
20.3 : Perspectives sur l'arbre phylogénétique
- Page ID
- 189288

Compétences à développer
- Décrire le transfert horizontal de gènes
- Illustrez comment les procaryotes et les eucaryotes transfèrent des gènes horizontalement
- Identifier les modèles de réseaux et d'anneaux des relations phylogénétiques et décrire en quoi ils diffèrent du concept original d'arbre phylogénétique
Les concepts de modélisation phylogénétique évoluent constamment. C'est l'un des domaines d'études les plus dynamiques de toute la biologie. Au cours des dernières décennies, de nouvelles recherches ont remis en question les idées des scientifiques sur la façon dont les organismes sont liés. De nouveaux modèles de ces relations ont été proposés pour examen par la communauté scientifique.
De nombreux arbres phylogénétiques ont été présentés comme modèles de la relation évolutive entre les espèces. Les arbres phylogénétiques sont nés de Charles Darwin, qui a dessiné le premier arbre phylogénétique en 1837 (Figure\(\PageIndex{1}\) a), qui a servi de modèle pour des études ultérieures pendant plus d'un siècle. Le concept d'un arbre phylogénétique avec un tronc unique représentant un ancêtre commun et dont les branches représentent la divergence des espèces par rapport à cet ancêtre, correspond bien à la structure de nombreux arbres communs, tels que le chêne (Figure\(\PageIndex{1}\) b). Cependant, les preuves issues de l'analyse moderne des séquences d'ADN et des algorithmes informatiques récemment développés ont suscité le scepticisme quant à la validité du modèle d'arbre standard au sein de la communauté scientifique.

Limites du modèle classique
La conception classique de l'évolution procaryote, incluse dans le modèle d'arbre classique, est que les espèces évoluent de manière clonale. C'est-à-dire qu'ils produisent eux-mêmes une progéniture avec seulement des mutations aléatoires provoquant la descente dans la variété des espèces modernes et éteintes connues de la science. Ce point de vue est quelque peu complexe chez les eucaryotes qui se reproduisent sexuellement, mais les lois de la génétique mendélienne expliquent que la variation de la progéniture résulte, encore une fois, d'une mutation au sein de l'espèce. Le concept de transfert de gènes entre des espèces non apparentées n'a été envisagé comme une possibilité que relativement récemment. Le transfert horizontal de gènes (HGT), également connu sous le nom de transfert latéral de gènes, est le transfert de gènes entre des espèces non apparentées. Il a été démontré que le HGT est un phénomène omniprésent, de nombreux évolutionnistes postulant que ce processus joue un rôle majeur dans l'évolution, compliquant ainsi le modèle simple de l'arbre. Il a été démontré que des gènes sont transmis entre des espèces qui ne sont apparentées que de façon lointaine en utilisant la phylogénie standard, ce qui ajoute une couche de complexité à la compréhension des relations phylogénétiques.
Les différentes manières dont le HGT se produit chez les procaryotes sont importantes pour comprendre la phylogénie. Bien qu'à l'heure actuelle, le HGT ne soit pas considéré comme important pour l'évolution des eucaryotes, le HGT est également présent dans ce domaine. Enfin, à titre d'exemple de transfert génétique ultime, des théories de fusion du génome entre des organismes symbiotiques ou endosymbiotiques ont été proposées pour expliquer un événement d'une grande importance : l'évolution de la première cellule eucaryote, sans laquelle les humains n'auraient pas pu voir le jour.
Transfert horizontal de gènes
Le transfert horizontal de gènes (HGT) est l'introduction de matériel génétique d'une espèce à une autre par des mécanismes autres que la transmission verticale du ou des parents à la progéniture. Ces transferts permettent même à des espèces apparentées de partager des gènes, influençant ainsi leur phénotype. On pense que le HGT est plus répandu chez les procaryotes, mais qu'environ 2 % seulement du génome procaryote peut être transféré par ce processus. Certains chercheurs estiment que de telles estimations sont prématurées : l'importance réelle de l'HGT pour les processus évolutifs doit être considérée comme un travail en cours. Au fur et à mesure que le phénomène est étudié de manière plus approfondie, il peut s'avérer plus fréquent. De nombreux scientifiques pensent que le HGT et sa mutation semblent être (en particulier chez les procaryotes) une source importante de variation génétique, qui est la matière première du processus de sélection naturelle. Ces transferts peuvent se produire entre deux espèces qui partagent une relation intime (tableau\(\PageIndex{1}\)).
