
14.6 : Réparation de l'ADN
- Page ID
- 189747

Compétences à développer
- Discutez des différents types de mutations de l'ADN
- Expliquer les mécanismes de réparation de
La réplication de l'ADN est un processus très précis, mais des erreurs peuvent parfois se produire, comme l'insertion d'une base incorrecte par une ADN polymérase. Les erreurs non corrigées peuvent parfois avoir de graves conséquences, comme le cancer. Les mécanismes de réparation corrigent les erreurs. Dans de rares cas, les erreurs ne sont pas corrigées, ce qui entraîne des mutations ; dans d'autres cas, les enzymes de réparation sont elles-mêmes mutées ou défectueuses.
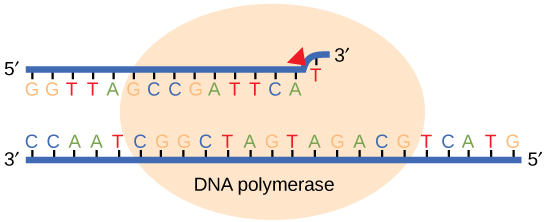
La plupart des erreurs lors de la réplication de l'ADN sont rapidement corrigées par l'ADN polymérase en relayant la base qui vient d'être ajoutée (Figure\(\PageIndex{1}\)). Lors de la relecture, le module ADN lit la base nouvellement ajoutée avant d'ajouter la suivante, afin de pouvoir effectuer une correction. La polymérase vérifie si la base nouvellement ajoutée s'est correctement associée à la base du brin modèle. S'il s'agit de la bonne base, le nucléotide suivant est ajouté. Si une base incorrecte a été ajoutée, l'enzyme coupe la liaison phosphodiester et libère le mauvais nucléotide. Ceci est réalisé par l'action exonucléasique de l'ADN pol III. Une fois que le nucléotide incorrect a été retiré, un nouveau nucléotide sera ajouté à nouveau.
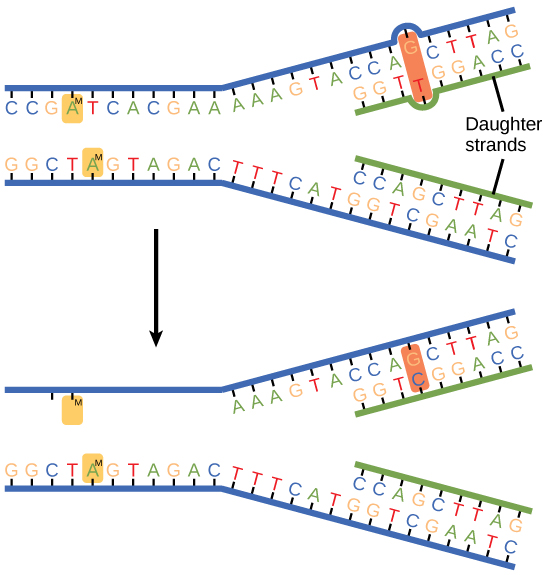
Certaines erreurs ne sont pas corrigées pendant la réplication, mais sont corrigées une fois la réplication terminée ; ce type de réparation est connu sous le nom de réparation des incohérences (Figure\(\PageIndex{2}\)). Les enzymes reconnaissent le nucléotide ajouté incorrectement et l'excisent ; celui-ci est ensuite remplacé par la base correcte. Si cela n'est pas corrigé, cela peut entraîner des dommages plus permanents. Comment les enzymes de réparation des mésappariements reconnaissent-elles laquelle des deux bases est la mauvaise ? Chez E. coli, après réplication, l'adénine de base azotée acquiert un groupe méthyle ; le brin d'ADN parental comportera des groupes méthyle, alors que le brin nouvellement synthétisé n'en possède pas. Ainsi, l'ADN polymérase est capable d'éliminer les bases mal incorporées du brin non méthylé nouvellement synthétisé. Chez les eucaryotes, le mécanisme n'est pas très bien compris, mais on pense qu'il implique la reconnaissance d'entailles non scellées dans le nouveau brin, ainsi qu'une association continue à court terme de certaines protéines de réplication avec le nouveau brin fille une fois la réplication terminée.
