adhésion cellulaire, qui a pour fonction de maintenir différents types de cellules immunitaires à proximité les uns des autres pendant la génération d'une réponse immunitaire des lymphocytes T.
Parmi les virions les plus complexes connus, le bactériophage T4, qui infecte la bactérie Escherichia coli, possède une structure de queue que le virus utilise pour se fixer aux cellules hôtes et une structure de tête qui abrite son ADN.
L'adénovirus, un virus animal non enveloppé qui cause des maladies respiratoires chez l'homme, utilise des pointes de glycoprotéines qui dépassent de ses capsomères pour se fixer aux cellules hôtes. Les virus non enveloppés incluent également ceux qui causent la poliomyélite (poliovirus), les verrues plantaires (papillomavirus) et l'hépatite A (virus de l'hépatite A).
Les virions enveloppés tels que le VIH, l'agent causal du sida, sont constitués d'acides nucléiques (ARN dans le cas du VIH) et de protéines de capside entourés d'une enveloppe bicouche phospholipidique et des protéines associées. Les glycoprotéines incorporées dans l'enveloppe virale sont utilisées pour se fixer aux cellules hôtes. Les autres protéines d'enveloppe sont les protéines matricielles qui stabilisent l'enveloppe et jouent souvent un rôle dans l'assemblage des virions de descendance. La varicelle, la grippe et les oreillons sont des exemples de maladies causées par des virus munis d'enveloppes. En raison de la fragilité de l'enveloppe, les virus non enveloppés sont plus résistants aux changements de température, de pH et à certains désinfectants que les virus enveloppés.
Dans l'ensemble, la forme du virion et la présence ou l'absence d'enveloppe nous renseignent peu sur la maladie que le virus peut provoquer ou sur les espèces qu'il peut infecter, mais elles constituent tout de même des moyens utiles pour commencer la classification virale (Figure\(\PageIndex{3}\)).
Exercice\(\PageIndex{1}\)
Parmi les affirmations suivantes concernant la structure du virus, laquelle est vraie ?
- Tous les virus sont enfermés dans une membrane virale.
- Le capsomère est composé de petites sous-unités protéiques appelées capsides.
- L'ADN est le matériel génétique de tous les virus.
- Les glycoprotéines aident le virus à se fixer à la cellule hôte.
- Réponse
-
D
Types d'acides nucléiques
Contrairement à presque tous les organismes vivants qui utilisent l'ADN comme matériel génétique, les virus peuvent utiliser de l'ADN ou de l'ARN comme leur matériel génétique. Le noyau du virus contient le génome ou le contenu génétique total du virus. Les génomes viraux ont tendance à être petits et ne contiennent que les gènes codant pour des protéines que le virus ne peut pas obtenir de la cellule hôte. Ce matériel génétique peut être simple ou double brin. Il peut également être linéaire ou circulaire. Alors que la plupart des virus contiennent un seul acide nucléique, d'autres ont des génomes qui en comportent plusieurs, appelés segments.
Dans les virus à ADN, l'ADN viral dirige les protéines de réplication de la cellule hôte pour synthétiser de nouvelles copies du génome viral et pour transcrire et traduire ce génome en protéines virales. Les virus à ADN provoquent des maladies humaines, telles que la varicelle, l'hépatite B et certaines maladies vénériennes, comme l'herpès et les verrues génitales.
Les virus à ARN contiennent uniquement de l'ARN comme matériel génétique. Pour répliquer leur génome dans la cellule hôte, les virus à ARN codent des enzymes capables de répliquer l'ARN dans l'ADN, ce qui ne peut pas être fait par la cellule hôte. Ces enzymes à ARN polymérase sont plus susceptibles de faire des erreurs de copie que les ADN polymérases et commettent donc souvent des erreurs lors de la transcription. Pour cette raison, les mutations dans les virus à ARN sont plus fréquentes que dans les virus à ADN. Cela les amène à changer et à s'adapter plus rapidement à leur hôte. Les maladies humaines causées par les virus à ARN incluent l'hépatite C, la rougeole et la rage.
Classification des virus
Pour comprendre les caractéristiques communes aux différents groupes de virus, un système de classification est nécessaire. Comme on ne pense pas que la plupart des virus aient évolué à partir d'un ancêtre commun, les méthodes utilisées par les scientifiques pour classer les êtres vivants ne sont pas très utiles. Les biologistes ont utilisé plusieurs systèmes de classification par le passé, basés sur la morphologie et la génétique des différents virus. Cependant, ces méthodes de classification antérieures regroupaient les virus différemment, en fonction des caractéristiques du virus utilisées pour les classer. La méthode de classification la plus couramment utilisée aujourd'hui s'appelle le schéma de classification de Baltimore et est basée sur la façon dont l'ARN messager (ARNm) est généré dans chaque type de virus particulier.