Tableau\(\PageIndex{1}\) : Résumé des mécanismes du HGT procaryote et eucaryote
| Mécanisme | Mode de transmission | Exemple | |
|---|---|---|---|
| Procaryotes | transformation | Capture d'ADN | de nombreux procaryotes |
| transduction | bactériophage (virus) | bactéries | |
| conjugaison | pilus | de nombreux procaryotes | |
| agents de transfert de gènes | particules semblables à des phages | bactéries violettes non sulfureuses | |
| Eucaryotes | provenant d'organismes alimentaires | inconnu | puceron |
| gènes sauteurs | transposons | plants de riz et de millet | |
| épiphytes/parasites | inconnu | champignons de l'if | |
| d'infections virales |
HGT chez les procaryotes
Il a été démontré que le mécanisme du HGT est assez courant dans les domaines procaryotes des bactéries et des archées, modifiant considérablement la façon dont leur évolution est perçue. La majorité des modèles évolutifs, tels que ceux de la théorie des endosymbiotes, proposent que les eucaryotes descendent de plusieurs procaryotes, ce qui rend le HGT d'autant plus important pour comprendre les relations phylogénétiques de toutes les espèces existantes et éteintes.
Les étudiants en microbiologie savent bien que des gènes sont transférés entre des bactéries communes. Ces transferts de gènes entre espèces constituent le principal mécanisme par lequel les bactéries acquièrent une résistance aux antibiotiques. Classiquement, on a pensé que ce type de transfert se produisait selon trois mécanismes différents :
- Transformation : l'ADN nu est absorbé par une bactérie
- Transduction : les gènes sont transférés à l'aide d'un virus
- Conjugaison : utilisation d'un tube creux appelé pilus pour transférer des gènes entre organismes
Plus récemment, un quatrième mécanisme de transfert de gènes entre les procaryotes a été découvert. De petites particules semblables à des virus appelées agents de transfert de gènes (GTA) transfèrent des segments génomiques aléatoires d'une espèce de procaryote à une autre. Il a été démontré que les GTA sont responsables des changements génétiques, parfois à une fréquence très élevée par rapport à d'autres processus évolutifs. Le premier GTA a été caractérisé en 1974 à l'aide de bactéries violettes non soufrées. Ces GTA, que l'on pense être des bactériophages qui ont perdu la capacité de se reproduire par eux-mêmes, transportent des fragments d'ADN aléatoires d'un organisme à l'autre. La capacité des GTA à agir à haute fréquence a été démontrée dans des études contrôlées utilisant des bactéries marines. Les transferts de gènes chez les procaryotes marins, que ce soit par des GTA ou par des virus, ont été estimés à 10 13 par an dans la seule mer Méditerranée. Les GTA et les virus sont considérés comme des véhicules HGT efficaces ayant un impact majeur sur l'évolution des procaryotes.
À la suite de cette analyse moderne de l'ADN, l'idée selon laquelle les eucaryotes auraient évolué directement à partir d'Archaea est tombée en désuétude. Bien que les eucaryotes partagent de nombreuses caractéristiques absentes des bactéries, telles que la boîte TATA (présente dans la région promotrice de nombreux gènes), la découverte selon laquelle certains gènes eucaryotes étaient plus homologues à l'ADN bactérien que l'ADN des archées a rendu cette idée moins défendable. De plus, la fusion des génomes des archées et des bactéries par endosymbiose a été proposée comme l'événement ultime de l'évolution eucaryote.
HGT chez les eucaryotes
Bien qu'il soit facile de voir comment les procaryotes échangent du matériel génétique par le HGT, on a d'abord pensé que ce processus était absent chez les eucaryotes. Après tout, les procaryotes ne sont que des cellules isolées directement exposées à leur environnement, alors que les cellules sexuelles des organismes multicellulaires sont généralement séquestrées dans des parties protégées du corps. Il découle de cette idée que les transferts de gènes entre eucaryotes multicellulaires devraient être plus difficiles. En effet, on pense que ce processus est plus rare chez les eucaryotes et qu'il a un impact évolutif beaucoup plus faible que chez les procaryotes. Malgré cela, la HGT entre des organismes apparentés à distance a été démontrée chez plusieurs espèces d'eucaryotes, et il est possible que d'autres exemples soient découverts à l'avenir.