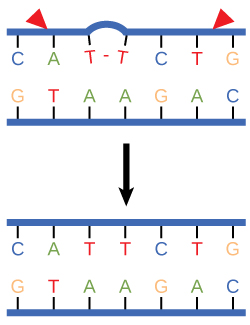
Dans un autre type de mécanisme de réparation, la réparation par excision nucléotidique, les enzymes remplacent les bases incorrectes en coupant à la fois les extrémités 3' et 5' de la base incorrecte (Figure\(\PageIndex{3}\)). Le segment d'ADN est retiré et remplacé par les nucléotides correctement appariés par l'action de l'ADN pol. Une fois les bases remplies, l'espace restant est scellé par une liaison phosphodiester catalysée par de l'ADN ligase. Ce mécanisme de réparation est souvent utilisé lorsque l'exposition aux UV provoque la formation de dimères de pyrimidine.
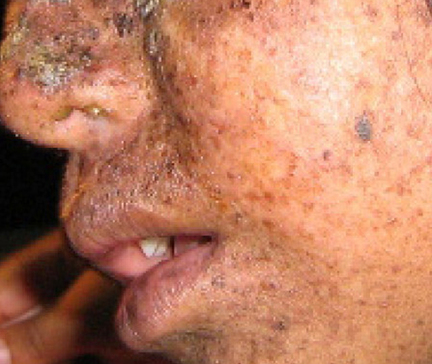
Un exemple bien étudié d'erreurs non corrigées est observé chez les personnes atteintes de xérodermie pigmentaire (Figure\(\PageIndex{4}\)). Les personnes atteintes ont une peau très sensible aux rayons UV du soleil. Lorsque les individus sont exposés aux UV, des dimères de pyrimidine, en particulier ceux de la thymine, se forment ; les personnes atteintes de xérodermie pigmentaire ne sont pas en mesure de réparer les dommages. Ils ne sont pas réparés en raison d'un défaut des enzymes de réparation de l'excision des nucléotides, alors que chez les individus normaux, les dimères de thymine sont excisés et le défaut est corrigé. Les dimères de thymine déforment la structure de la double hélice de l'ADN, ce qui peut entraîner des problèmes lors de la réplication de l'ADN. Les personnes atteintes de xérodermie pigmentaire peuvent avoir un risque plus élevé de contracter un cancer de la peau que celles qui n'en sont pas atteintes.
Les erreurs de réplication de l'ADN ne sont pas la seule raison pour laquelle des mutations apparaissent dans l'ADN. Des mutations, des variations de la séquence nucléotidique d'un génome, peuvent également survenir en raison de dommages causés à l'ADN. Ces mutations peuvent être de deux types : induites ou spontanées. Les mutations induites sont celles qui résultent d'une exposition à des produits chimiques, à des rayons UV, à des rayons X ou à tout autre agent environnemental. Les mutations spontanées se produisent sans exposition à aucun agent environnemental ; elles sont le résultat de réactions naturelles qui se produisent dans l'organisme.
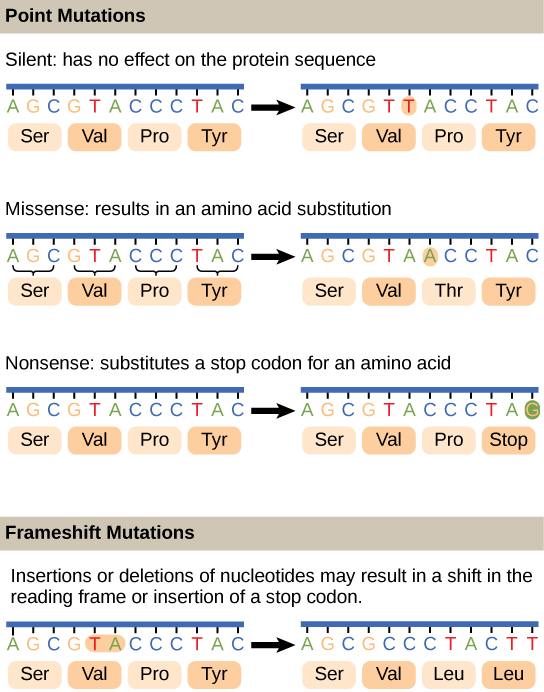
Les mutations peuvent avoir un large éventail d'effets. Certaines mutations ne sont pas exprimées ; elles sont appelées mutations silencieuses. Les mutations ponctuelles sont les mutations qui affectent une seule paire de bases. Les mutations nucléotidiques les plus courantes sont les substitutions, dans lesquelles une base est remplacée par une autre. Elles peuvent être de deux types, soit des transitions, soit des transversions. La substitution par transition fait référence au remplacement d'une purine ou d'une pyrimidine par une base du même type ; par exemple, une purine telle que l'adénine peut être remplacée par la guanine purique. La substitution par transversion fait référence au remplacement d'une purine par une pyrimidine, ou vice versa ; par exemple, la cytosine, une pyrimidine, est remplacée par l'adénine, une purine. Les mutations peuvent également résulter de l'ajout d'une base, appelée insertion, ou de la suppression d'une base, également appelée délétion. Parfois, un fragment d'ADN d'un chromosome peut être transféré vers un autre chromosome ou vers une autre région du même chromosome ; c'est ce que l'on appelle la translocation. Ces types de mutations sont illustrés sur la figure\(\PageIndex{5}\).