Anciens systèmes de classification
Les virus sont classés de différentes manières : en fonction de facteurs tels que leur contenu principal (tableau\(\PageIndex{1}\) et figure\(\PageIndex{2}\)), la structure de leurs capsides et leur enveloppe extérieure. Le type de matériel génétique (ADN ou ARN) et sa structure (simple ou double brin, linéaire ou circulaire, segmenté ou non segmenté) sont utilisés pour classer les structures centrales du virus.
| Classifications fondamentales | Exemples |
|---|---|
|
|
|
|
|
|
|
|
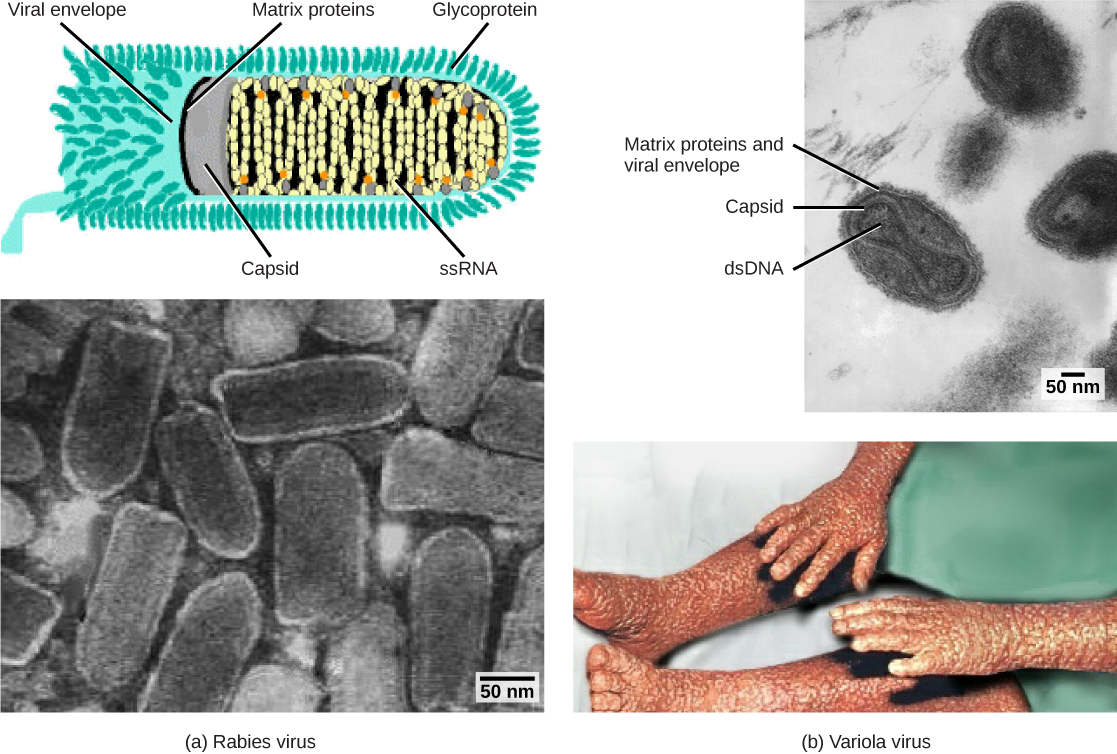
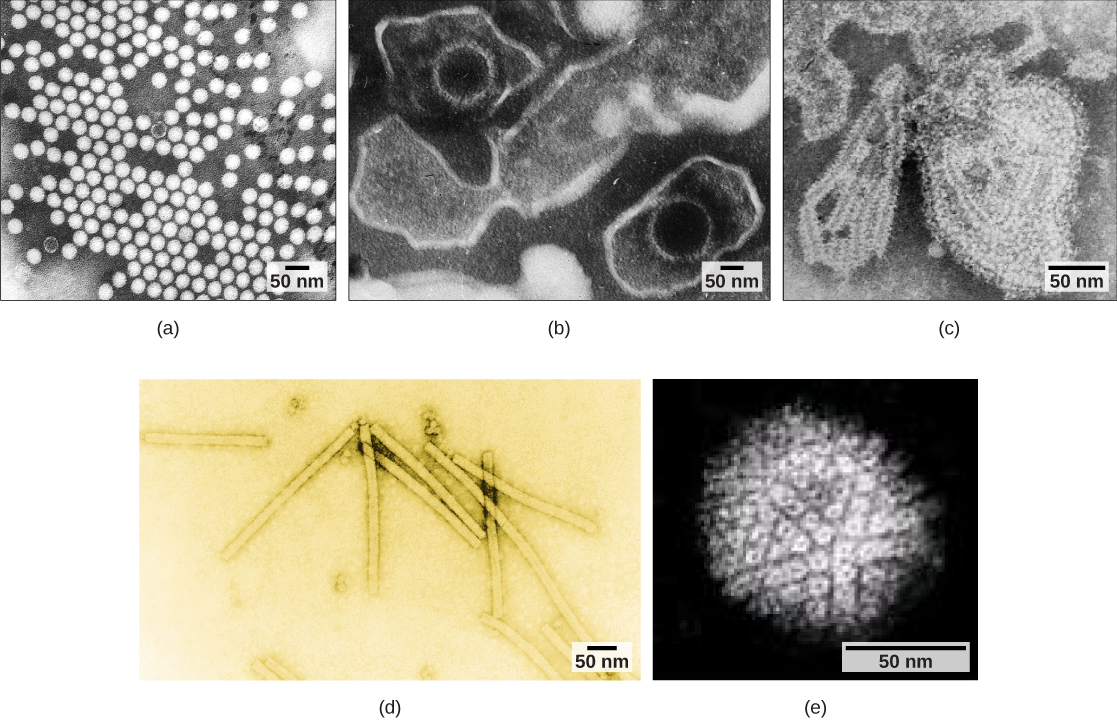
Les virus peuvent également être classés en fonction de la conception de leurs capsides (Figure\(\PageIndex{3}\) et Figure\(\PageIndex{4}\)). Les capsides sont classées comme étant icosaédriques nues, icosaédriques enveloppées, hélicoïdales enveloppées, hélicoïdales nues et complexes (Figure\(\PageIndex{5}\) et Figure\(\PageIndex{6}\)). Le type de matériel génétique (ADN ou ARN) et sa structure (simple ou double brin, linéaire ou circulaire, segmenté ou non segmenté) sont utilisés pour classer les structures centrales du virus (Tableau\(\PageIndex{2}\)).
| Classification des capsides | Exemples |
|---|---|
| Icosaédrique nue | Virus de l'hépatite A, poliovirus |
| Icosaèdre enveloppé | Virus Epstein-Barr, virus de l'herpès simplex, virus de la rubéole, virus de la fièvre jaune, VIH-1 |
| Hélicoïdal enveloppé | Virus de la grippe, virus des oreillons, virus de la rougeole, virus de la rage |
| Hélicoïdal nu | Virus de la mosaïque |
| Complexe contenant de nombreuses protéines ; certains présentent des combinaisons de structures de capsides icosaédriques et hélicoïdales | Herpèsvirus, virus de la variole, virus de l'hépatite B, bactériophage T4 |
Classement de Baltimore
Le système de classification des virus le plus couramment utilisé a été développé par le biologiste David Baltimore, lauréat du prix Nobel, au début des années 1970. Outre les différences morphologiques et génétiques mentionnées ci-dessus, le système de classification de Baltimore regroupe les virus en fonction de la façon dont l'ARNm est produit au cours du cycle réplicatif du virus.