Chez les plantes, un transfert de gènes a été observé chez des espèces qui ne peuvent pas se polliniser par des moyens normaux. Il a été démontré que les transposons ou « gènes sauteurs » se transfèrent entre les espèces de riz et de millet. De plus, les espèces fongiques qui se nourrissent des ifs, dont le médicament anticancéreux TAXOL® est dérivé de l'écorce, ont acquis la capacité de fabriquer elles-mêmes du taxol, un exemple clair de transfert de gènes.
Chez les animaux, un exemple particulièrement intéressant de HGT se trouve chez les espèces de pucerons (Figure\(\PageIndex{2}\)). Les pucerons sont des insectes dont la couleur varie en fonction de la teneur en caroténoïdes. Les caroténoïdes sont des pigments fabriqués par une variété de plantes, de champignons et de microbes, et ils remplissent diverses fonctions chez les animaux, qui obtiennent ces produits chimiques à partir de leur alimentation. Les humains ont besoin de caroténoïdes pour synthétiser la vitamine A, et nous les obtenons en mangeant des fruits et légumes oranges : carottes, abricots, mangues et patates douces. D'autre part, les pucerons ont acquis la capacité de fabriquer eux-mêmes les caroténoïdes. Selon l'analyse de l'ADN, cette capacité est due au transfert de gènes fongiques à l'insecte par le HGT, probablement parce que l'insecte consommait des champignons pour se nourrir. Une enzyme caroténoïde appelée désaturase est responsable de la coloration rouge observée chez certains pucerons, et il a également été démontré que lorsque ce gène est inactivé par mutation, les pucerons reprennent leur couleur verte plus courante (Figure\(\PageIndex{2}\)).

Fusion du génome et évolution des eucaryotes
Les scientifiques pensent que le summum de l'HGT se produit grâce à la fusion du génome entre différentes espèces de procaryotes lorsque deux organismes symbiotiques deviennent endosymbiotiques. Cela se produit lorsqu'une espèce est absorbée à l'intérieur du cytoplasme d'une autre espèce, ce qui aboutit finalement à un génome composé de gènes provenant à la fois de l'endosymbiote et de l'hôte. Ce mécanisme est un aspect de la théorie des endosymbiotes, qui est acceptée par la majorité des biologistes comme étant le mécanisme par lequel les cellules eucaryotes obtiennent leurs mitochondries et leurs chloroplastes. Cependant, le rôle de l'endosymbiose dans le développement du noyau est plus controversé. On pense que l'ADN nucléaire et l'ADN mitochondrial ont une origine évolutive différente (distincte), l'ADN mitochondrial étant dérivé des génomes circulaires de bactéries englouties par d'anciennes cellules procaryotes. L'ADN mitochondrial peut être considéré comme le plus petit chromosome. Il est intéressant de noter que l'ADN mitochondrial n'est hérité que de la mère. L'ADN mitochondrial se dégrade dans le sperme lorsque le sperme se dégrade dans l'ovule fécondé ou dans d'autres cas lorsque les mitochondries situées dans le flagelle du sperme ne pénètrent pas dans l'ovule.
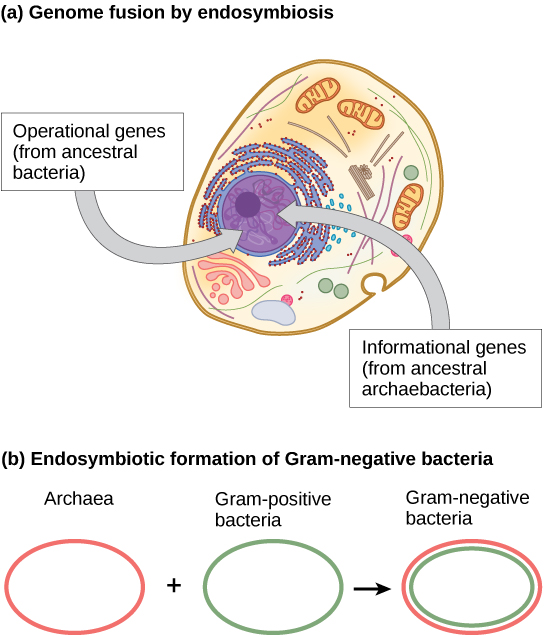
Au cours de la dernière décennie, James Lake, de l'Institut d'astrobiologie UCLA/NASA, a proposé que le processus de fusion du génome par endosymbiose soit responsable de l'évolution des premières cellules eucaryotes (Figure\(\PageIndex{3}\) a). À l'aide d'une analyse de l'ADN et d'un nouvel algorithme mathématique appelé reconstruction conditionnée (CR), son laboratoire a proposé que les cellules eucaryotes se développent à partir d'une fusion de gènes endosymbiotiques entre deux espèces, l'une archée et l'autre une bactérie. Comme nous l'avons mentionné, certains gènes eucaryotes ressemblent à ceux d'Archaea, tandis que d'autres ressemblent à ceux des bactéries. Un événement de fusion endosymbiotique, tel que celui proposé par Lake, expliquerait clairement cette observation. En revanche, ce travail est nouveau et l'algorithme CR est relativement peu étayé, ce qui amène de nombreux scientifiques à résister à cette hypothèse.