Exercice\(\PageIndex{1}\)
Une mutation par décalage de trame qui entraîne l'insertion de trois nucléotides est souvent moins délétère qu'une mutation qui entraîne l'insertion d'un nucléotide. Pourquoi ?
- Réponse
-
Si trois nucléotides sont ajoutés, un acide aminé supplémentaire sera incorporé dans la chaîne protéique, mais le cadre de lecture ne changera pas.
On sait que des mutations dans les gènes réparateurs peuvent provoquer le cancer. De nombreux gènes réparateurs mutés ont été impliqués dans certaines formes de cancer du pancréas, de cancer du côlon et de cancer colorectal. Les mutations peuvent affecter les cellules somatiques ou germinales. Si de nombreuses mutations s'accumulent dans une cellule somatique, elles peuvent entraîner des problèmes tels que la division cellulaire incontrôlée observée dans le cancer. Si une mutation se produit dans les cellules germinales, elle sera transmise à la génération suivante, comme dans le cas de l'hémophilie et de la xérodermie pigmentaire.
Résumé
L'ADN polymérase peut commettre des erreurs lors de l'ajout de nucléotides. Il édite l'ADN en relisant chaque base nouvellement ajoutée. Les bases incorrectes sont supprimées et remplacées par la base correcte, puis une nouvelle base est ajoutée. La plupart des erreurs sont corrigées lors de la réplication, mais lorsque cela ne se produit pas, le mécanisme de réparation des mésappariements est utilisé. Les enzymes de réparation des mésappariements reconnaissent la base mal incorporée et l'extraient de l'ADN pour la remplacer par la base correcte. Dans un autre type de réparation, la réparation par excision nucléotidique, la base incorrecte est retirée ainsi que quelques bases situées aux extrémités 5' et 3', et celles-ci sont remplacées par une copie de la matrice à l'aide de l'ADN polymérase. Les extrémités du fragment nouvellement synthétisé sont fixées au reste de l'ADN à l'aide de l'ADN ligase, qui crée une liaison phosphodiester.
La plupart des erreurs sont corrigées et, si ce n'est pas le cas, elles peuvent entraîner une mutation définie comme une modification permanente de la séquence d'ADN. Les mutations peuvent être de nombreux types, tels que la substitution, la délétion, l'insertion et la translocation. Les mutations des gènes réparateurs peuvent avoir de graves conséquences, telles que le cancer. Des mutations peuvent être induites ou se produire spontanément.
Lexique
- mutation induite
- mutation résultant d'une exposition à des produits chimiques ou à des agents environnementaux
- mutation
- variation de la séquence nucléotidique d'un génome
- réparation des discordances
- type de mécanisme de réparation dans lequel les bases non appariées sont supprimées après la réplication
- réparation par excision nucléotidique
- type de mécanisme de réparation de l'ADN dans lequel la mauvaise base, ainsi que quelques nucléotides en amont ou en aval, sont retirés
- relecture
- fonction de l'ADN pol dans laquelle il lit la base nouvellement ajoutée avant d'ajouter la suivante
- mutation ponctuelle
- mutation qui affecte une seule base
- mutation silencieuse
- mutation qui n'est pas exprimée
- mutation spontanée
- mutation qui se produit dans les cellules à la suite de réactions chimiques se produisant naturellement sans exposition à aucun agent externe
- substitution de transition
- lorsqu'une purine est remplacée par une purine ou qu'une pyrimidine est remplacée par une autre pyrimidine
- substitution par transversion
- lorsqu'une purine est remplacée par une pyrimidine ou lorsqu'une pyrimidine est remplacée par une purine