Les virus du groupe I contiennent de l'ADN double brin (DSDNA) comme génome. Leur ARNm est produit par transcription de la même manière que pour l'ADN cellulaire. Le génome des virus du groupe II est constitué d'ADN monocaténaire (ADNss). Ils convertissent leurs génomes monocaténaires en un intermédiaire d'ADNdN avant que la transcription en ARNm puisse avoir lieu. Les virus du groupe III utilisent le dsRNA comme génome. Les brins se séparent et l'un d'eux est utilisé comme matrice pour la génération d'ARNm à l'aide de l'ARN polymérase dépendante de l'ARN codée par le virus. Les virus du groupe IV ont comme génome l'ARNss avec une polarité positive. Une polarité positive signifie que l'ARN génomique peut servir directement d'ARNm. Les intermédiaires du dsRNA, appelés intermédiaires réplicatifs, sont fabriqués lors du processus de copie de l'ARN génomique. De multiples brins d'ARN complets de polarité négative (complémentaires à l'ARN génomique à brin positif) sont formés à partir de ces intermédiaires, qui peuvent ensuite servir de modèles pour la production d'ARN de polarité positive, y compris à la fois de l'ARN génomique complet et des ARNm viraux plus courts. Les virus du groupe V contiennent des génomes d'ARNss de polarité négative, ce qui signifie que leur séquence est complémentaire de l'ARNm. Comme pour les virus du groupe IV, les intermédiaires du dsRNA sont utilisés pour fabriquer des copies du génome et produire de l'ARNm. Dans ce cas, le génome à brin négatif peut être converti directement en ARNm. De plus, des brins d'ARN positifs sur toute la longueur sont conçus pour servir de modèles pour la production du génome à brins négatifs. Les virus du groupe VI possèdent des génomes d'ARNss diploïdes (deux copies) qui doivent être convertis, à l'aide de l'enzyme transcriptase inverse, en dSDNA ; le dsDNA est ensuite transporté vers le noyau de la cellule hôte et inséré dans le génome de l'hôte. Ensuite, l'ARNm peut être produit par transcription de l'ADN viral qui a été intégré au génome de l'hôte. Les virus du groupe VII possèdent des génomes partiels du dsDNA et fabriquent des intermédiaires d'ARNss qui agissent comme des ARNm, mais sont également reconvertis en génomes d'ADNds par transcriptase inverse, nécessaire à la réplication du génome. Les caractéristiques de chaque groupe de la classification de Baltimore sont résumées dans le tableau\(\PageIndex{3}\) avec des exemples de chaque groupe.