Des travaux plus récents de Lake (Figure\(\PageIndex{3}\) b) suggèrent que les bactéries gram-négatives, uniques dans leur domaine en ce sens qu'elles contiennent deux membranes lipidiques bicouches, sont en effet issues d'une fusion endosymbiotique d'espèces archéologiques et bactériennes. La double membrane serait le résultat direct de l'endosymbiose, l'endosymbiote prélevant la deuxième membrane de l'hôte au fur et à mesure de son internalisation. Ce mécanisme a également été utilisé pour expliquer les doubles membranes présentes dans les mitochondries et les chloroplastes. Les travaux de Lake ne sont pas exempts de scepticisme, et les idées font toujours l'objet de débats au sein de la communauté des sciences biologiques. Outre l'hypothèse de Lake, il existe plusieurs autres théories concurrentes quant à l'origine des eucaryotes. Comment a évolué le noyau eucaryote ? Selon une théorie, les cellules procaryotes auraient produit une membrane supplémentaire qui entourait le chromosome bactérien. L'ADN de certaines bactéries est entouré de deux membranes ; toutefois, rien n'indique la présence d'un nucléole ou de pores nucléaires. D'autres protéobactéries possèdent également des chromosomes liés à la membrane. Si le noyau eucaryote évoluait ainsi, on pourrait s'attendre à ce que l'un des deux types de procaryotes soit plus étroitement apparenté aux eucaryotes.
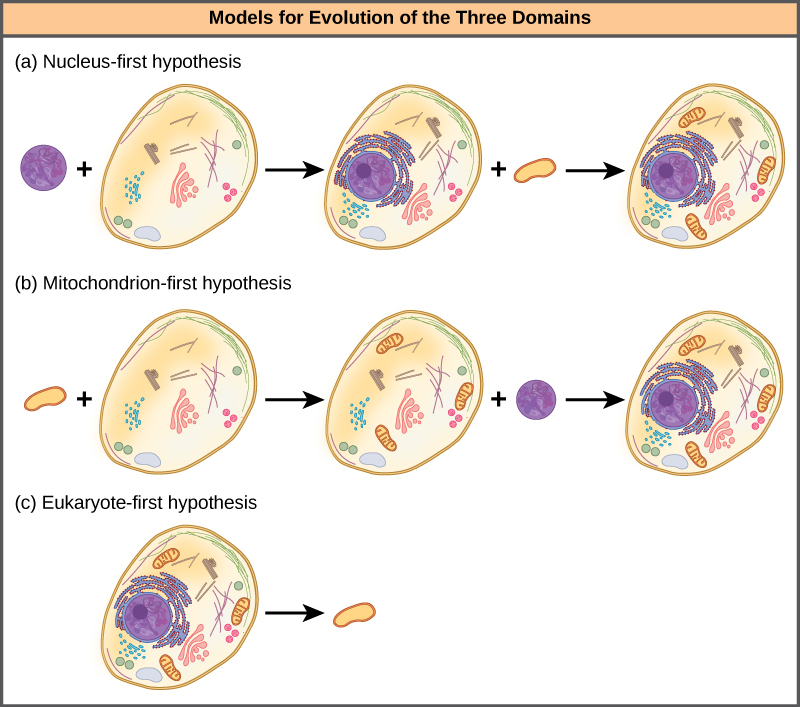
L'hypothèse du noyau d'abord suggère que le noyau a d'abord évolué chez les procaryotes (Figure\(\PageIndex{4}\) a), suivi d'une fusion ultérieure du nouvel eucaryote avec des bactéries devenues des mitochondries. L'hypothèse des mitochondries d'abord suggère que les mitochondries se sont d'abord établies chez un hôte procaryote (Figure\(\PageIndex{4}\) b), qui a ensuite acquis un noyau, par fusion ou par d'autres mécanismes, pour devenir la première cellule eucaryote. Plus intéressant encore, l'hypothèse des eucaryotes d'abord suggère que les procaryotes ont en fait évolué à partir des eucaryotes en perdant leurs gènes et leur complexité (Figure\(\PageIndex{4}\) c). Toutes ces hypothèses peuvent être testées. Seuls le temps et d'autres expériences permettront de déterminer quelle hypothèse est la mieux étayée par des données.