| Groupe | Caractéristiques | Mode de production d'ARNm | Exemple |
|---|---|---|---|
| JE | ADN bicaténaire | L'ARNm est transcrit directement à partir de la matrice d'ADN | Herpès simplex (herpèsvirus) |
| II | ADN monocaténaire | L'ADN est converti en une forme double brin avant que l'ARN ne soit transcrit | Parvovirus canin (parvovirus) |
| III | ARN bicaténaire | L'ARNm est transcrit à partir du génome de l'ARN | Gastro-entérite infantile (rotavirus) |
| INTRAVEINEUSE | ARN monocaténaire (+) | Le génome fonctionne comme ARNm | Rhume (pircornavirus) |
| V | ARN monocaténaire (-) | L'ARNm est transcrit à partir du génome de l'ARN | Rage (rhabdovirus) |
| VI | Virus à ARN monocaténaire avec transcriptase inverse | La transcriptase inverse fabrique de l'ADN à partir du génome de l'ARN ; l'ADN est ensuite incorporé dans le génome de l'hôte ; l'ARNm est transcrit à partir de l'ADN incorporé | Virus de l'immunodéficience humaine (VIH) |
| VII | Virus à ADN double brin avec transcriptase inverse | Le génome viral est de l'ADN double brin, mais l'ADN viral est répliqué par un intermédiaire d'ARN ; l'ARN peut servir directement d'ARNm ou de matrice pour fabriquer de l'ARNm | Virus de l'hépatite B (hepadnavirus) |
Résumé
Les virus sont de minuscules entités acellulaires qui ne peuvent généralement être observées qu'au microscope électronique. Leurs génomes contiennent de l'ADN ou de l'ARN, jamais les deux, et ils se répliquent à l'aide des protéines de réplication d'une cellule hôte. Les virus sont divers et infectent les archées, les bactéries, les champignons, les plantes et les animaux. Les virus sont constitués d'un noyau d'acide nucléique entouré d'une capside protéique avec ou sans enveloppe lipidique externe. La forme de la capside, la présence d'une enveloppe et la composition du noyau dictent certains éléments de la classification des virus. La méthode de classification la plus couramment utilisée, la classification de Baltimore, classe les virus en fonction de la façon dont ils produisent leur ARNm.
Lexique
- acellulaire
- cellules manquantes
- capside
- enrobage protéique du noyau viral
- capsomère
- sous-unité protéique qui constitue la capside
- enveloppe
- bicouche lipidique qui enveloppe certains virus
- virus du groupe I
- virus doté d'un génome dSDNA
- virus du groupe II
- virus avec un génome SSDNA
- virus du groupe III
- virus avec un génome d'ARNdsR
- virus du groupe IV
- virus avec un génome d'ARNss à polarité positive
- virus du groupe V
- virus avec un génome d'ARNss à polarité négative
- virus du groupe VI
- virus dont les génomes d'ARNss sont convertis en dsDNA par transcriptase inverse
- virus du groupe VII
- virus avec un ARNm monocaténaire converti en ADNd pour la réplication du génome
- protéine matricielle
- protéine d'enveloppe qui stabilise l'enveloppe et joue souvent un rôle dans l'assemblage des virions de descendance
- polarité négative
- Virus à ARNs dont le génome est complémentaire à celui de leur AR
- polarité positive
- virus SSRNA dont le génome contient les mêmes séquences de base et les mêmes codons que ceux que l'on trouve dans son ARNm
- intermédiaire réplicatif
- Intermédiaire d'ARNds fabriqué lors du processus de copie de l'ARN génomique
- transcriptase inverse
- enzyme présente dans les groupes VI et VII de Baltimore qui convertit l'ARN simple brin en ADN double brin
- récepteur viral
- glycoprotéine utilisée pour fixer un virus aux cellules hôtes par l'intermédiaire de molécules présentes sur la cellule
- virion
- particule virale individuelle à l'extérieur d'une cellule hôte
- noyau viral
- contient le génome du virus