Modèles Web et de réseaux
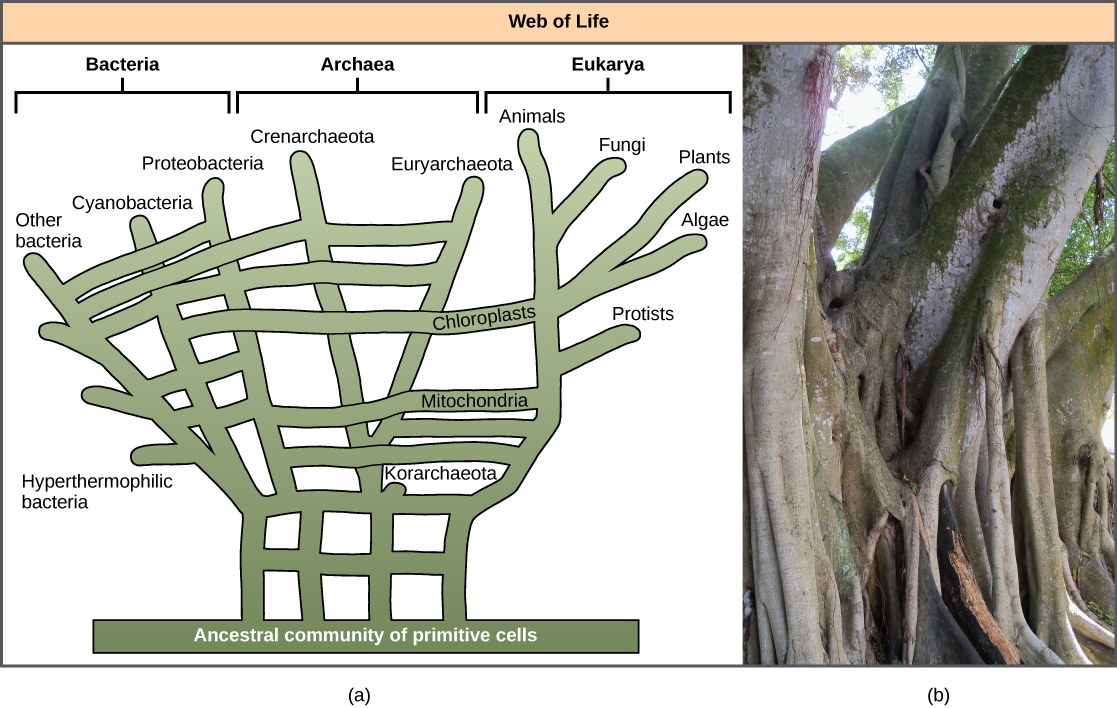
La reconnaissance de l'importance du HGT, notamment dans l'évolution des procaryotes, a amené certains à proposer l'abandon du modèle classique de l' « arbre de vie ». En 1999, W. Ford Doolittle a proposé un modèle phylogénétique qui ressemble davantage à une toile ou à un réseau qu'à un arbre. L'hypothèse est que les eucaryotes n'ont pas évolué à partir d'un seul ancêtre procaryote, mais à partir d'un pool de nombreuses espèces qui partageaient des gènes par des mécanismes HGT. Comme le montre la Figure\(\PageIndex{5}\) a, certains procaryotes individuels étaient responsables du transfert des bactéries responsables du développement mitochondrial vers les nouveaux eucaryotes, tandis que d'autres espèces ont transféré les bactéries à l'origine des chloroplastes. Ce modèle est souvent appelé la « toile de la vie ». Afin de sauvegarder l'analogie des arbres, certains ont proposé d'utiliser le ficus (Figure\(\PageIndex{5}\) b) avec ses multiples troncs à des fins phylogénétiques afin de représenter le rôle évolutif diminué du HGT.
Modèles Ring of Life
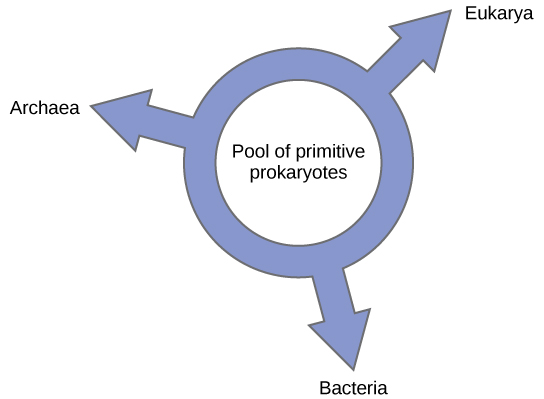
D'autres ont proposé d'abandonner tout modèle de phylogénie semblable à un arbre au profit d'une structure en anneau, appelée « anneau de vie » (Figure\(\PageIndex{6}\)) ; un modèle phylogénétique où les trois domaines de la vie ont évolué à partir d'un pool de procaryotes primitifs. Lake, utilisant à nouveau l'algorithme de reconstruction conditionnée, propose un modèle en anneau dans lequel les espèces des trois domaines (archées, bactéries et eucaryas) ont évolué à partir d'un seul pool de procaryotes échangeant des gènes. Son laboratoire suggère que cette structure est la mieux adaptée aux données issues d'analyses approfondies de l'ADN effectuées dans son laboratoire, et que le modèle en anneau est le seul qui tienne correctement compte de l'HGT et de la fusion génomique. Cependant, d'autres phylogénéticiens restent très sceptiques à l'égard de ce modèle.
En résumé, le modèle « arbre de vie » proposé par Darwin doit être modifié pour inclure le HGT. Cela signifie-t-il abandonner complètement le modèle d'arbre ? Même Lake soutient que tous les efforts devraient être faits pour découvrir une modification du modèle d'arbre afin de lui permettre d'ajuster exactement ses données, et que seule l'incapacité de le faire influencera les gens vers sa proposition d'anneau.
Cela ne signifie pas qu'un arbre, une toile ou un anneau seront complètement corrélés à une description précise des relations phylogénétiques de la vie. L'une des conséquences de la nouvelle réflexion sur les modèles phylogénétiques est l'idée que la conception originale de Darwin de l'arbre phylogénétique est trop simple, mais qu'elle avait du sens sur la base de ce que l'on savait à l'époque. Cependant, la recherche d'un modèle plus utile continue : chaque modèle sert d'hypothèses à tester avec la possibilité de développer de nouveaux modèles. C'est ainsi que la science progresse. Ces modèles sont utilisés sous forme de visualisations pour aider à construire des relations évolutives hypothétiques et à comprendre l'énorme quantité de données analysées.
Résumé
L'arbre phylogénétique, utilisé pour la première fois par Darwin, est le modèle classique d' « arbre de vie » décrivant les relations phylogénétiques entre les espèces, et le modèle le plus couramment utilisé aujourd'hui. De nouvelles idées sur l'HGT et la fusion du génome ont amené certains à suggérer de réviser le modèle pour qu'il ressemble à des toiles ou à des anneaux.
Lexique
- hypothèse eucaryote première
- proposition selon laquelle les procaryotes ont évolué à partir d'eucaryotes
- agent de transfert de gènes (GTA)
- particule semblable à un bactériophage qui transfère des segments génomiques aléatoires d'une espèce de procaryote à une autre
- fusion du génome
- fusion de deux génomes procaryotes, vraisemblablement par endosymbiose
- transfert horizontal de gènes (HGT)
- (également transfert latéral de gènes) transfert de gènes entre des espèces non apparentées
- hypothèse de la mitochondrie d'abord
- proposition selon laquelle les procaryotes auraient d'abord acquis une mitochondrie, suivie du développement nucléaire
- hypothèse selon laquelle le noyau est d'abord
- proposition selon laquelle les procaryotes ont d'abord acquis un noyau, puis la mitochondrie
- anneau de vie
- modèle phylogénétique où les trois domaines de la vie ont évolué à partir d'un pool de procaryotes primitifs
- toile de vie
- modèle phylogénétique qui tente d'intégrer les effets du transfert horizontal de gènes sur l'évolution